
Bioactive Components in Whole Grains for the Regulation of Skeletal Muscle Function




Abstract
:1. Introduction
2. Whole Grain Bioactive Compounds in the Regulation of Skeletal Muscle Function
2.1. Phenolic Compounds
2.1.1. Phenolic Acids
2.1.2. Resveratrol
2.1.3. Flavonoids
2.2. Carotenoids
2.3. Tocotrienol
2.4. β-Glucan
2.5. γ-Oryzanol
2.6. β-Sitosterol
2.7. Alkylresorcinols
2.8. Betaine
2.9. Octacosanol

{kind=link}
{kind=link}
| Components | Structure | Derived Grains | Content (mg/kg) | References |
|---|---|---|---|---|
| γ-oryzanol | 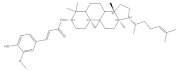 | Rice | 1550.0–8400.0 | [119] |
| Wheat | 297.0–584.0 | [120] | ||
| Corn | 200.0–250.0 | [120] | ||
| β-sitosterol | 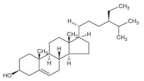 | Wheat | 365.6–673.1 | [124] |
| Barley | 510.0–676.0 | [51] | ||
| Quinoa | 76.0–556.0 | [39] | ||
| Oat | 210.0–409.2 | [39] | ||
| Alkylresorcinols | 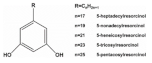 | |||
| Rye | 360.0–3200.0 | [108] | ||
| Wheat | 317.0–1732.0 | [108] | ||
| Ferulic acid | 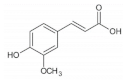 | Barley | 41.0–210.0 | [108] |
| Corn | 55.2 | [43] | ||
| Wheat | 472.0–813.8 | [40] | ||
| Barley | 41.0–210.0 | [108] | ||
| Quinoa | 126.8–281.7 | [42] | ||
| Brown rice | 7.1–52.7 | [50] | ||
| Rice | 155.6–271.1 | [41] | ||
| Oat | 1493.6 | [39] | ||
| p-coumaric acid | 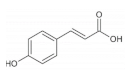 | Black rice | 84.8 ± 1.4 | [50] |
| Red rice | 176.9 ± 2.5 | [50] | ||
| Brown rice | 76.1–152.0 | [50] | ||
| Wheat | 23.8–35.6 | [40] | ||
| Barley | 0.8–58.4 | [51] | ||
| Quinoa | 31.3–42.0 | [42] | ||
| Resveratrol | 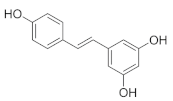 | Buckwheat | 5.7–7.9 | [54] |
| Quercetin | 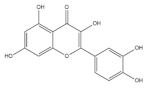 | Buckwheat | 3700.0 | [64] |
| Rice | 22.0–28.0 | [50] | ||
| Barley | 1.4–8.7 | [51] | ||
| Quinoa | 11.3–42.8 | [42] | ||
| Corn | 15.8 | [43] | ||
| Oat | 89.0 | [39] | ||
| Oligomeric procyanidins | 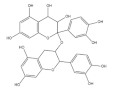 | Black rice | 3500.0 | [74] |
| Red rice | 200.0 | [74] | ||
| Cyanidin-3-glucoside | 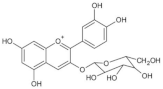 | Wheat | 1590.0 | [78] |
| Rice | 2682.0–4700.0 | [50] | ||
| Purple corn | 1430.0 | [43] | ||
| Rye | 2270.0 | [78] | ||
| Barley | 1020.0 | [78] | ||
| Oat | 430.0 | [78] | ||
| Catechins | 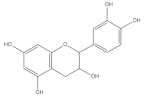 | Barley | 1312.47 ± 7.11 | [84] |
| Wheat | 355.87 | [83] | ||
| Buckwheat | 46.47 ± 0.17 | [84] | ||
| Rutin | 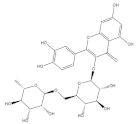 | Wheat | 236.2 | [93] |
| Quinoa | 609.1 | [94] | ||
| Buckwheat | 69.95 ± 2.25 | [84] | ||
| Carotenoids | 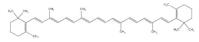 | Corn | 9.69–13.0 | [101] |
| Wheat | 32.1–39.7 | [101] | ||
| Barley | 0.15–10.5 | [101] | ||
| Betaine | 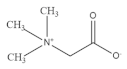 | Oat | 200.0–1000.0 | [132] |
| Wheat | 270.0–1110.0 | [132] | ||
| Rye | 444.0–2213.0 | [132] | ||
| Barley | 460.0–980.0 | [132] | ||
| Octacosanol |  | Rice | 95.7 | [138] |
| Wheat | 0.4–8.9 | [139] | ||
| β-glucan | 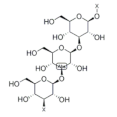 | Rice | 140.0–570.0 | [114] |
| Barley | 4000.0–7000.0 | [108] | ||
| Oat | 51,800.0–282,000.0 | [39] | ||
| Rye | 1200.0–2900.0 | [108] | ||
| Wheat | 400.0–1400.0 | [108] | ||
| Tocotrienol | 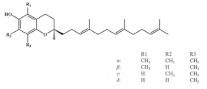 | Black rice | 31.9 ± 0.4 | [50] |
| Red rice | 36.9 ± 1.6 | [50] | ||
| Brown rice | 19.6 ± 0.4 | [50] | ||
| Wheat | 27.81 | [108] | ||
| Rye | 27.78 | [108] | ||
| Barley | 18.73 | [108] | ||
| Oat | 11.59 | [108] | ||
| Buckwheat | 54.6–552.2 | [39] |
3. Clinical Trials
| Compounds | Experimental Model | Effective Dose | Targets | Target Process | Effect | References |
|---|---|---|---|---|---|---|
| γ-oryzanol | Male C57BL/6 mice | 14.78 mg/kg/d | PPARδ ERRγ | Upregulate mitochondrial biogenesis and promote fatty acid β-oxidation | Reduce muscle weakness, alleviate inflammation and enhance muscle strength | [121] |
| Male Wistar rats | 0.5% (w/w) | GLUT4 | Promote antioxidant and anti-inflammatory effects | Alleviate insulin resistance in muscle | [122] | |
| β-sitosterol | ICR mice C2C12 cells | 35 or 350 μg/kg/d 15 μM | UCP3 | Upregulate the activity of complex proteins in the mitochondrial electron transport chain and induce mitochondrial uncoupling | Meet energy demand and promote muscle strength | [126] |
| Broilers | 100 mg/kg | PGC-1α/TFAM signaling | Upregulate oxidative status and mitochondrial biogenesis | Promote muscle performance | [127] | |
| L6 cells | 20 μM | LKB1/AMPK signaling | Increase ACC phosphorylation and triglyceride metabolism | Promote glucose homeostasis and lipid metabolism | [128] | |
| Male diabetic rats | 20 mg/kg/d | Rab/IRAP/Munc 18 pathway | Upregulate GLUT4 translocation and glycolytic and gluconeogenesis enzymes | Alleviate insulin resistance in muscle | [129] | |
| Male C57BL/6 mice | 200 mg/kg/d | FoxO1 signaling | Alleviate muscle protein loss and inhibit protein degradation | Alleviate muscle atrophy | [157] | |
| Alkylresorcinols | Male C57BL/6 mice | 0.4% (w/w) | Pyruvate dehydrogenase kinase 4 (PDK4) | Promote lipid-autophagy-induced lipid metabolism disorder | Inhibit the reduction in muscle size and prevent muscle atrophy | [4] |
| Male Wistar rats | 400 mg/kg/d | Glutathione peroxidase (GPx) | Inhibit oxidative damage caused by lipid accumulation and increase the expression of lactate dehydrogenase (LDH), alanine transaminase (ALT) | Alleviate myocardial infarction and exert an anti-fatigue effect | [130] | |
| Male C57BL/6J mice | 30 or 150 mg/kg/d | SIRT3/PGC-1α signaling | Increase mitochondrial content and mitochondrial biogenesis | Improve exercise capacity | [131] | |
| Ferulic acid | Male SD rats | 0.6 mg/kg/d | PKCε | Inhibit lipid-accumulation-induced inflammation | Alleviate insulin resistance | [44] |
| Isolated rat psoas muscle cells from Sprague Dawley male albino rats | 15–240 µg/mL | α-glucosidase and α-amylase | Promote fatty acid oxidative decomposition and inhibit carbohydrate and lipid hydrolyzing enzymes | Alleviate oxidative stress and mitigate redox imbalance | [5] | |
| C2C12 cells | 25 μM | PI3K/Cpkc signaling | Promote glucose uptake and glycogen synthesis | Improve insulin resistance | [45] | |
| Zebrafish | 0.06 mg/d | MyoD myogenin | Promote muscle growth | Increase the CSA of muscle fiber and muscle mass | [46] | |
| Duroc × Landrace × Yorkshire (DLY) weaned piglets | 0.05% or 0.45% (w/w) | SIRT1/AMPK/PGC-1α signaling | Improve the activity of SDH and MDH, increase the expression of slow MyHC protein | Increase the proportion of slow-twitch fiber and promote mitochondria function | [47] | |
| C2C12 cells | 0.5 or 1 μM | SIRT1/AMPK signaling | Increase the protein level of slow MyHC and decrease the protein level of fast MyHC | Promote slow oxidative muscle fiber formation and inhibit fast muscle fiber formation and exert an anti-fatigue function | [48] | |
| p-coumaric acid | L6 cells | 100 μM | AMPK signaling | Promote the fatty acid β-oxidation | Inhibit lipid-accumulation-induced inflammation in muscle | [53] |
| C2C12 cells | 0.1 mM | AMPK signaling | Increase expression of myogenin and myoD | Improve myogenic differentiation | [52] | |
