Role of Innate Inflammation in the Regulation of Tissue Remodeling during Tooth Eruption


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Preparation
2.3. Immunohistochemistry
2.4. Quantitative Real-Time PCR Analysis
2.5. Statistical Analysis
3. Results
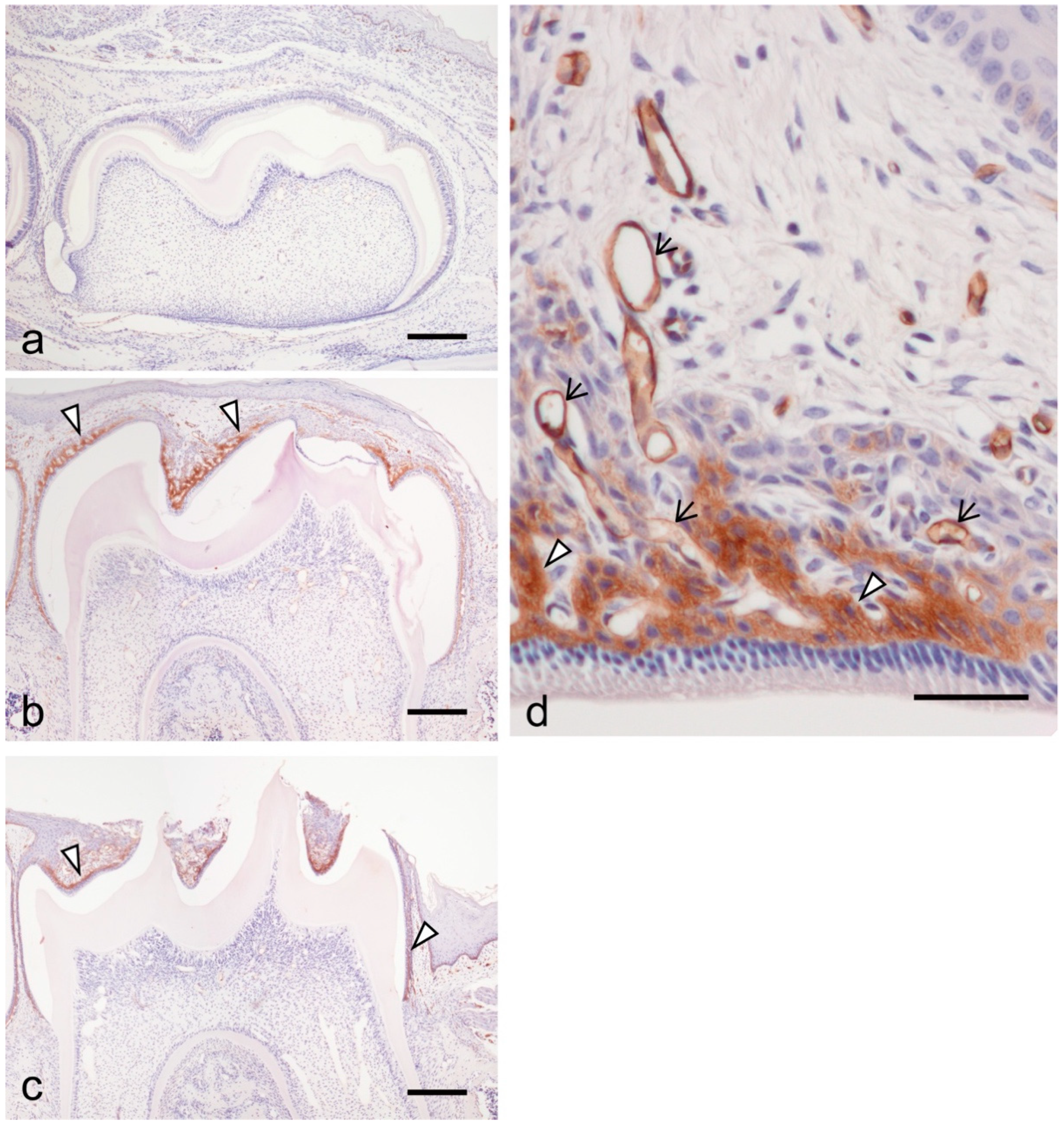
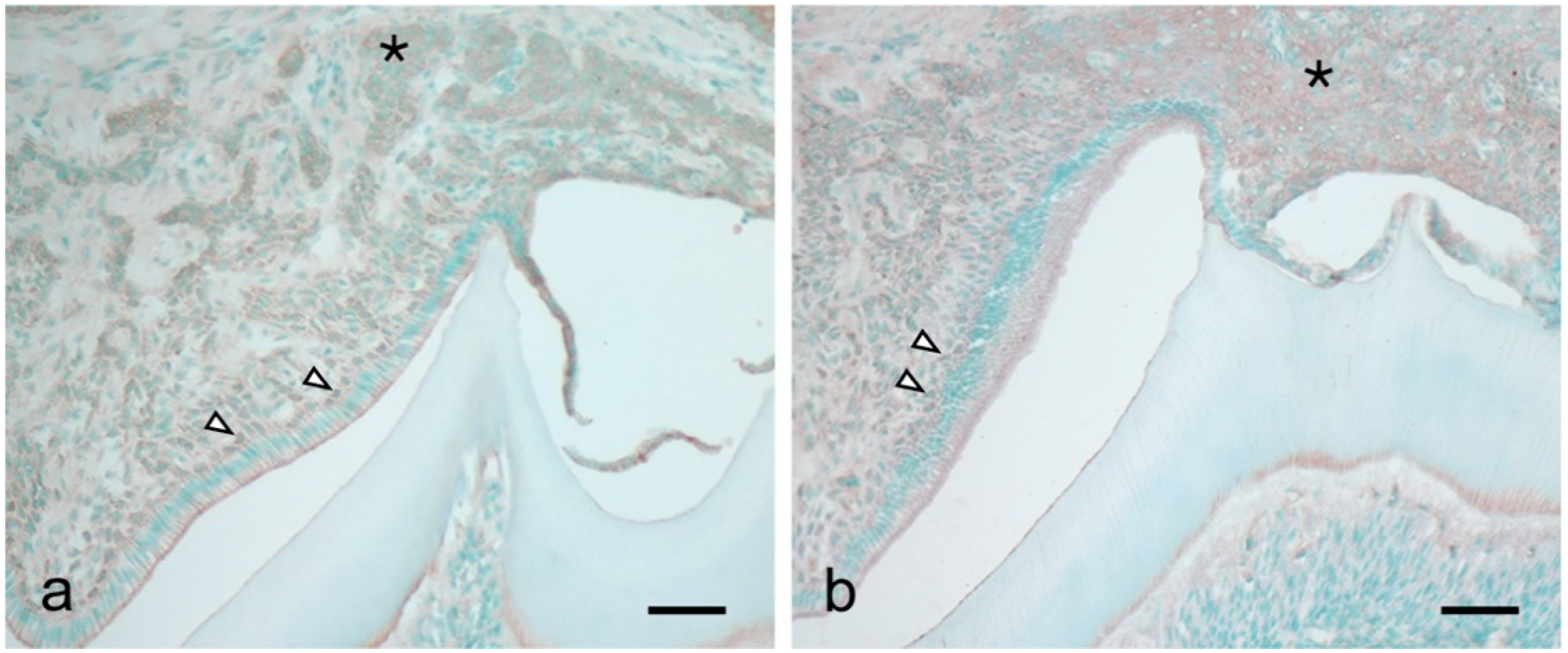
3.1. ICAM-1 Expression during Tooth Eruption
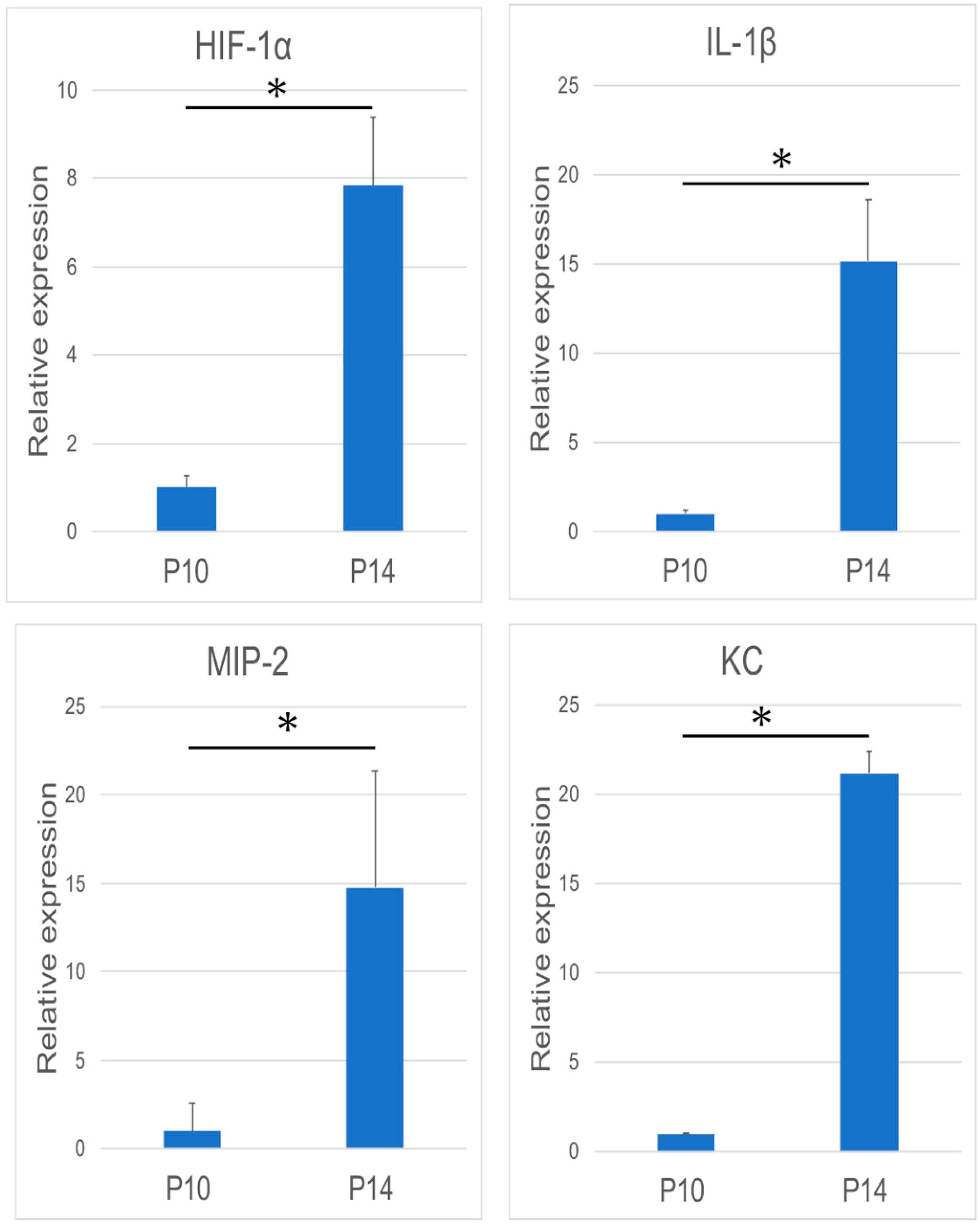
3.2. Chemokine and Cytokine Expression during Tooth Eruption
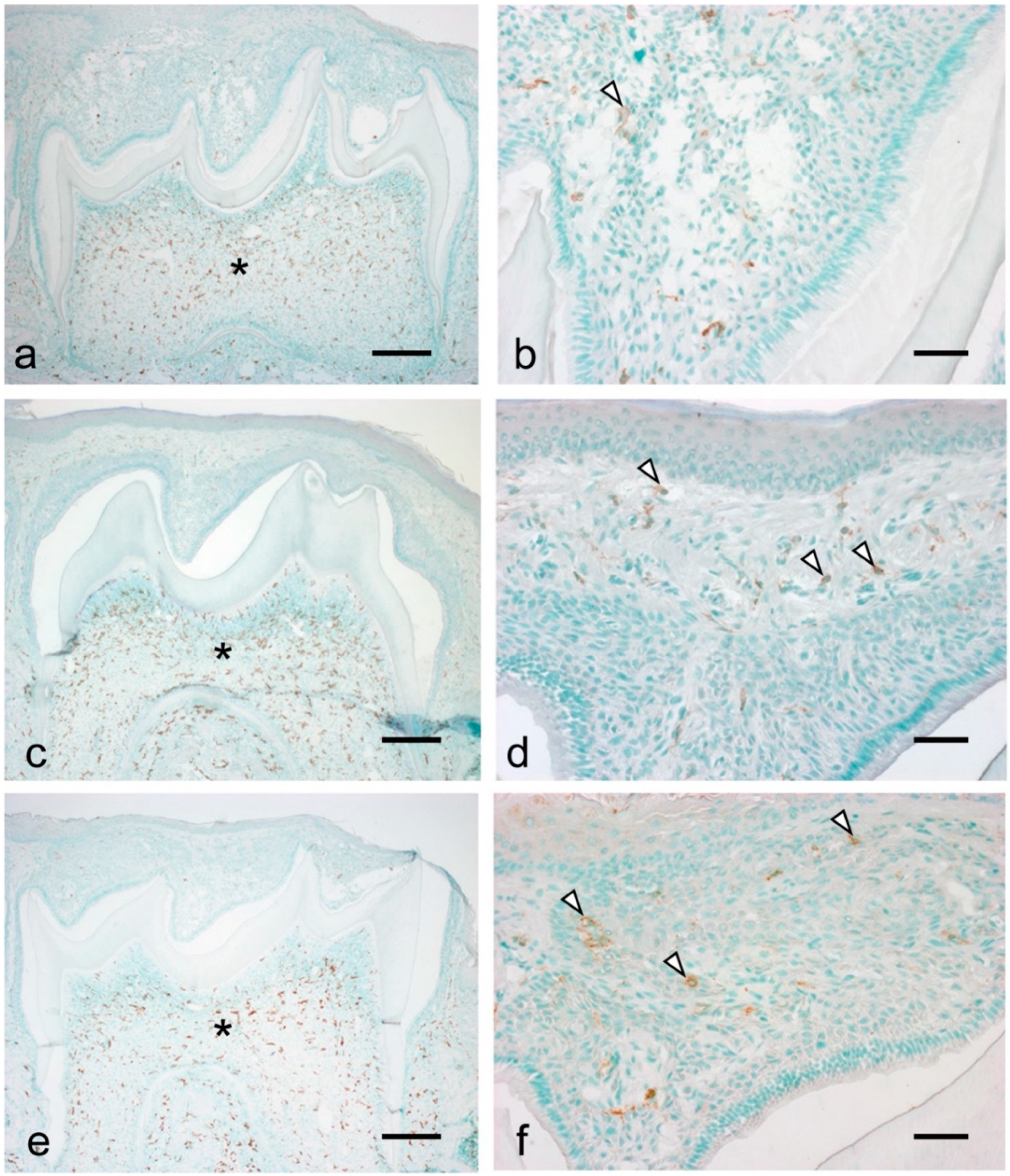
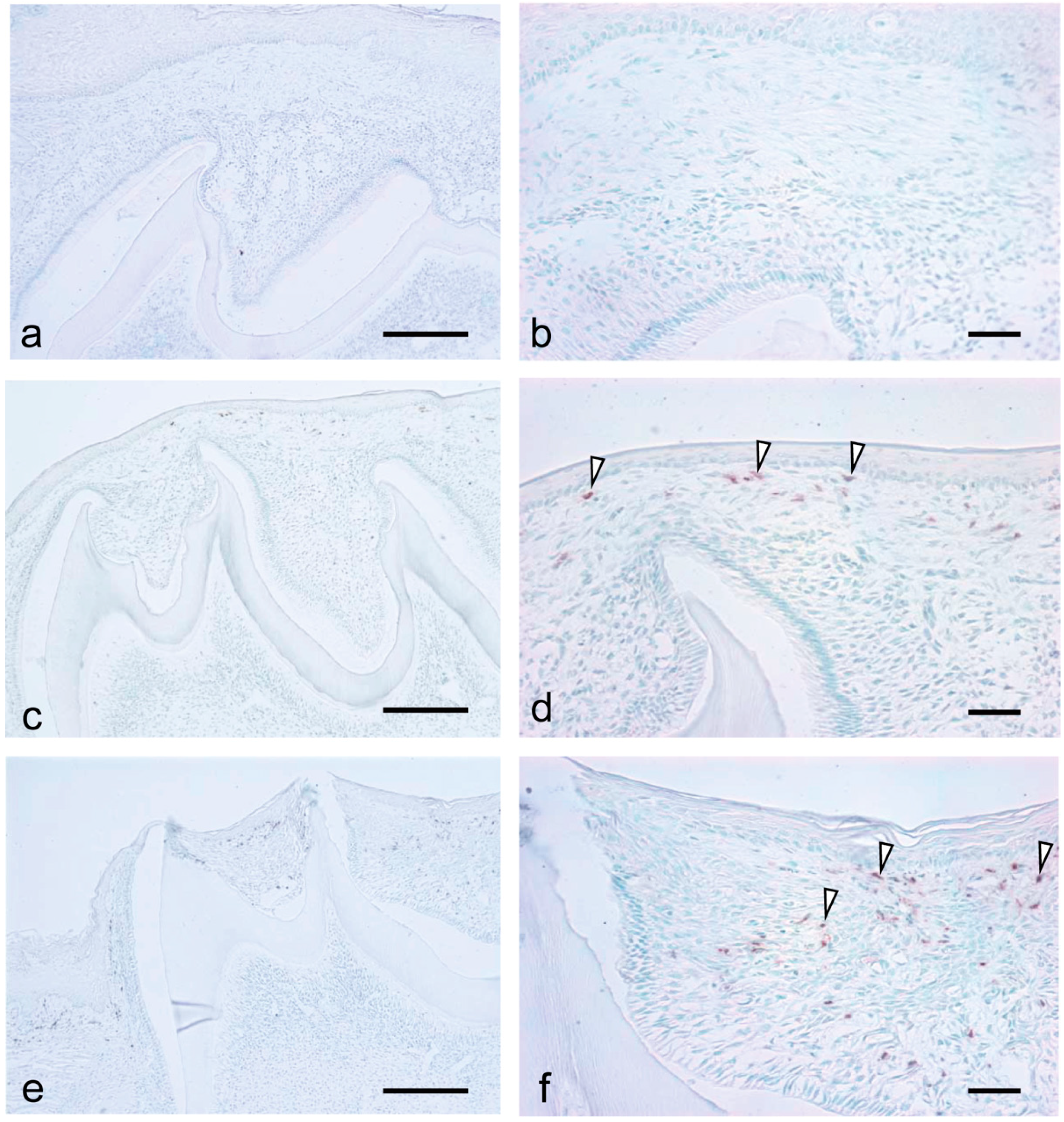
3.3. Migration of Macrophages and Neutrophils during Tooth Eruption
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Akalp, U.; Bryant, S.J.; Vernerey, F.J. Tuning tissue growth with scaffold degradation in enzyme-sensitive hydrogels: A mathematical model. Soft Matter 2016, 12, 7505–7520. [Google Scholar] [CrossRef]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular Matrix Degradation and Remodeling in Development and Disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a005058. [Google Scholar] [CrossRef]
