
Limosilactobacillus fermentum ING8, a Potential Multifunctional Non-Starter Strain with Relevant Technological Properties and Antimicrobial Activity




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of New LAB Strains
2.2. DNA Extraction and RAPD-PCR Analysis
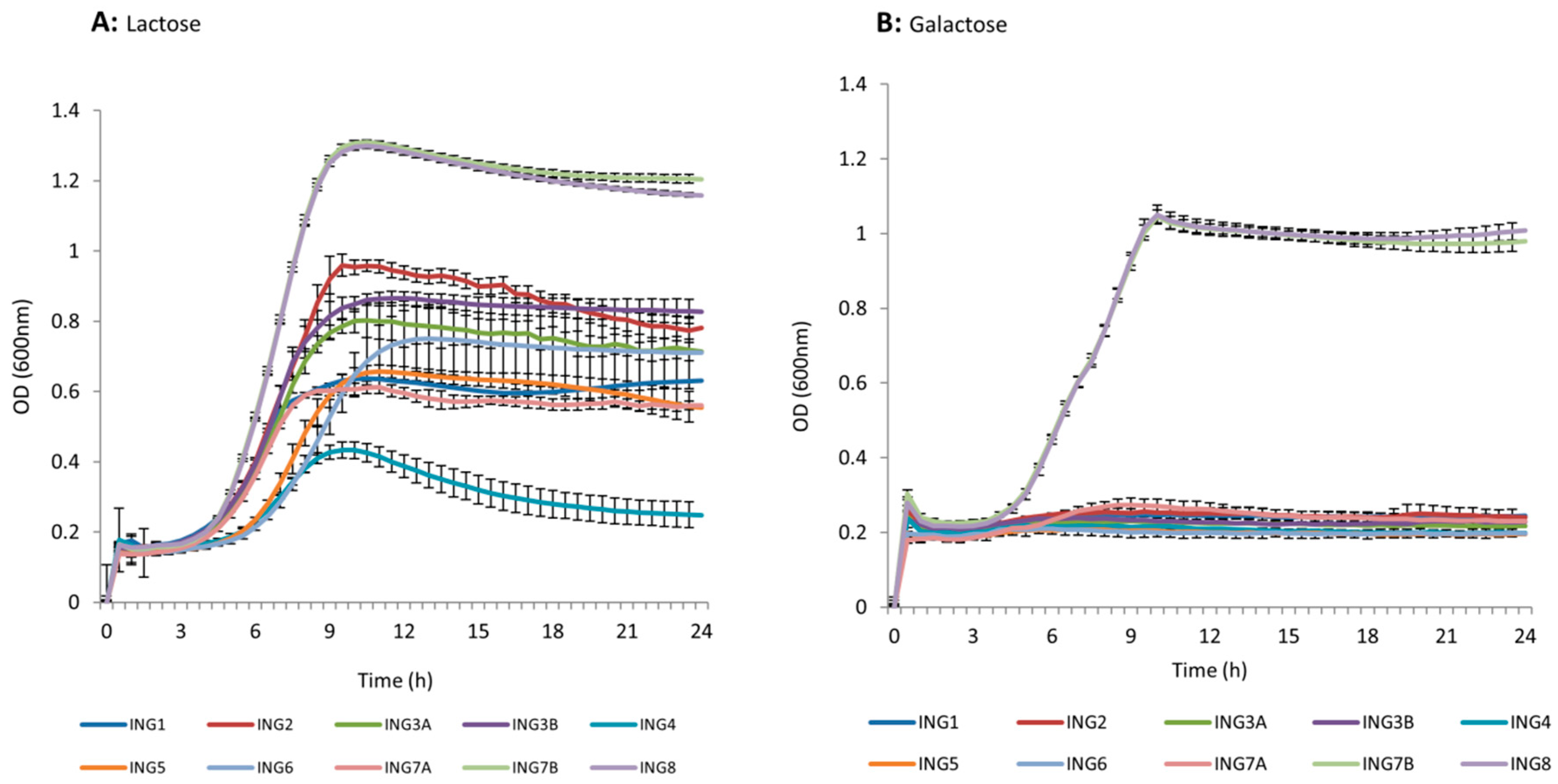
2.3. Lactose/Galactose Utilizers Selection
2.4. Exopolysaccharide Production and Quantification
2.5. Antimicrobial Activity Determination
2.6. Safety Assessments of Strain L. fermentum ING8
2.6.1. Minimum Inhibitory Concentration (MIC)
2.6.2. Hemolytic Activity Test
2.7. Technological Characterization of L. fermentum ING8
2.7.1. Enzymatic Activities
2.7.2. NaCl Tolerance Test
2.7.3. Growth at Different Temperatures
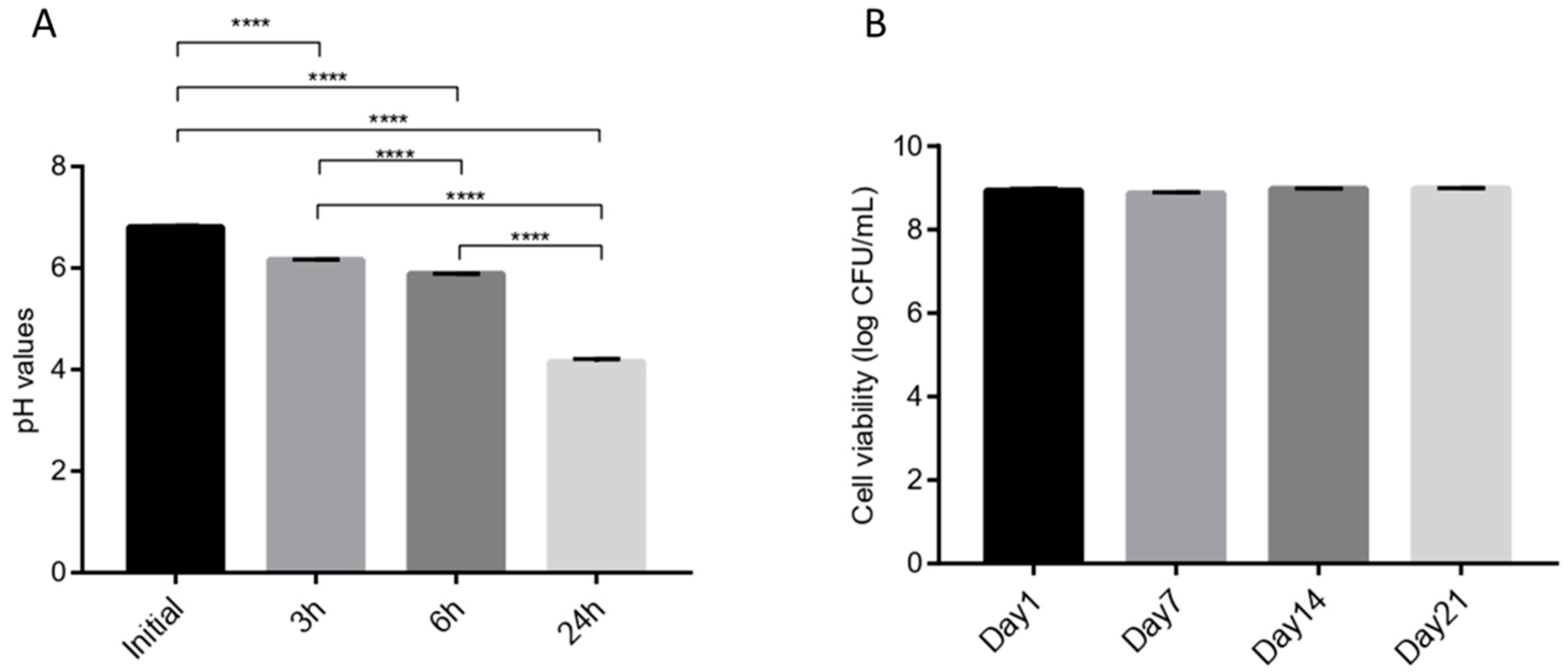
2.7.4. Acidification and Milk Coagulation
2.7.5. Viability during Storage
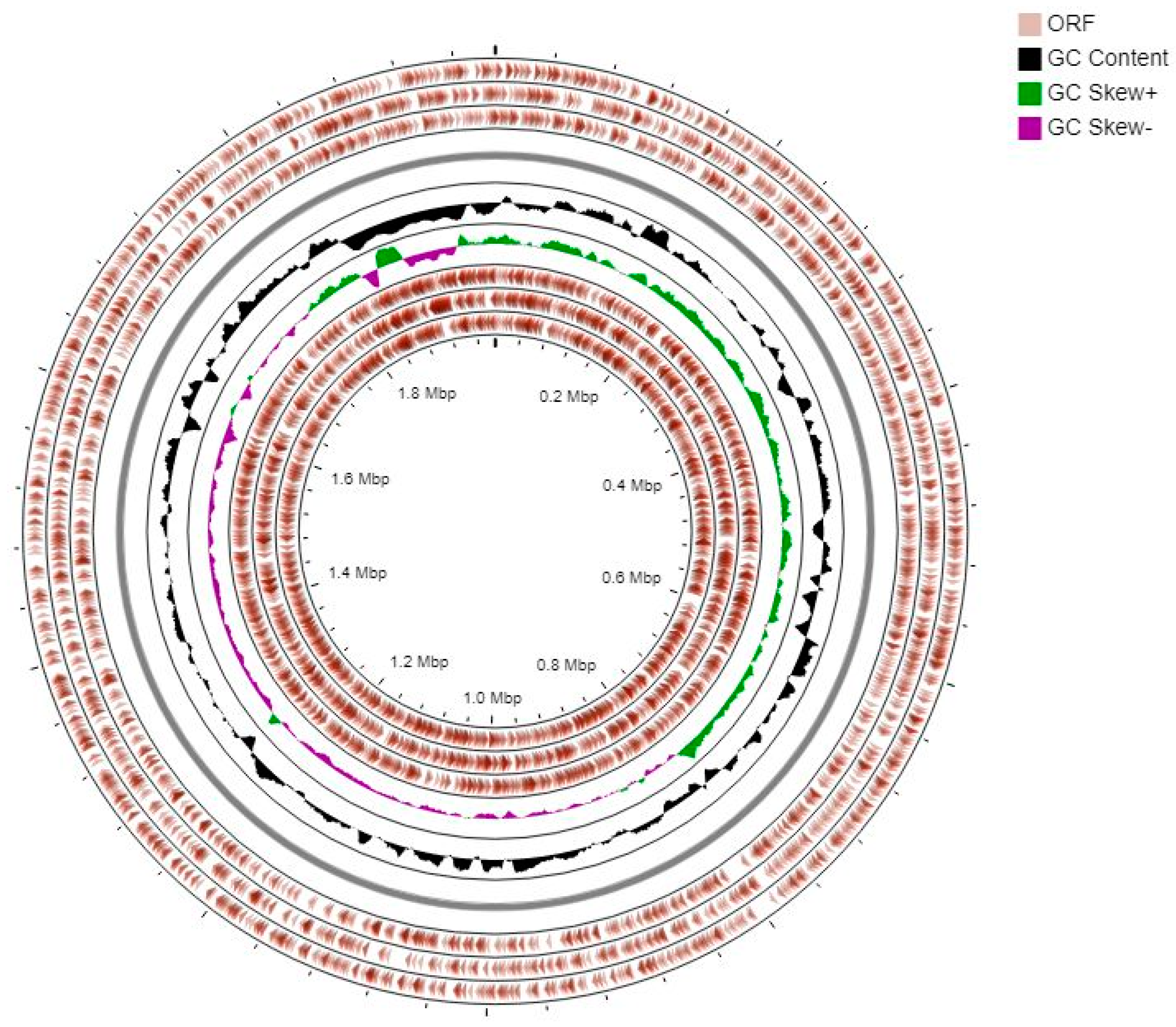
2.8. Genome Sequencing, Assembly, and Annotation of L. fermentum ING8
2.9. Statistical Analysis
3. Results
3.1. Strains Isolation and RAPD-PCR Analysis
3.2. Lactose and Galactose Utilization
3.3. Exopolysaccharide Production
3.4. Antimicrobial Activity Determination
3.5. Safety Assessments of L. fermentum ING8
3.6. Technological Properties of L. fermentum ING8
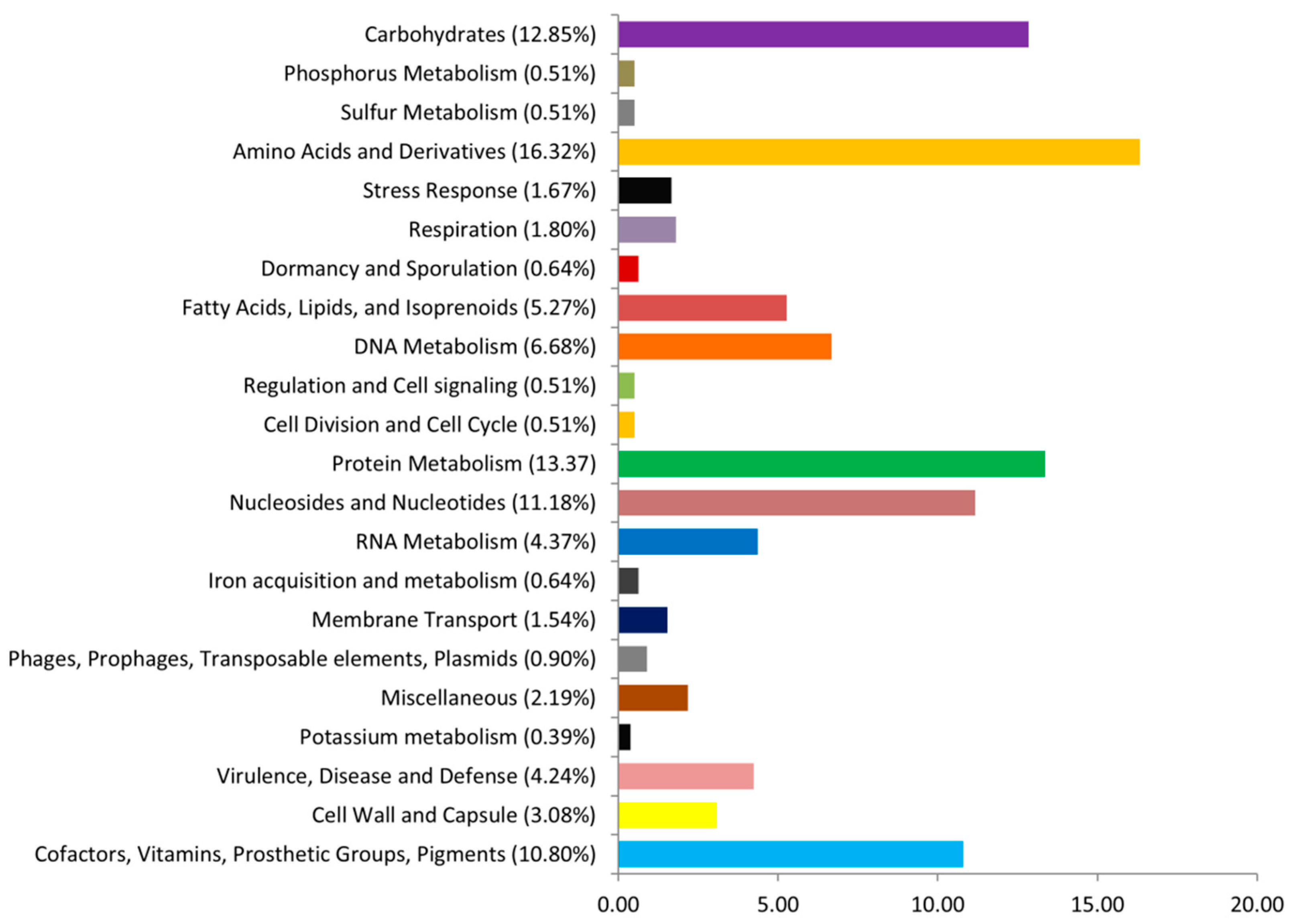
3.7. Genomic Analysis of L. fermentum ING8
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shangpliang, H.N.J.; Sharma, S.; Rai, R.; Tamang, J.P. Some technological properties of lactic acid bacteria isolated from Dahi and Datshi, naturally fermented milk products of Bhutan. Front. Microbiol. 2017, 8, 116. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Sur, M. Augmentation of biofunctionality of dahi. Int. J. Dairy Sci. 2017, 12, 243–253. [Google Scholar] [CrossRef]