| Resveratrol | Male C57 BL/6J mice | 50 mg/kg/d | AMPK/ FOXO3 signaling | Improve mitochondrial function | Improve muscle atrophy | [55] |
| Male C57BL/6J mice | 0.4% (w/w) | AMPK/PGC-1α signaling | Increase the level of muscle regeneration proteins including MyoG, Myf5 and Pax7 and mitochondrial biogenesis | Enhance muscle proliferation, differentiation and regeneration of impaired muscle | [57] | |
| L6 cells | 25 μM | PKA/LKB1/ AMPK pathway | Improve mitochondrial dysfunction and oxidative stress | Increase muscle mass and myofiber size and improve induced muscle atrophy | [56] | |
| Male C57BL/6J mice | 15 mg/kg/d | Increase muscle glycogen synthesis and reduce ROS levels | Reduce insulin resistance and promote lipid metabolism | [58] | ||
| C2C12 cells | 50 μM | AKT signaling | Modulate redox levels and glucose absorption | Reduce insulin resistance | [59] | |
| C2C12 cells | 100 μM | AKT/mTOR/FOXO1 signaling | Inhibit the atrophy-related ubiquitin ligase | Improve muscle hypertrophy and muscle atrophy | [60] | |
| Male Kunming mice | 400 mg/kg/d | AdiopR1–AMPK–PGC-1α signaling | Increase the expression of myosin heavy chain (MyHC) 1, MyHC2a and MyHC2x | Improve the transformation from fast- to slow-twitch muscle fibers and exercise performance | [61] | |
| C2C12 cells | 50 μM | miR-22-3p | Increase the activities of lactate dehydrogenase (SDH) and malate dehydrogenase (MDH) | Promote muscle-fiber-type conversion from fast-twitch to slow-twitch muscle fibers and exert an anti-fatigue effect | [62] | |
| Male ICR mice | 25 mg/kg/d | Increase the activities of LDH (lactic dehydrogenase) and creatine kinase (CK) | Improve muscle recovery and inflammation | [63] | ||
| Quercetin | Male Balb/c mice | 0.5% (w/w) | Adiponectin signaling | Decrease the fast MyHC and MyHC IIb protein expression | Promote muscle-fiber-type transformation from fast-twitch to slow-twitch muscle fibers | [66] |
| Male C57BL/6 mice | 0.05% (w/w) | HO-1/NRF2 signaling | Decrease inflammatory response and oxidative stress | Reduce obesity-induced muscle atrophy | [68] | |
| Male C57BL/6 mice | 0.2% (w/w) | PGC-1α signaling | Improve mitochondrial biogenesis and oxidative phosphorylation | Alleviate disuse-induced muscle atrophy | [69] | |
| C2C12 cells | 20 µM | AMPK signaling | Enhance insulin-stimulated glucose uptake and decrease inflammatory response | Ameliorate inflammation-induced insulin resistance | [70] | |
| Oligomeric procyanidins (OPCs) | Male ICR mice | 15 mg/kg/d | mTOR signaling | Increase glucose uptake and glycolysis, improve heat generation and inhibit gluconeogenesis and lipogenesis | Improve glucose homeostasis, lipid metabolism and insulin sensitivity | [75] |
| Human primary skeletal muscle cells | 10 or 25 μM | AKT signaling | Increase glycogen synthesis and glucose uptake | Improve glucose utilization and alleviate insulin resistance | [76] | |
| Male ICR mice | 10 μg/kg/d | AMPK signaling | Promote GLUT4 translocation | Increase insulin sensitivity | [77] | |
| Cyanidin-3-glucoside (Cy3G) | Male ICR mice | 1 mg/kg/d | PGC-1α signaling | Improve mitochondrial content and mitochondrial biogenesis | Improve exercise capacity | [82] |
| Human skeletal muscle cells | 100 µM | Inhibit the activity of α-amylase and α-glucosidase | Alleviate diabetes | [81] | ||
| Catechin | C2C12 cells | 20 µM | MyoD, MyoG, and MyHC | Promote myotube differentiation | Improve skeletal muscle regeneration and repair | [87] |
| Male C57BL/6 mice | 25 mg/kg/d | Wnt signaling | Promote myotube differentiation | Promote muscle regeneration | [90] | |
| C2C12 cells | 10 μM | Akt | Promote myotube differentiation | Improve muscle regeneration | [86] | |
| Male rats | 1.0 mg/kg/d | p38 MAPK signaling | Promote mitochondrial respiratory capacity and mitochondrial biogenesis | Enhance the ability of resisting fatigue | [88] | |
| Male Goto–Kakizaki (GK) rats | 100 mg/kg/d | ROS-ERK/JNK-p53 pathway | Promote mitochondrial autophagy | Alleviate diabetic-induced sarcopenia | [89] | |
| Male Sprague Dawley rats | 200 mg/kg/d | IGF-1 | Downregulate UPS-mediated muscle protein degradation and upregulate IGF-1-mediated muscle protein synthesis | Increase muscle mass | [91] | |
| Male SD rats | 0.1% (w/w) | Inhibit the expression of oxidative-modified proteins | Prevent muscle from oxidative stress induced by free radicals | [92] | ||
| Rutin | Male Sprague Dawley rats | 0.1% (w/w) | AMPK signaling | Enhance mitochondrial DNA (mtDNA) content and mitochondrial biogenesis | Improve obesity-induced muscle mitochondrial dysfunction | [97] |
| Male C57BL/6 mice | 60 mg/kg/d | PGC-1α signaling | Upregulate mediated mitochondrial biogenesis and decrease the level of lactic acid | Improve fatigue-resistance capacity | [98] | |
| C2C12 cells | 100 μM | NF-κB signaling | Decrease the expression of IL-6 and iNOS and the production of ROS | Inhibit oxidative-stress-induced skeletal muscle injury | [99] | |
| Lutein | Male Wistar rats | 0.5 mg/kg/d | NF-κB signaling | Reduce oxidative stress and inflammation and decrease the production of ROS | Improve skeletal muscle IR injury | [103] |
| β-carotene | Male Kwl: ddY mice | 0.5 mg/kg/d | IGF-1 | Promote protein synthesis and reduce ubiquitin-mediated muscle protein degradation | Increase muscle mass and prevent muscle hypertrophy | [105] |
| C2C12 cells | 10 μM | FOXO3A | Decrease the level of Atrogin-1 and MuRF1 | Increase muscle mass and exhibit an improvement effect on oxidative-stress-induced muscle atrophy | [107] | |
| Betaine | C2C12 cells | 10 mM | NFATc1/ MyoD signaling | Upregulate the expression of miR-29b-3p and promote myotube differentiation and the expression of slow MyHC proteins | Promote muscle cell differentiation and the transformation from fast muscle to slow muscle fiber | [136] |
| C2C12 cells | 10 mM | IGF-1 signaling | Increase the expression of MyoD and myogenin | Promote muscle fiber differentiation and growth | [134] | |
| C2C12 cells | 2 or 5 mM | PGC-1α signaling | Increase mitochondrial biogenesis and ATP production | Promote muscle differentiation and the transformation from fast muscle to slow muscle fiber | [137] | |
| Octacosanol | Male Wistar rats | 2.0 μCi/dose | Promote energy mobilization and energy supply | Enhance physical performance | [140] | |
| Male C57BL/6 mice | 200 mg/kg/d | Bcl3/TLRs/MAPK signaling | Increase the expression of Prx, Trim63 and ATPase activity | Exert an anti-fatigue effect | [143] | |
| Male SD rats | 0.75% (w/w) | Creatine phosphorylation | Promote the shift from fast-twitch to slow-twitch myofibers | Exert an anti-fatigue effect | [141] | |
| Male Wistar rats | 2.0 μCi/dose | ACC phosphorylation | Meet energy demand | Improve exercise capacity | [142] | |
| β-glucan | Male SD rats | 312.5 mg/kg/d | Decrease the activity of lactate dehydrogenase and the creatine kinase | Increase exercise capacity and facilitate the recovery from fatigue | [116] | |
| C2C12 cells | 20 mg/mL | Myf5 and Mox2 | Increase muscle cell proliferation and differentiation | Promote the transformation from fast muscle fibers to slow muscle fibers | [117] | |