- Mammoto, T.; E Ingber, D. Mechanical control of tissue and organ development. Development 2010, 137, 1407–1420. [Google Scholar] [CrossRef]
- Wise, G.E.; Yao, S.; Odgren, P.; Pan, F. CSF-1 Regulation of Osteoclastogenesis for Tooth Eruption. J. Dent. Res. 2005, 84, 837–841. [Google Scholar] [CrossRef]
- Wise, G.E.; King, G.J. Mechanisms of Tooth Eruption and Orthodontic Tooth Movement. J. Dent. Res. 2008, 87, 414–434. [Google Scholar] [CrossRef]
- Wise, G.E. Cellular and molecular basis of tooth eruption. Orthod. Craniofac. Res. 2009, 12, 67–73. [Google Scholar] [CrossRef]
- Marks, S.C.; Cahill, D.R.; Wise, G.E. The cytology of the dental follicle and adjacent alveolar bone during tooth eruption in the dog. Am. J. Anat. 1983, 168, 277–289. [Google Scholar] [CrossRef]
- Wise, G.E.; Marks, S.C.; Cahill, D.R. Ultrastructural features of the dental follicle associated with formation of the tooth eruption pathway in the dog. J. Oral Pathol. Med. 1985, 14, 15–26. [Google Scholar] [CrossRef]