- Akgun, A.; Yazici, F.; Gulec, H.A. Effect of reduced fat content on the physicochemical and microbiological properties of buffalo milk yoghurt. LWT 2016, 74, 521–527. [Google Scholar] [CrossRef]
- Ibrahim, S.A. Lactic Acid Bacteria: Lactobacillus spp. Other Species Ref Modul Food Sci; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Bao, Y.; Zhang, Y.; Zhang, Y.; Liu, Y.; Wang, S.; Dong, X.; Wang, Y.; Zhang, H. Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control 2010, 21, 695–701. [Google Scholar] [CrossRef]
- Plavec, T.V.; Berlec, A. Safety aspects of genetically modified lactic acid bacteria. Microorganisms 2020, 8, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ale, E.C.; Rojas, M.F.; Reinheimer, J.A.; Binetti, A.G. Lactobacillus fermentum: Could EPS production ability be responsible for functional properties? Food Microbiol. 2020, 90, 103465. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Alegría, Á.; Bron, P.A.; De Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C. Stress physiology of lactic acid bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igoshi, A.; Sato, Y.; Kameyama, K.; Murata, M. Galactose is the limiting factor for the browning or discoloration of cheese during storage. J. Nutr. Sci. Vitaminol. 2017, 63, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Crow, V.; Curry, B. Lactobacillus spp.|Other Species. In Encyclopedia of Dairy Sciences; Elsevier: Amsterdam, The Netherlands, 2002; pp. 1507–1511. [Google Scholar]