| Duchenne muscular dystrophy (DMD) zebrafish model | 8 mg/L | Mitochondrial respiration enzyme | Improve mitochondrial respiration and prevent oxidative stress | Improve exercise capacity | [118] | |
| Tocotrienol | Male C57BL/6J mice | 100 or 300 mg/kg/d | AMPK/SIRT1/PGC-1α signaling | Upregulate the expression of proliferation and differentiation related proteins Increase mitochondrial biogenesis | Prevent diabetes-related skeletal muscle atrophy | [110] |
| Human skeletal muscle myoblasts | 50 μg/mL | miR-206 | Increase the expression of IGF1R and decrease the expression of Pax7 | Promote muscle cell proliferation and differentiation | [111] | |
| Stress-induced premature senescence (SIPS) Human skeletal muscle myoblasts (CHQ5B) | 50 μg/mL | Wnt signaling FOXO pathway | Downregulate the expression of MSTN and increase the expression of muscle cells regeneration related proteins such as EREG, SHC1 and SHC3 | Promote muscle cell regeneration and alleviate muscle loss | [112] | |
| Male C57BL/6J mice | 400 mg/kg/d | COXⅠ-Ⅴ | Promote mitochondrial respiration and reduce lipid peroxidation | Increase muscle mass and improve glucose homeostasis | [113] |
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Liu, J.; Yu, L.L.; Wu, Y. Bioactive Components and Health Beneficial Properties of Whole Wheat Foods. J. Agric. Food Chem. 2020, 68, 12904–12915. [Google Scholar] [CrossRef] [PubMed]
- Nogala-Katucka, M.; Kawka, A.; Dwiecki, K.; Siger, A. Evaluation of bioactive compounds in cereals study of wheat, barley, oat and selected grain products. Acta Sci. Pol.-Technol. Aliment. 2020, 19, 405–423. [Google Scholar] [CrossRef]
- Liu, J.; Hao, Y.; Wang, Z.; Ni, F.; Wang, Y.; Gong, L.; Sun, B.; Wang, J. Identification, Quantification, and Anti-inflammatory Activity of 5-n-Alkylresorcinols from 21 Different Wheat Varieties. J. Agric. Food Chem. 2018, 66, 9241–9247. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, S.; Yahata, N.; Saitoh, K.; Yoshimura, T.; Wang, Y.; Taniyama, S.; Nikawa, T.; Tachibana, K.; Hirasaka, K. Dietary supplementation with alkylresorcinols prevents muscle atrophy through a shift of energy supply. J. Nutr. Biochem. 2018, 61, 147–154. [Google Scholar] [CrossRef]
- Salau, V.F.; Erukainure, O.L.; Koorbanally, N.A.; Islam, M.S. Ferulic acid promotes muscle glucose uptake and modulate dysregulated redox balance and metabolic pathways in ferric-induced pancreatic oxidative injury. J. Food Biochem. 2022, 46, e13641. [Google Scholar] [CrossRef]
- Strasser, B.; Wolters, M.; Weyh, C.; Kruger, K.; Ticinesi, A. The Effects of Lifestyle and Diet on Gut Microbiota Composition, Inflammation and Muscle Performance in Our Aging Society. Nutrients 2021, 13, 2045. [Google Scholar] [CrossRef]
- Arenas-Jal, M.; Sune-Negre, J.M.; Perez-Lozano, P.; Garcia-Montoya, E. Trends in the food and sports nutrition industry: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 2405–2421. [Google Scholar] [CrossRef]
- Manore, M.; Meeusen, R.; Roelands, B.; Moran, S.; Popple, A.D.; Naylor, M.J.; Burke, L.M.; Stear, S.J.; Castell, L.M. BJSM reviews: A-Z of nutritional supplements: Dietary supplements, sports nutrition foods and ergogenic aids for health and performance-Part 16. Br. J. Sports Med. 2011, 45, 73–74. [Google Scholar] [CrossRef] [PubMed]
- Peeling, P.; Castell, L.M.; Derave, W.; de Hon, O.; Burke, L.M. Sports Foods and Dietary Supplements for Optimal Function and Performance Enhancement in Track-and-Field Athletes. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 198–209. [Google Scholar] [CrossRef]
- Tomlinson, D.J.; Erskine, R.M.; Morse, C.I.; Winwood, K.; Onambele-Pearson, G. The impact of obesity on skeletal muscle strength and structure through adolescence to old age. Biogerontology 2016, 17, 467–483. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Du, Y.; Trakooljul, N.; Brand, B.; Muryni, E.; Krischek, C.; Wicke, M.; Schwerin, M.; Wimmers, K.; Ponsuksili, S. Muscle Transcriptional Profile Based on Muscle Fiber, Mitochondrial Respiratory Activity, and Metabolic Enzymes. Int. J. Biol. Sci. 2015, 11, 1348–1362. [Google Scholar] [CrossRef] [PubMed]
- Blaauw, B.; Schiaffino, S.; Reggiani, C. Mechanisms Modulating Skeletal Muscle Phenotype. Compr. Physiol. 2013, 3, 1645–1687. [Google Scholar] [CrossRef]
- Khor, S.C.; Razak, A.M.; Ngah, W.Z.W.; Yusof, Y.A.M.; Karim, N.A.; Makpol, S. The Tocotrienol-Rich Fraction Is Superior to Tocopherol in Promoting Myogenic Differentiation in the Prevention of Replicative Senescence of Myoblasts. PLoS ONE 2016, 11, e0149265. [Google Scholar] [CrossRef]
- Lecker, S.H.; Goldberg, A.L.; Mitch, W.E. Protein degradation by the ubiquitin-proteasome pathway in normal and disease states. J. Am. Soc. Nephrol. 2006, 17, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.-N.; Ren, Y.; Chen, J.-Q.; Wang, Y.-Z. Effects of myogenin on muscle fiber types and key metabolic enzymes in gene transfer mice and C2C12 myoblasts. Gene 2013, 532, 246–252. [Google Scholar] [CrossRef]
- Qaisar, R.; Renaud, G.; Hedstrom, Y.; Pollanen, E.; Ronkainen, P.; Kaprio, J.; Alen, M.; Sipila, S.; Artemenko, K.; Bergquist, J.; et al. Hormone replacement therapy improves contractile function and myonuclear organization of single muscle fibres from postmenopausal monozygotic female twin pairs. J. Physiol.-Lond. 2013, 591, 2333–2344. [Google Scholar] [CrossRef] [PubMed]
- Qaisar, R.; Bhaskaran, S.; Van Remmenn, H. Muscle fiber type diversification during exercise and regeneration. Free. Radic. Biol. Med. 2016, 98, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Hoppeler, H.; Fluck, M. Plasticity of skeletal muscle mitochondria: Structure and function. Med. Sci. Sports Exerc. 2003, 35, 95–104. [Google Scholar] [CrossRef]
- Gan, Z.; Fu, T.; Kelly, D.P.; Vega, R.B. Skeletal muscle mitochondrial remodeling in exercise and diseases. Cell Res. 2018, 28, 969–980. [Google Scholar] [CrossRef]
- Heydemann, A. Skeletal Muscle Metabolism in Duchenne and Becker Muscular Dystrophy-Implications for Therapies. Nutrients 2018, 10, 796. [Google Scholar] [CrossRef] [Green Version]
- Narkar, V.A.; Downes, M.; Yu, R.T.; Embler, E.; Wang, Y.-X.; Banayo, E.; Mihaylova, M.M.; Nelson, M.C.; Zou, Y.; Juguilon, H.; et al. AMPK and PPAR delta Agonists are exercise mimetics. Cell 2008, 134, 405–415. [Google Scholar] [CrossRef]
- Liu, J.; Liang, X.; Zhou, D.; Lai, L.; Xiao, L.; Liu, L.; Fu, T.; Kong, Y.; Zhou, Q.; Vega, R.B.; et al. Coupling of mitochondrial function and skeletal muscle fiber type by a miR-499/Fnip1/AMPK circuit. Embo Mol. Med. 2016, 8, 1212–1228. [Google Scholar] [CrossRef]
- Reyes, N.L.; Banks, G.B.; Tsang, M.; Margineantu, D.; Gu, H.; Djukovic, D.; Chan, J.; Torres, M.; Liggitt, H.D.; Hirenallur-S, D.K.; et al. Fnip1 regulates skeletal muscle fiber type specification, fatigue resistance, and susceptibility to muscular dystrophy. Proc. Natl. Acad. Sci. USA 2015, 112, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Ljubicic, V.; Jasmin, B.J. AMP-activated protein kinase at the nexus of therapeutic skeletal muscle plasticity in Duchenne muscular dystrophy. Trends Mol. Med. 2013, 19, 614–624. [Google Scholar] [CrossRef]