- Wise, G.; Fan, W. Changes in the Tartrate-resistant Acid Phosphatase Cell Population in Dental Follicles and Bony Crypts of Rat Molars during Tooth Eruption. J. Dent. Res. 1989, 68, 150–156. [Google Scholar] [CrossRef]
- Yoo, H.I.; Kang, J.H.; Yang, S.Y.; Yong, J.H.; Moon, J.S.; Kim, M.S.; Jung, J.Y.; Koh, J.T.; Kim, W.J.; Oh, W.M.; et al. Differential expression of cxcl-14 during eruptive movement of rat molar germs. J. Exp. Zool. Part B Mol. Dev. Evol. 2011, 316, 418–426. [Google Scholar] [CrossRef]
- Cavalli, G.; Koenders, M.; Kalabokis, V.; Kim, J.; Tan, A.C.; Garlanda, C.; Mantovani, A.; Dagna, L.; Joosten, L.A.B.; Dinarello, C.A. Treating experimental arthritis with the innate immune inhibitor interleukin-37 reduces joint and systemic inflammation. Rheumatology 2016, 55, 2220–2229. [Google Scholar] [CrossRef] [PubMed]
- Opitz, B.; Van Laak, V.; Eitel, J.; Suttorp, N. Innate Immune Recognition in Infectious and Noninfectious Diseases of the Lung. Am. J. Respir. Crit. Care Med. 2010, 181, 1294–1309. [Google Scholar] [CrossRef] [PubMed]
- Rovina, N.; Panagiotou, M.; Pontikis, K.; Kyriakopoulou, M.; Koulouris, N.G.; Koutsoukou, A. Immune response to myco-bacterial infection: Lessons from flow cytometry. Clin. Dev. Immunol. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Tsuzurahara, F.; Nakamura, M. Macrophages are key cells for the initiation of Meckel’s cartilage disappearance. J. Oral Biosci. 2010, 52, 150–154. [Google Scholar]
- Tsuzurahara, F.; Soeta, S.; Kawawa, T.; Baba, K.; Nakamura, M. The role of macrophages in the disappearance of Meckel’s cartilage during mandibular development in mice. Acta Histochem. 2011, 113, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, Y.; Usui, M.; Yamamoto, G.; Takagi, Y.; Tachikawa, T.; Nakamura, M. Role of the junctional epithelium in periodontal innate defense and homeostasis. J. Periodontal Res. 2012, 47, 750–757. [Google Scholar] [CrossRef]