- Wu, Q.; Cheung, C.K.W.; Shah, N.P. Towards galactose accumulation in dairy foods fermented by conventional starter cultures: Challenges and strategies. Trends Food Sci. Technol. 2015, 41, 24–36. [Google Scholar] [CrossRef]
- Gaenzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Bibi, A.; Xiong, Y.; Rajoka, M.S.R.; Mehwish, H.M.; Radicetti, E.; Umair, M.; Shoukat, M.; Khan, M.K.I.; Aadil, R.M. Recent Advances in the Production of Exopolysaccharide (EPS) from Lactobacillus spp. and its application in the food industry: A Review. Sustainability 2021, 13, 12429. [Google Scholar] [CrossRef]
- Ryan, P.M.; Ross, R.P.; Fitzgerald, G.F.; Caplice, N.M.; Stanton, C. Sugar-coated: Exopolysaccharide producing lactic acid bacteria for food and human health applications. Food Funct. 2015, 6, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Reiner, K. Catalase test protocol. Am. Soc. Microbiol. 2010, 1–6. Available online: https://asm.org/Protocols/Catalase-Test-Protocol (accessed on 1 January 2022).
- Shields, P.; Cathcart, L. Oxidase test protocol. Am. Soc. Microbiol. 2010, 1–9. Available online: https://asm.org/Protocols/Oxidase-Test-Protocol (accessed on 1 January 2022).
- Tarrah, A.; da Silva Duarte, V.; de Castilhos, J.; Pakroo, S.; Junior, W.J.F.L.; Luchese, R.H.; Guerra, A.F.; Rossi, R.C.; Ziegler, D.R.; Corich, V. Probiotic potential and biofilm inhibitory activity of Lactobacillus casei group strains isolated from infant feces. J. Funct. Foods 2019, 54, 489–497. [Google Scholar] [CrossRef]