- Spaulding, H.R.; Kelly, E.M.; Quindry, J.C.; Sheffield, J.B.; Hudson, M.B.; Selsby, J.T. Autophagic dysfunction and autophagosome escape in the mdx mus musculus model of Duchenne muscular dystrophy. Acta Physiol. 2018, 222, e12944. [Google Scholar] [CrossRef]
- Theilen, N.T.; Kunkel, G.H.; Tyagi, S.C. The Role of Exercise and TFAM in Preventing Skeletal Muscle Atrophy. J. Cell. Physiol. 2017, 232, 2348–2358. [Google Scholar] [CrossRef]
- Komatsu, R.; Okazaki, T.; Ebihara, S.; Kobayashi, M.; Tsukita, Y.; Nihei, M.; Sugiura, H.; Niu, K.; Ebihara, T.; Ichinose, M. Aspiration pneumonia induces muscle atrophy in the respiratory, skeletal, and swallowing systems. J. Cachexia Sarcopenia Muscle 2018, 9, 643–653. [Google Scholar] [CrossRef]
- Han, Z.; Chang, C.; Zhu, W.; Zhang, Y.; Zheng, J.; Kang, X.; Jin, G.; Gong, Z. Role of SIRT2 in regulating the dexamethasone-activated autophagy pathway in skeletal muscle atrophy. Biochem. Cell Biol. 2021, 99, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Theilen, N.T.; Jeremic, N.; Weber, G.J.; Tyagi, S.C. TFAM overexpression diminishes skeletal muscle atrophy after hindlimb suspension in mice. Arch. Biochem. Biophys. 2019, 666, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Siu, P.M. Muscle Apoptotic Response to Denervation, Disuse, and Aging. Med. Sci. Sports Exerc. 2009, 41, 1876–1886. [Google Scholar] [CrossRef]
- Falduto, M.T.; Czerwinski, S.M.; Hickson, R.C. Glucocorticoid-induced muscle atrophy prevention by exercise in fast-twitch fibers. J. Appl. Physiol. 1990, 69, 1058–1062. [Google Scholar] [CrossRef] [PubMed]
- Abrigo, J.; Campos, F.; Simon, F.; Riedel, C.; Cabrera, D.; Vilos, C.; Cabello-Verrugio, C. TGF-beta requires the activation of canonical and non-canonical signalling pathways to induce skeletal muscle atrophy. Biol. Chem. 2018, 399, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Lin, I.H.; Chang, J.-L.; Hua, K.; Huang, W.-C.; Hsu, M.-T.; Chen, Y.-F. Skeletal muscle in aged mice reveals extensive transformation of muscle gene expression. BMC Genet. 2018, 19, 55. [Google Scholar] [CrossRef]
- de Sire, R.; Rizzatti, G.; Ingravalle, F.; Pizzoferrato, M.; Petito, V.; Lopetuso, L.; Graziani, C.; de Sire, A.; Mentella, M.C.; Mele, M.C.; et al. Skeletal muscle-gut axis: Emerging mechanisms of sarcopenia for intestinal and extra intestinal diseases. Minerva Gastroenterol. Dietol. 2018, 64, 351–362. [Google Scholar] [CrossRef]
- Rasbach, K.A.; Gupta, R.K.; Ruas, J.L.; Wu, J.; Naseri, E.; Estall, J.L.; Spiegelman, B.M. PGC-1 alpha regulates a HIF2 alpha-dependent switch in skeletal muscle fiber types. Proc. Natl. Acad. Sci. USA 2010, 107, 21866–21871. [Google Scholar] [CrossRef] [PubMed]
- Pardo, P.S.; Boriek, A.M. The physiological roles of Sirt1 in skeletal muscle. Aging-Us 2011, 3, 430–437. [Google Scholar] [CrossRef]
- Sakuma, K.; Aoi, W.; Yamaguchi, A. Molecular mechanism of sarcopenia and cachexia: Recent research advances. Pflug. Arch.-Eur. J. Physiol. 2017, 469, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, Z.; Liu, J.; Hao, Y.; Sun, B.; Wang, J. Potential Health Benefits of Whole Grains: Modulation of Mitochondrial Biogenesis and Energy Metabolism. J. Agric. Food Chem. 2021, 69, 14065–14074. [Google Scholar] [CrossRef]
- Raguindin, P.F.; Itodo, O.A.; Stoyanov, J.; Dejanovic, G.M.; Gamba, M.; Asllanaj, E.; Minder, B.; Bussler, W.; Metzger, B.; Muka, T.; et al. A systematic review of phytochemicals in oat and buckwheat. Food Chemistry 2021, 338, 127982. [Google Scholar] [CrossRef] [PubMed]
- Saini, P.; Kumar, N.; Kumar, S.; Mwaurah, P.W.; Panghal, A.; Attkan, A.K.; Singh, V.K.; Garg, M.K.; Singh, V. Bioactive compounds, nutritional benefits and food applications of colored wheat: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2021, 61, 3197–3210. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li, G.; Li, M.; Xu, F.; Beta, T.; Bao, J. Genotypic variation in phenolic acids, vitamin E and fatty acids in whole grain rice. Food Chemistry 2016, 197, 776–782. [Google Scholar] [CrossRef]
- Han, Y.; Chi, J.; Zhang, M.; Zhang, R.; Fan, S.; Huang, F.; Xue, K.; Liu, L. Characterization of saponins and phenolic compounds: Antioxidant activity and inhibitory effects on alpha-glucosidase in different varieties of colored quinoa (Chenopodium quinoa Willd). Biosci. Biotechnol. Biochem. 2019, 83, 2128–2139. [Google Scholar] [CrossRef]
- Cristianini, M.; Sanchez, J.S. Extraction of bioactive compounds from purple corn using emerging technologies: A review. J. Food Sci. 2020, 85, 862–869. [Google Scholar] [CrossRef]
- Gogoi, B.; Chatterjee, P.; Mukherjee, S.; Buragohain, A.K.; Bhattacharya, S.; Dasgupta, S. A polyphenol rescues lipid induced insulin resistance in skeletal muscle cells and adipocytes. Biochem. Biophys. Res. Commun. 2014, 452, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.-B.; Chiang, B.-H. Amelioration of insulin resistance using the additive effect of ferulic acid and resveratrol on vesicle trafficking for skeletal muscle glucose metabolism. Phytother. Res. 2020, 34, 808–816. [Google Scholar] [CrossRef]
- Wen, Y.; Ushio, H. Ferulic Acid Promotes Hypertrophic Growth of Fast Skeletal Muscle in Zebrafish Model. Nutrients 2017, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Anis, E.; Zafeer, M.F.; Firdaus, F.; Islam, S.N.; Khan, A.A.; Ali, A.; Hossain, M.M. Ferulic acid reinstates mitochondrial dynamics through PGC1 alpha expression modulation in 6-hydroxydopamine lesioned rats. Phytother. Res. 2020, 34, 214–226. [Google Scholar] [CrossRef]
- Chen, X.; Guo, Y.; Jia, G.; Zhao, H.; Liu, G.; Huang, Z. Ferulic acid regulates muscle fiber type formation through the Sirt1/AMPK signaling pathway. Food Funct. 2019, 10, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Huang, Z.; Chen, D.; Yu, B.; Chen, H.; Yu, J.; Luo, Y.; Zheng, P.; He, J. Effects of dietary ferulic acid supplementation on growth performance and skeletal muscle fiber type conversion in weaned piglets. J. Sci. Food Agric. 2021, 101, 5116–5123. [Google Scholar] [CrossRef] [PubMed]
- Gong, E.S.; Liu, C.; Li, B.; Zhou, W.; Chen, H.; Li, T.; Wu, J.; Zeng, Z.; Wang, Y.; Si, X.; et al. Phytochemical profiles of rice and their cellular antioxidant activity against ABAP induced oxidative stress in human hepatocellular carcinoma HepG2 cells. Food Chem. 2020, 318, 126484. [Google Scholar] [CrossRef]
- Li, Y.; Li, T.; Liu, R.H. Bioactive compounds of highland barley and their health benefits. J. Cereal Sci. 2022, 103, 103366. [Google Scholar] [CrossRef]
- Ilavenil, S.; Kim, D.H.; Srigopalram, S.; Arasu, M.V.; Lee, K.D.; Lee, J.C.; Lee, J.S.; Renganathan, S.; Choi, K.C. Potential Application of p-Coumaric Acid on Differentiation of C2C12 Skeletal Muscle and 3T3-L1 Preadipocytes-An In Vitro and In Silico Approach. Molecules 2016, 21, 997. [Google Scholar] [CrossRef]
- Yoon, S.-A.; Kang, S.-I.; Shin, H.-S.; Kang, S.-W.; Kim, J.-H.; Ko, H.-C.; Kim, S.-J. p-Coumaric acid modulates glucose and lipid metabolism via AMP-activated protein kinase in L6 skeletal muscle cells. Biochem. Biophys. Res. Commun. 2013, 432, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Nemcova, L.; Zima, J.; Barek, J.; Janovska, D. Determination of resveratrol in grains, hulls and leaves of common and tartary buckwheat by HPLC with electrochemical detection at carbon paste electrode. Food Chem. 2011, 126, 374–378. [Google Scholar] [CrossRef]