- Andrighetto, C.; Zampese, L.; Lombardi, A. RAPD-PCR characterization of lactobacilli isolated from artisanal meat plants and traditional fermented sausages of Veneto region (Italy). Lett. Appl. Microbiol. 2001, 33, 26–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, S.C.; Coelho, M.C.; Todorov, S.D.; Franco, B.; Dapkevicius, M.L.E.; Silva, C.C.G. Technological properties of bacteriocin-producing lactic acid bacteria isolated from P ico cheese an artisanal cow’s milk cheese. J. Appl. Microbiol. 2014, 116, 573–585. [Google Scholar] [CrossRef]
- Smitinont, T.; Tansakul, C.; Tanasupawat, S.; Keeratipibul, S.; Navarini, L.; Bosco, M.; Cescutti, P. Exopolysaccharide-producing lactic acid bacteria strains from traditional thai fermented foods: Isolation, identification and exopolysaccharide characterization. Int. J. Food Microbiol. 1999, 51, 105–111. [Google Scholar] [CrossRef]
- Faye, T.; Langsrud, T.; Nes, I.F.; Holo, H. Biochemical and genetic characterization of propionicin T1, a new bacteriocin from Propionibacterium thoenii. Appl. Environ. Microbiol. 2000, 66, 4230–4236. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163. [Google Scholar] [CrossRef]
- Tarrah, A.; da Silva Duarte, V.; Pakroo, S.; Corich, V.; Giacomini, A. Genomic and phenotypic assessments of safety and probiotic properties of Streptococcus macedonicus strains of dairy origin. Food Res. Int. 2020, 130, 108931. [Google Scholar] [CrossRef] [PubMed]
- Taheri, H.R.; Moravej, H.; Tabandeh, F.; Zaghari, M.; Shivazad, M. Screening of lactic acid bacteria toward their selection as a source of chicken probiotic. Poult. Sci. 2009, 88, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- Pakroo, S.; Ghion, G.; Tarrah, A.; Giacomini, A.; Corich, V. Effects of 2′-Fucosyllactose-Based Encapsulation on Probiotic Properties in Streptococcus thermophilus. Appl. Sci. 2021, 11, 5761. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Bosi, E.; Donati, B.; Galardini, M.; Brunetti, S.; Sagot, M.-F.; Lió, P.; Crescenzi, P.; Fani, R.; Fondi, M. MeDuSa: A multi-draft based scaffolder. Bioinformatics 2015, 31, 2443–2451. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Koller, M.; Bona, R.; Braunegg, G.; Hermann, C.; Horvat, P.; Kroutil, M.; Martinz, J.; Neto, J.; Pereira, L.; Varila, P. Production of polyhydroxyalkanoates from agricultural waste and surplus materials. Biomacromolecules 2005, 6, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.; Prazeres, A.R.; Rivas, J. Cheese whey wastewater: Characterization and treatment. Sci. Total Environ. 2013, 445, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Mollea, C.; Marmo, L.; Bosco, F. Valorisation of cheese whey, a by-product from the dairy industry. In Food Industry; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Curry, B.; Crow, V. Encyclopedia of Dairy Sciences; Elsevier Science Ltd.: New York, NY, USA, 2002. [Google Scholar]
- Behare, P.V.; Singh, R.; Tomar, S.K.; Nagpal, R.; Kumar, M.; Mohania, D. Effect of exopolysaccharide-producing strains of Streptococcus thermophilus on technological attributes of fat-free lassi. J. Dairy Sci. 2010, 93, 2874–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, K.; Shi, T.; Nagami, K.; Leo, F.; Nakamura, T.; Yasuda, K.; Senda, A.; Motoshima, H.; Urashima, T. Effects