- Liu, J.; Peng, Y.; Wang, X.; Fan, Y.; Qin, C.; Shi, L.; Tang, Y.; Cao, K.; Li, H.; Long, J.; et al. Mitochondrial Dysfunction Launches Dexamethasone-Induced Skeletal Muscle Atrophy via AMPK/FOXO3 Signaling. Mol. Pharm. 2016, 13, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhu, X.; Chen, K.; Lang, H.; Zhang, Y.; Hou, P.; Ran, L.; Zhou, M.; Zheng, J.; Yi, L.; et al. Resveratrol prevents sarcopenic obesity by reversing mitochondrial dysfunction and oxidative stress via the PKA/LKB1/AMPK pathway. Aging-Us 2019, 11, 2217–2240. [Google Scholar] [CrossRef]
- Niu, W.; Wang, H.; Wang, B.; Mao, X.; Du, M. Resveratrol improves muscle regeneration in obese mice through enhancing mitochondrial biogenesis. J. Nutr. Biochem. 2021, 98, 108804. [Google Scholar] [CrossRef]
- Gong, L.; Guo, S.; Zou, Z. Resveratrol ameliorates metabolic disorders and insulin resistance in high-fat diet-fed mice. Life Sci. 2020, 242, 117212. [Google Scholar] [CrossRef]
- Quan, Y.; Hua, S.; Li, W.; Zhan, M.; Li, Y.; Lu, L. Resveratrol bidirectionally regulates insulin effects in skeletal muscle through alternation of intracellular redox homeostasis. Life Sci. 2020, 242, 117188. [Google Scholar] [CrossRef]
- Wang, D.-T.; Yin, Y.; Yang, Y.-J.; Lv, P.-J.; Shi, Y.; Lu, L.; Wei, L.-B. Resveratrol prevents TNF-alpha-induced muscle atrophy via regulation of Akt/mTOR/FoxO1 signaling in C2C12 myotubes. Int. Immunopharmacol. 2014, 19, 206–213. [Google Scholar] [CrossRef]
- Jiang, Q.; Cheng, X.; Cui, Y.; Xia, Q.; Yan, X.; Zhang, M.; Lan, G.; Liu, J.; Shan, T.; Huang, Y. Resveratrol regulates skeletal muscle fibers switching through the AdipoR1-AMPK-PGC-1 alpha pathway. Food Funct. 2019, 10, 3334–3343. [Google Scholar] [CrossRef]
- Wen, W.; Chen, X.; Huang, Z.; Chen, D.; Chen, H.; Luo, Y.; He, J.; Zheng, P.; Yu, J.; Yu, B. Resveratrol regulates muscle fiber type conversion via miR-22-3p and AMPK/SIRT1/PGC-1 alpha pathway. J. Nutr. Biochem. 2020, 77, 108297. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-J.; Ho, C.-S.; Lee, M.-C.; Ho, C.-S.; Huang, C.-C.; Kan, N.-W. Protective Effects of Resveratrol Supplementation on Contusion Induced Muscle Injury. Int. J. Med. Sci. 2020, 17, 53–62. [Google Scholar] [CrossRef]
- Gabr, A.M.M.; Fayek, N.M.; Mahmoud, H.M.; El-Bahr, M.K.; Ebrahim, H.S.; Sytar, O.; El-Halawany, A.M. Effect of Light Quality and Media Components on Shoot Growth, Rutin, and Quercetin Production from Common Buckwheat. ACS Omega 2022, 7, 26566–26572. [Google Scholar] [CrossRef]
- Paulina Luna-Castillo, K.; Lin, S.; Francisco Munoz-Valle, J.; Vizmanos, B.; Lopez-Quintero, A.; Marquez-Sandoval, F. Functional Food and Bioactive Compounds on the Modulation of the Functionality of HDL-C: A Narrative Review. Nutrients 2021, 13, 1165. [Google Scholar] [CrossRef]
- Chen, X.; Liang, D.; Huang, Z.; Jia, G.; Zhao, H.; Liu, G. Quercetin regulates skeletal muscle fiber type switching via adiponectin signaling. Food Funct. 2021, 12, 2693–2702. [Google Scholar] [CrossRef] [PubMed]
- Iwabu, M.; Yamauchi, T.; Okada-Iwabu, M.; Sato, K.; Nakagawa, T.; Funata, M.; Yamaguchi, M.; Namiki, S.; Nakayama, R.; Tabata, M.; et al. Adiponectin and AdipoR1 regulate PGC-1 alpha and mitochondria by Ca2+ and AMPK/SIRT1. Nature 2010, 464, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, C.-S.; Joe, Y.; Chung, H.T.; Ha, T.Y.; Yu, R. Quercetin Reduces Tumor Necrosis Factor Alpha-Induced Muscle Atrophy by Upregulation of Heme Oxygenase-1. J. Med. Food 2018, 21, 551–559. [Google Scholar] [CrossRef]
- Mukai, R.; Matsui, N.; Fujikura, Y.; Matsumoto, N.; Hou, D.-X.; Kanzaki, N.; Shibata, H.; Horikawa, M.; Iwasa, K.; Hirasaka, K.; et al. Preventive effect of dietary quercetin on disuse muscle atrophy by targeting mitochondria in denervated mice. J. Nutr. Biochem. 2016, 31, 67–76. [Google Scholar] [CrossRef]
- Dai, X.; Ding, Y.; Zhang, Z.; Cai, X.; Bao, L.; Li, Y. Quercetin But Not Quercitrin Ameliorates Tumor Necrosis Factor-Alpha-Induced Insulin Resistance in C2C12 Skeletal Muscle Cells. Biol. Pharm. Bull. 2013, 36, 788–795. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Yang, J.-S.; Lu, C.-C.; Chiu, Y.-J.; Chen, H.-C.; Chung, M.-I.; Wu, Y.-T.; Chen, F.-A. Effect of Quercetin on Dexamethasone-Induced C2C12 Skeletal Muscle Cell Injury. Molecules 2020, 25, 3267. [Google Scholar] [CrossRef]
- Liang, Q.; Chen, Y.; Li, C.; Lu, L. Quercetin attenuates Ox-LDL-induced calcification in vascular smooth muscle cells by regulating ROS-TLR4 signaling pathway. Nan Fang Yi Ke Da Xue Xue Bao J. South. Med. Univ. 2018, 38, 980–985. [Google Scholar] [CrossRef]
- Chen, X.; Liang, D.; Huang, Z.; Jia, G.; Zhao, H.; Liu, G. Anti-fatigue effect of quercetin on enhancing muscle function and antioxidant capacity. J. Food Biochem. 2021, 45, e13968. [Google Scholar] [CrossRef] [PubMed]
- Finocchiaro, F.; Ferrari, B.; Gianinetti, A.; Dall’Asta, C.; Galaverna, G.; Scazzina, F.; Pellegrini, N. Characterization of antioxidant compounds of red and white rice and changes in total antioxidant capacity during processing. Mol. Nutr. Food Res. 2007, 51, 1006–1019. [Google Scholar] [CrossRef]
- Li, X.; Sui, Y.; Wu, Q.; Xie, B.; Sun, Z. Attenuated mTOR Signaling and Enhanced Glucose Homeostasis by Dietary Supplementation with Lotus Seedpod Oligomeric Procyanidins in Streptozotocin (STZ)-Induced Diabetic Mice. J. Agric. Food Chem. 2017, 65, 3801–3810. [Google Scholar] [CrossRef]
- Bowser, S.M.; Moore, W.T.; McMillan, R.P.; Dorenkott, M.R.; Goodrich, K.M.; Ye, L.; O’Keefe, S.F.; Hulver, M.W.; Neilson, A.P. High-molecular-weight cocoa procyanidins possess enhanced insulin-enhancing and insulin mimetic activities in human primary skeletal muscle cells compared to smaller procyanidins. J. Nutr. Biochem. 2017, 39, 48–58. [Google Scholar] [CrossRef]
- Yamashita, Y.; Wang, L.; Nanba, F.; Ito, C.; Toda, T.; Ashida, H. Procyanidin Promotes Translocation of Glucose Transporter 4 in Muscle of Mice through Activation of Insulin and AMPK Signaling Pathways. PLoS ONE 2016, 11, e0161704. [Google Scholar] [CrossRef] [PubMed]
- Corol, D.-I.; Ravel, C.; Raksegi, M.; Bedo, Z.; Charmet, G.; Beale, M.H.; Shewry, P.R.; Ward, J.L. Effects of Genotype and Environment on the Contents of Betaine, Choline, and Trigonelline in Cereal Grains. J. Agric. Food Chem. 2012, 60, 5471–5481. [Google Scholar] [CrossRef]
- Guo, H.; Guo, J.; Jiang, X.; Li, Z.; Ling, W. Cyanidin-3-O-beta-glucoside, a typical anthocyanin, exhibits antilipolytic effects in 3T3-L1 adipocytes during hyperglycemia: Involvement of FoxO1-mediated transcription of adipose triglyceride lipase. Food Chem. Toxicol. 2012, 50, 3040–3047. [Google Scholar] [CrossRef]
- Matsukawa, T.; Inaguma, T.; Han, J.; Villareal, M.O.; Isoda, H. Cyanidin-3-glucoside derived from black soybeans ameliorate type 2 diabetes through the induction of differentiation of preadipocytes into smaller and insulin-sensitive adipocytes. J. Nutr. Biochem. 2015, 26, 860–867. [Google Scholar] [CrossRef]
- Giang Thanh Thi, H.; Kase, E.T.; Wangensteen, H.; Barsett, H. Phenolic Elderberry Extracts, Anthocyanins, Procyanidins, and Metabolites Influence Glucose and Fatty Acid Uptake in Human Skeletal Muscle Cells. J. Agric. Food Chem. 2017, 65, 2677–2685. [Google Scholar] [CrossRef]
- Matsukawa, T.; Motojima, H.; Sato, Y.; Takahashi, S.; Villareal, M.O.; Isoda, H. Upregulation of skeletal muscle PGC-1 alpha through the elevation of cyclic AMP levels by Cyanidin-3-glucoside enhances exercise performance. Sci. Rep. 2017, 7, 44799. [Google Scholar] [CrossRef] [PubMed]
- Dinelli, G.; Segura Carretero, A.; Di Silvestro, R.; Marotti, I.; Fu, S.; Benedettelli, S.; Ghiselli, L.; Fernandez Gutierrez, A. Determination of phenolic compounds in modern and old varieties of durum wheat using liquid chromatography coupled with time-of-flight mass spectrometry. J. Chromatogr. A 2009, 1216, 7229–7240. [Google Scholar] [CrossRef]
- Hes, M.; Dziedzic, K.; Gorecka, D.; Drozdzynska, A.; Gujska, E. Effect of Boiling in Water of Barley and Buckwheat Groats on the Antioxidant Properties and Dietary Fiber Composition. Plant Foods Hum. Nutr. 2014, 69, 276–282. [Google Scholar] [CrossRef]
- Li, P.; Liu, A.; Xiong, W.; Lin, H.; Xiao, W.; Huang, J.; Zhang, S.; Liu, Z. Catechins enhance skeletal muscle performance. Crit. Rev. Food Sci. Nutr. 2020, 60, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Kim, K.M.; Byun, M.R.; Hwang, J.-H.; Park, J.I.; Oh, H.T.; Kim, H.K.; Jeong, M.G.; Hwang, E.S.; Hong, J.-H. Catechins activate muscle stem cells by Myf5 induction and stimulate muscle regeneration. Biochem. Biophys. Res. Commun. 2017, 489, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, A.; Liu, C.; Qu, Z.; Xiao, W.; Huang, J.; Liu, Z.; Zhang, S. Role and mechanism of catechin in skeletal muscle cell differentiation. J. Nutr. Biochem. 2019, 74, 108225–108260. [Google Scholar] [CrossRef]
- Huettemann, M.; Lee, I.; Perkins, G.A.; Britton, S.L.; Koch, L.G.; Malek, M.H. (-)-Epicatechin is associated with increased angiogenic and mitochondrial signalling in the hindlimb of rats selectively bred for innate low running capacity. Clin. Sci. 2013, 124, 663–674. [Google Scholar] [CrossRef]
- Yan, J.; Feng, Z.; Liu, J.; Shen, W.; Wang, Y.; Wertz, K.; Weber, P.; Long, J.; Liu, J. Enhanced autophagy plays a cardinal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats: Ameliorating effects of (-)-epigallocatechin-3-gallate. J. Nutr. Biochem. 2012, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Kim, K.M.; Byun, M.R.; Hwang, J.-H.; Park, J.I.; Oh, H.T.; Jeong, M.G.; Hwang, E.S.; Hong, J.-H. (-)-Epigallocatechin-3-gallate stimulates myogenic differentiation through TAZ activation. Biochem. Biophys. Res. Commun. 2017, 486, 378–384. [Google Scholar] [CrossRef]
- Meador, B.M.; Mirza, K.A.; Tian, M.; Skelding, M.B.; Reaves, L.A.; Edens, N.K.; Tisdale, M.J.; Pereira, S.L. The Green Tea Polyphenol Epigallocatechin-3-Gallate (EGCg) Attenuates Skeletal Muscle Atrophy in a Rat Model of Sarcopenia. J. Frailty Aging 2015, 4, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T.; Hayashi, H.; Fujimaki, N.; Nishizawa, N.; Kitts, D.D. Induction of oxidatively modified proteins in skeletal muscle by electrical stimulation and its suppression by dietary supplementation of (-)-epigallocatechin gallate. Biosci. Biotechnol. Biochem. 2000, 64, 1004–1010. [Google Scholar] [CrossRef]
- Li, Y.; Ma, D.; Sun, D.; Wang, C.; Zhang, J.; Xie, Y.; Guo, T. Total phenolic, flavonoid content, and antioxidant activity of flour, noodles, and steamed bread made from different colored wheat grains by three milling methods. Crop J. 2015, 3, 328–334. [Google Scholar] [CrossRef]
- Maria Gomez-Caravaca, A.; Segura-Carretero, A.; Fernandez-Gutierrez, A.; Caboni, M.F. Simultaneous Determination of Phenolic Compounds and Saponins in Quinoa (Chenopodium quinoa Willd) by a Liquid Chromatography-Diode Array Detection-Electrospray Ionization-Time-of-Flight Mass Spectrometry Methodology. J. Agric. Food Chem. 2011, 59, 10815–10825. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.R.; Hsiao, G.; Chou, P.H.; Shen, M.Y.; Chou, D.S. Mechanisms involved in the antiplatelet activity of rutin, a glycoside of the flavonol quercetin, in human platelets. J. Agric. Food Chem. 2004, 52, 4414–4418. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Hickner, R.C.; Cortright, R.L.; Dohm, G.L.; Houmard, J.A. Lipid oxidation is reduced in obese human skeletal muscle. Am. J. Physiol.-Endocrinol. Metab. 2000, 279, E1039–E1044. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Lee, M.-S.; Chang, E.; Shin, Y.; Oh, S.; Kim, I.-H.; Kim, Y. Rutin Increases Muscle Mitochondrial Biogenesis with AMPK Activation in High-Fat Diet-Induced Obese Rats. Nutrients 2015, 7, 8152–8169. [Google Scholar] [CrossRef]
- Su, K.-Y.; Yu, C.Y.; Chen, Y.-W.; Huang, Y.-T.; Chen, C.-T.; Wu, H.-F.; Chen, Y.-L.S. Rutin, a Flavonoid and Principal Component of Saussurea Involucrata, Attenuates Physical Fatigue in a Forced Swimming Mouse Model. Int. J. Med. Sci. 2014, 11, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Adewole, D.; Yu, L.; Sid, V.; Wang, B.; Karmin, O.; Yang, C. Rutin attenuates inflammatory responses induced by lipopolysaccharide in an in vitro mouse muscle cell (C2C12) model. Poult. Sci. 2019, 98, 2756–2764. [Google Scholar] [CrossRef]
- Weber, D.; Grune, T. The contribution of beta-carotene to vitamin A supply of humans. Mol. Nutr. Food Res. 2012, 56, 251–258. [Google Scholar] [CrossRef]
- Banwo, K.; Olojede, A.O.; Adesulu-Dahunsi, A.T.; Verma, D.K.; Thakur, M.; Tripathy, S.; Singh, S.; Patel, A.R.; Gupta, A.K.; Aguilar, C.N.; et al. Functional importance of bioactive compounds of foods with Potential Health Benefits: A review on recent trends. Food Biosci. 2021, 43, 101320. [Google Scholar] [CrossRef]
- Loskutov, I.G.; Khlestkina, E.K. Wheat, Barley, and Oat Breeding for Health Benefit Components in Grain. Plants 2021, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Zhang, Q.; Yan, F.-F.; Wan, J.-F.; Lin, C.-S. Lutein protects against ischemia/reperfusion injury in rat skeletal muscle by modulating oxidative stress and inflammation. Immunopharmacol. Immunotoxicol. 2015, 37, 329–334. [Google Scholar] [CrossRef]
- Nagao, A. Metabolism of Carotenoids in Mammals. In Carotenoids: Biosynthetic and Biofunctional Approaches; Misawa, N., Ed.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2021; Volume 1261, pp. 67–78. [Google Scholar]
- Kitakaze, T.; Harada, N.; Imagita, H.; Yamaji, R. beta-Carotene Increases Muscle Mass and Hypertrophy in the Soleus Muscle in Mice. J. Nutr. Sci. Vitaminol. 2015, 61, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Foletta, V.C.; White, L.J.; Larsen, A.E.; Leger, B.; Russell, A.P. The role and regulation of MAFbx/atrogin-1 and MuRF1 in skeletal muscle atrophy. Pflug. Arch. -Eur. J. Physiol. 2011, 461, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Kariya, Y.; Kitakaze, T.; Yamaji, R.; Harada, N.; Sakamoto, T.; Hosotani, K.; Nakano, Y.; Inui, H. The preventive effect of beta-carotene on denervation-induced soleus muscle atrophy in mice. Br. J. Nutr. 2013, 109, 1349–1358. [Google Scholar] [CrossRef]