of carbohydrate source on physicochemical properties of the exopolysaccharide produced by Lactobacillus fermentum TDS030603 in a chemically defined medium. Carbohydr. Polym. 2010, 79, 1040–1045. [Google Scholar] [CrossRef]
- Wei, Y.; Li, F.; Li, L.; Huang, L.; Li, Q. Genetic and biochemical characterization of an exopolysaccharide with in vitro antitumoral activity produced by Lactobacillus fermentum YL-11. Front. Microbiol. 2019, 10, 2898. [Google Scholar] [CrossRef] [PubMed]
- Butorac, K.; Novak, J.; Bellich, B.; Terán, L.C.; Banić, M.; Leboš Pavunc, A.; Zjalić, S.; Cescutti, P.; Šušković, J.; Kos, B. Lyophilized alginate-based microspheres containing Lactobacillus fermentum D12, an exopolysaccharides producer, contribute to the strain’s functionality in vitro. Microb. Cell Fact. 2021, 20, 1–17. [Google Scholar] [CrossRef]
- Saxena, D.; Chakraborty, S.K.; Sabikhi, L.; Singh, D. Process optimization for a nutritious low-calorie high-fiber whey-based ready-to-serve watermelon beverage. J. Food Sci. Technol. 2015, 52, 960–967. [Google Scholar] [CrossRef] [Green Version]
- Povolo, S.; Toffano, P.; Basaglia, M.; Casella, S. Polyhydroxyalkanoates production by engineered Cupriavidus necator from waste material containing lactose. Bioresour. Technol. 2010, 101, 7902–7907. [Google Scholar] [CrossRef]
- Eustache, J.-M. Extraction of Glycoproteins and Sialic Acid from Whey. U.S. Patent 6,738,537, 16 August 1977. Available online: https://insight.rpxcorp.com/patent/US4042575A (accessed on 1 January 2022).