- Siurek, B.; Rosicka-Kaczmarek, J.; Nebesny, E. Bioactive compounds in cereal grains—Occurrence, structure, technological significance and nutritional benefits-a review. Food Sci. Technol. Int. 2012, 18, 559–568. [Google Scholar] [CrossRef]
- Borneo, R.; Leon, A.E. Whole grain cereals: Functional components and health benefits. Food Funct. 2012, 3, 110–119. [Google Scholar] [CrossRef]
- Lee, H.; Lim, Y. Tocotrienol-rich fraction supplementation reduces hyperglycemia-induced skeletal muscle damage through regulation of insulin signaling and oxidative stress in type 2 diabetic mice. J. Nutr. Biochem. 2018, 57, 77–85. [Google Scholar] [CrossRef]
- Razak, A.M.; Khor, S.C.; Jaafar, F.; Karim, N.A.; Makpol, S. Targeting myomiRs by tocotrienol-rich fraction to promote myoblast differentiation. Genes Nutr. 2018, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.J.; Zurinah, W.N.W.; Moul, V.; Norwahidah, A.K. Tocotrienol-Rich Fraction (TRF) Treatment Promotes Proliferation Capacity of Stress-Induced Premature Senescence Myoblasts and Modulates the Renewal of Satellite Cells: Microarray Analysis. Oxidative Med. Cell. Longev. 2019, 2019, 9141343. [Google Scholar] [CrossRef]
- Chung, E.; Campise, S.N.; Joiner, H.E.; Tomison, M.D.; Kaur, G.; Dufour, J.M.; Cole, L.; Ramalingam, L.; Moustaid-Moussa, N.; Shen, C.-L. Effect of annatto-extracted tocotrienols and green tea polyphenols on glucose homeostasis and skeletal muscle metabolism in obese male mice. J. Nutr. Biochem. 2019, 67, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.-D.; Shin, G.-H.; Kim, J.-M.; Choi, S.-I.; Lee, J.-H.; Lee, S.J.; Park, S.J.; Woo, K.S.; Oh, S.K.; Lee, O.-H. Comparative Analysis of gamma-Oryzanol, beta-Glucan, Total Phenolic Content and Antioxidant Activity in Fermented Rice Bran of Different Varieties. Nutrients 2017, 9, 571. [Google Scholar] [CrossRef]
- Prasadi, V.P.N.; Joye, I.J. Dietary Fibre from Whole Grains and Their Benefits on Metabolic Health. Nutrients 2020, 12, 3045. [Google Scholar] [CrossRef]
- Xu, C.; Lv, J.; Lo, Y.M.; Cui, S.W.; Hu, X.; Fan, M. Effects of oat beta-glucan on endurance exercise and its anti-fatigue properties in trained rats. Carbohydr. Polym. 2013, 92, 1159–1165. [Google Scholar] [CrossRef]
- Li, Y.; Fan, Y.; Pan, H.; Qian, H.; Qi, X.; Wu, G.; Zhang, H.; Xu, M.; Rao, Z.; Wang, L.; et al. Effects of functional beta-glucan on proliferation, differentiation, metabolism and its anti-fibrosis properties in muscle cells. Int. J. Biol. Macromol. 2018, 117, 287–293. [Google Scholar] [CrossRef]
- Brogi, L.; Marchese, M.; Cellerino, A.; Licitra, R.; Naef, V.; Mero, S.; Bibbiani, C.; Fronte, B. beta-Glucans as Dietary Supplement to Improve Locomotion and Mitochondrial Respiration in a Model of Duchenne Muscular Dystrophy. Nutrients 2021, 13, 1619. [Google Scholar] [CrossRef] [PubMed]
- Ratseewo, J.; Meeso, N.; Siriamornpun, S. Changes in amino acids and bioactive compounds of pigmented rice as affected by far-infrared radiation and hot air drying. Food Chem. 2020, 306, 125644. [Google Scholar] [CrossRef]
- Zhu, D.; Sanchez-Ferrer, A.; Nystroem, L. Antioxidant Activity of Individual Steryl Ferulates from Various Cereal Grain Sources. J. Nat. Prod. 2016, 79, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Son, H.J.; Seo, H.D.; Ha, T.Y.; Ahn, J.; Lee, H.; Shin, S.H.; Jung, C.H.; Jang, Y.J. gamma-Oryzanol Improves Exercise Endurance and Muscle Strength by Upregulating PPAR delta and ERR gamma Activity in Aged Mice. Mol. Nutr. Food Res. 2021, 65, 2000652. [Google Scholar] [CrossRef] [PubMed]
- Mattei, L.; Francisqueti-Ferron, F.V.; Garcia, J.L.; Ferron, A.J.T.; Silva, C.C.V.d.A.; Gregolin, C.S.; Nakandakare-Maia, E.T.; Silva, J.d.C.P.; Moreto, F.; Minatel, I.O.; et al. Antioxidant and anti-inflammatory properties of gamma- oryzanol attenuates insulin resistance by increasing GLUT-4 expression in skeletal muscle of obese animals. Mol. Cell. Endocrinol. 2021, 537, 111423. [Google Scholar] [CrossRef]
- Dahleh, M.M.M.; Araujo, S.M.; Bortolotto, V.C.; Pinheiro, F.C.; Poetini, M.R.; Musachio, E.A.S.; Meichtry, L.B.; Couto, S.d.F.; Prigol, M. Exercise associated with gamma-oryzanol supplementation suppresses oxidative stress and prevents changes in locomotion in Drosophila melanogaster. Free. Radic. Res. 2021, 55, 198–209. [Google Scholar] [CrossRef]
- Chen, C.Y.O.; Kamil, A.; Blumberg, J.B. Phytochemical composition and antioxidant capacity of whole wheat products. Int. J. Food Sci. Nutr. 2015, 66, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Junaid, M.; Ullah, F.; Subhan, F.; Sadiq, A.; Ali, G.; Ovais, M.; Shahid, M.; Ahmad, A.; Wadood, A.; et al. Anti-Alzheimer’s Studies on beta-Sitosterol Isolated from Polygonum hydropiper L. Front. Pharmacol. 2017, 8, 697. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.S.; Leong, P.K.; Chen, J.; Leung, H.Y.; Chan, W.M.; Ko, K.M. beta-Sitosterol increases mitochondrial electron transport by fluidizing mitochondrial membranes and enhances mitochondrial responsiveness to increasing energy demand by the induction of uncoupling in C2C12 myotubes. J. Funct. Foods 2016, 23, 253–260. [Google Scholar] [CrossRef]
- Cheng, Y.; Chen, Y.; Li, J.; Qu, H.; Zhao, Y.; Wen, C.; Zhou, Y. Dietary beta-Sitosterol Improves Growth Performance, Meat Quality, Antioxidant Status, and Mitochondrial Biogenesis of Breast Muscle in Broilers. Animals 2019, 9, 71. [Google Scholar] [CrossRef]
- Hwang, S.-L.; Kim, H.-N.; Jung, H.-H.; Kim, J.-E.; Choi, D.-K.; Hur, J.-M.; Lee, J.-Y.; Song, H.; Song, K.-S.; Huh, T.-L. Beneficial effects of beta-sitosterol on glucose and lipid metabolism in L6 myotube cells are mediated by AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2008, 377, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Prasad, M.; Mohamed Helal, G.; El-Sherbiny, M.; Abdelmonem Elsherbini, D.M.; Rajagopal, P.; Palanisamy, C.P.; Veeraraghavan, V.P.; Jayaraman, S.; Surapaneni, K.M. Beta-Sitosterol Facilitates GLUT4 Vesicle Fusion on the Plasma Membrane via the Activation of Rab/IRAP/Munc 18 Signaling Pathways in Diabetic Gastrocnemius Muscle of Adult Male Rats. Bioinorg. Chem. Appl. 2022, 2022, 7772305. [Google Scholar] [CrossRef] [PubMed]
- Dianita, R.; Jantan, I.; Amran, A.Z.; Jalil, J. Protective Effects of Labisia pumila var. alata on Biochemical and Histopathological Alterations of Cardiac Muscle Cells in Isoproterenol-Induced Myocardial Infarction Rats. Molecules 2015, 20, 4746–4763. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Q.; Hao, Y.; Wang, Z.; Yang, H.; Liu, J.; Wang, J. Protective effect of 5-heptadecylresorcinol against obesity-associated skeletal muscle dysfunction by modulating mitochondrial biogenesis via the activation of SIRT3/PGC-1α signaling pathway. J. Funct. Foods 2022, 95, 105178. [Google Scholar] [CrossRef]
- Filipcev, B.; Kojic, J.; Krulj, J.; Bodroza-Solarov, M.; Ilic, N. Betaine in Cereal Grains and Grain-Based Products. Foods 2018, 7, 49. [Google Scholar] [CrossRef]
- Ismaeel, A. Effects of betaine supplementation on muscle strength and power: A systematic review. J. Strength Cond. Res. 2017, 31, 2338–2346. [Google Scholar] [CrossRef] [PubMed]
- Senesi, P.; Luzi, L.; Montesano, A.; Mazzocchi, N.; Terruzzi, I. Betaine supplement enhances skeletal muscle differentiation in murine myoblasts via IGF-1 signaling activation. J. Transl. Med. 2013, 11, 174. [Google Scholar] [CrossRef]
- Apicella, J.M.; Lee, E.C.; Bailey, B.L.; Saenz, C.; Anderson, J.M.; Craig, S.A.S.; Kraemer, W.J.; Volek, J.S.; Maresh, C.M. Betaine supplementation enhances anabolic endocrine and Akt signaling in response to acute bouts of exercise. Eur. J. Appl. Physiol. 2013, 113, 793–802. [Google Scholar] [CrossRef]
- Du, J.; Shen, L.; Zhang, P.; Tan, Z.; Cheng, X.; Luo, J.; Zhao, X.; Yang, Q.; Gu, H.; Jiang, A.a.; et al. The regulation of skeletal muscle fiber-type composition by betaine is associated with NFATc1/MyoD. J. Mol. Med. 2018, 96, 685–700. [Google Scholar] [CrossRef]
- Ma, J.; Meng, X.; Kang, S.Y.; Zhang, J.; Jung, H.W.; Park, Y.-K. Regulatory effects of the fruit extract of Lycium chinense and its active compound, betaine, on muscle differentiation and mitochondrial biogenesis in C2C12 cells. Biomed. Pharmacother. 2019, 118, 109297. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.M.; Lee, M.J.; Yoon, S.H.; Jung, M.Y. A Gas Chromatography-Tandem Quadrupole Mass Spectrometric Analysis of Policosanols in Commercial Vegetable Oils. J. Food Sci. 2011, 76, C891–C899. [Google Scholar] [CrossRef]
- Irmak, S.; Dunford, N.T. Policosanol contents and compositions of wheat varieties. J. Agric. Food Chem. 2005, 53, 5583–5586. [Google Scholar] [CrossRef]
- Kabir, Y.; Kimura, S. Tissue distribution of (8-14C)-octacosanol in liver and muscle of rats after serial administration. Ann. Nutr. Metab. 1995, 39, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, S.; Han, D.S.; Park, T. Octacosanol supplementation increases running endurance time and improves biochemical parameters after exhaustion in trained rats. J. Med. Food 2003, 6, 345–351. [Google Scholar] [CrossRef]
- Kabir, Y.; Kimura, S. Biodistribution and metabolism of orally administered octacosanol in rats. Ann. Nutr. Metab. 1993, 37, 33–38. [Google Scholar] [CrossRef]
- Zhou, Y.; Cao, F.; Wu, Q.; Luo, Y.; Guo, T.; Han, S.; Huang, M.; Hu, Z.; Bai, J.; Luo, F.; et al. Dietary Supplementation of Octacosanol Improves Exercise-Induced Fatigue and Its Molecular Mechanism. J. Agric. Food Chem. 2021, 69, 7603–7618. [Google Scholar] [CrossRef] [PubMed]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Higher dietary fibre intake is associated with increased skeletal muscle mass and strength in adults aged 40 years and older. J. Cachexia Sarcopenia Muscle 2021, 12, 2134–2144. [Google Scholar] [CrossRef] [PubMed]
- Kaviani, M.; Chilibeck, P.D.; Jochimi, J.; Gordon, J.; Zello, G.A. The Glycemic Index of Sport Nutrition Bars Affects Performance and Metabolism During Cycling and Next-Day Recovery. J. Hum. Kinet. 2019, 66, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, B.; Zahiragic, L.; Bierwagen, A.; Kabisch, S.; Groener, J.B.; Nowotny, P.J.; Fleitmann, A.K.; Herder, C.; Pacini, G.; Erlund, I.; et al. Low-energy diets differing in fibre, red meat and coffee intake equally improve insulin sensitivity in type 2 diabetes: A randomised feasibility trial. Diabetologia 2015, 58, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Lee, S. Dietary Patterns Related to Appendicular Skeletal Muscle Mass: The Korea National Health and Nutrition Examination Survey 2008-2011. J. Am. Coll. Nutr. 2019, 38, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Mafi, F.; Biglari, S.; Afousi, A.G.; Gaeini, A.A. Improvement in Skeletal Muscle Strength and Plasma Levels of Follistatin and Myostatin Induced by an 8-Week Resistance Training and Epicatechin Supplementation in Sarcopenic Older Adults. J. Aging Phys. Act. 2019, 27, 384–391. [Google Scholar] [CrossRef]
- Taub, P.R.; Ramirez-Sanchez, I.; Ciaraldi, T.P.; Perkins, G.; Murphy, A.N.; Naviaux, R.; Hogan, M.; Maisel, A.S.; Henry, R.R.; Ceballos, G.; et al. Alterations in Skeletal Muscle Indicators of Mitochondrial Structure and Biogenesis in Patients with Type 2 Diabetes and Heart Failure: Effects of Epicatechin Rich Cocoa. Clin. Transl. Sci. 2012, 5, 43–47. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Villani, A.; Wright, H.; Slater, G.; Buckley, J. A randomised controlled intervention study investigating the efficacy of carotenoid-rich fruits and vegetables and extra-virgin olive oil on attenuating sarcopenic symptomology in overweight and obese older adults during energy intake restriction: Protocol paper. BMC Geriatr. 2018, 18, 2. [Google Scholar] [CrossRef] [Green Version]
- Semba, R.D.; Lauretani, F.; Ferrucci, L. Carotenoids as protection against sarcopenia in older adults. Arch. Biochem. Biophys. 2007, 458, 141–145. [Google Scholar] [CrossRef]
- Harari, A.; Coster, A.C.F.; Jenkins, A.; Xu, A.; Greenfield, J.R.; Harats, D.; Shaish, A.; Samocha-Bonet, D. Obesity and Insulin Resistance Are Inversely Associated with Serum and Adipose Tissue Carotenoid Concentrations in Adults. J. Nutr. 2020, 150, 38–46. [Google Scholar] [CrossRef]
- Long, J.-A.; Zhong, R.-H.; Chen, S.; Wang, F.; Luo, Y.; Lu, X.-T.; Yishake, D.; Chen, Y.-M.; Fang, A.-P.; Zhu, H.-L. Dietary betaine intake is associated with skeletal muscle mass change over 3 years in middle-aged adults: The Guangzhou Nutrition and Health Study. Br. J. Nutr. 2021, 125, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Arazi, H.; Aboutalebi, S.; Taati, B.; Cholewa, J.M.; Candow, D.G. Effects of short-term betaine supplementation on muscle endurance and indices of endocrine function following acute high-intensity resistance exercise in young athletes. J. Int. Soc. Sports Nutr. 2022, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cholewa, J.M.; Wyszczelska-Rokiel, M.; Glowacki, R.; Jakubowski, H.; Matthews, T.; Wood, R.; Craig, S.A.S.; Paolone, V. Effects of betaine on body composition, performance, and homocysteine thiolactone. J. Int. Soc. Sports Nutr. 2013, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Hah, Y.-S.; Lee, W.K.; Lee, S.; Kim, E.J.; Lee, J.H.; Lee, S.-J.; Ji, Y.H.; Kim, S.G.; Lee, H.-H.; Hong, S.Y.; et al. beta-Sitosterol Attenuates Dexamethasone-Induced Muscle Atrophy via Regulating FoxO1-Dependent Signaling in C2C12 Cell and Mice Model. Nutrients 2022, 14, 2894. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Yang, H.; Song, S.; Liu, J.; Wang, Z.; Wang, J. Bioactive Components in Whole Grains for the Regulation of Skeletal Muscle Function. Foods 2022, 11, 2752. https://doi.org/10.3390/foods11182752
Li Q, Yang H, Song S, Liu J, Wang Z, Wang J. Bioactive Components in Whole Grains for the Regulation of Skeletal Muscle Function. Foods. 2022; 11(18):2752. https://doi.org/10.3390/foods11182752
Chicago/Turabian StyleLi, Qing, Haihong Yang, Shuimiao Song, Jie Liu, Ziyuan Wang, and Jing Wang. 2022. "Bioactive Components in Whole Grains for the Regulation of Skeletal Muscle Function" Foods 11, no. 18: 2752. https://doi.org/10.3390/foods11182752
APA StyleLi, Q., Yang, H., Song, S., Liu, J., Wang, Z., & Wang, J. (2022). Bioactive Components in Whole Grains for the Regulation of Skeletal Muscle Function. Foods, 11(18), 2752. https://doi.org/10.3390/foods11182752