- Giaretta, S.; Treu, L.; Vendramin, V.; da Silva Duarte, V.; Tarrah, A.; Campanaro, S.; Corich, V.; Giacomini, A. Comparative transcriptomic analysis of Streptococcus thermophilus TH1436 and TH1477 showing different capability in the use of galactose. Front. Microbiol. 2018, 9, 1765. [Google Scholar] [CrossRef] [Green Version]
- Porto, M.C.W.; Kuniyoshi, T.M.; Azevedo, P.O.S.; Vitolo, M.; Oliveira, R.P. Pediococcus spp.: An important genus of lactic acid bacteria and pediocin producers. Biotechnol. Adv. 2017, 35, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Hickey, C.D.; Fallico, V.; Wilkinson, M.G.; Sheehan, J.J. Redefining the effect of salt on thermophilic starter cell viability, culturability and metabolic activity in cheese. Food Microbiol. 2018, 69, 219–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarrah, A.; Pakroo, S.; Junior, W.J.F.L.; Guerra, A.F.; Corich, V.; Giacomini, A. Complete Genome Sequence and Carbohydrates-Active EnZymes (CAZymes) Analysis of Lactobacillus paracasei DTA72, a Potential Probiotic Strain with Strong Capability to Use Inulin. Curr. Microbiol. 2020, 77, 2867–2874. [Google Scholar] [CrossRef] [PubMed]
- Tarrah, A.; Pakroo, S.; Corich, V.; Giacomini, A. Whole-genome sequence and comparative genome analysis of Lactobacillus paracasei DTA93, a promising probiotic lactic acid bacterium. Arch Microbiol. 2020, 202, 1997–2003. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ING 1 | ING 2 | ING 3A | ING 3B | ING 4 | ING 5 | ING 6 | ING 7A | ING 8 | |
|---|---|---|---|---|---|---|---|---|---|
| L. monocytogenes ATCC19117 | + | + | +++ | + | ++ | + | + | + | ++++ |
| B. cereus ATCC 11778 | ++ | + | + | + | + | + | + | + | ++++ |
| E. coli APEC18042/2 | + | + | + | + | ++ | +++ | + | + | ++++ |
| Antibiotics | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Strain/Antibiotics | chloramphenicol | Gentamicin | tetracycline | erythromycin | ampicillin | streptomycin | vancomycin | Ciprofloxacin | Kanamycin |
| L. fermentum ING8 | 4 (4) | 8 (16) | <0.125 (8) | 0.5 (1) | 0.5 (2) | 16 (64) | >64 (n.r.) ** | 4 (n.r.) | 32 (32) |
| Different Temperatures (°C) * | Different NaCl (%) ** | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 30 | 37 | 45 | 50 | 0 | 2 | 6 | 10 | Proteolytic activity *** | Lipolytic activity *** | Amylolytic activity *** | |
| L. fermentum ING8 | +++ | +++ | ++ | ++ | +++ | +++ | ++ | - | - | - | - |
| Feature | Value |
|---|---|
| Genome size | 1,981,384 |
| G + C content (%) | 51.3 |
| Contig N50 | 41442 |
| Contig L50 | 17 |
| Number of contigs | 158 |
| Number of Protein Coding Sequences (CDSs) | 2103 |
| Number of tRNAs | 57 |
| Number of rRNAs (5S, 16S, 23S) | 3 |
| Number of genes related to plasmid and acquired antibiotic-resistant gene | 0 |
| Number of genes related to Virulence, Disease, and Defense | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pakroo, S.; Tarrah, A.; Takur, R.; Wu, M.; Corich, V.; Giacomini, A. Limosilactobacillus fermentum ING8, a Potential Multifunctional Non-Starter Strain with Relevant Technological Properties and Antimicrobial Activity. Foods 2022, 11, 703. https://doi.org/10.3390/foods11050703
Pakroo S, Tarrah A, Takur R, Wu M, Corich V, Giacomini A. Limosilactobacillus fermentum ING8, a Potential Multifunctional Non-Starter Strain with Relevant Technological Properties and Antimicrobial Activity. Foods. 2022; 11(5):703. https://doi.org/10.3390/foods11050703
Chicago/Turabian StylePakroo, Shadi, Armin Tarrah, Rohit Takur, Manyu Wu, Viviana Corich, and Alessio Giacomini. 2022. "Limosilactobacillus fermentum ING8, a Potential Multifunctional Non-Starter Strain with Relevant Technological Properties and Antimicrobial Activity" Foods 11, no. 5: 703. https://doi.org/10.3390/foods11050703
APA StylePakroo, S., Tarrah, A., Takur, R., Wu, M., Corich, V., & Giacomini, A. (2022). Limosilactobacillus fermentum ING8, a Potential Multifunctional Non-Starter Strain with Relevant Technological Properties and Antimicrobial Activity. Foods, 11(5), 703. https://doi.org/10.3390/foods11050703