- Nakamura, M. Histological and immunological characteristics of the junctional epithelium. Jpn. Dent. Sci. Rev. 2018, 54, 59–65. [Google Scholar] [CrossRef]
- Kitamura, Y.; Fujikawa, K.; Fukushima, M.; Funae, T.; Makino, Y.; Ohki, R.; Nakamura, M. Immunohistochemical analysis of the origin of junctional epithelium in mice. J. Oral Health Dent. 2020, 3, 211–216. [Google Scholar]
- Rothlein, R.; Czajkowski, M.; O’Neill, M.M.; Marlin, S.D.; Mainolfi, E.; Merluzzi, V.J. Induction of intercellular adhesion molecule 1 on primary and continuous cell lines by pro-inflammatory cytokines. Regulation by pharmacologic agents and neutralizing antibodies. J. Immunol. 1988, 141, 1665–1669. [Google Scholar]
- Kuppner, M.C.; Van Meir, E.G.; Hamou, M.F.; De Tribolet, N. Cytokine regulation of intercellular adhesion molecule-1 (ICAM-1) expression on human glioblastoma cells. Clin. Exp. Immunol. 2008, 81, 142–148. [Google Scholar] [CrossRef]
- De Pizzol Júnior, J.P.; Sasso-Cerri, E.; Cerri, P.S. Apoptosis and reduced microvascular density of the lamina propria during tooth eruption in rats. J. Anat. 2015, 227, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Imtiyaz, H.Z.; Simon, M.C. Hypoxia-Inducible Factors as Essential Regulators of Inflammation. Curr. Top. Microbiol. Immunol. 2010, 345, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF Transcription Factors, Inflammation, and Immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, M.; Sasa, R.; Inoue, M.; Nakamura, M. Immunological Characterization of Junctional Epithelium: An Immunohistochemical Study. Dent. Med. Res. 2009, 29, 253–258. [Google Scholar] [CrossRef][Green Version]
- Yamada, Y.; Tashiro, E.; Taketani, S.; Imoto, M.; Kataoka, T. Mycotrienin II, a translation inhibitor that prevents ICAM-1 expression induced by pro-inflammatory cytokines. J. Antibiot. 2011, 64, 361–366. [Google Scholar] [CrossRef]
- Yang, C.-M.; Luo, S.-F.; Hsieh, H.-L.; Chi, P.-L.; Lin, C.-C.; Wu, C.-C.; Hsiao, L.-D. Interleukin-1β induces ICAM-1 expression enhancing leukocyte adhesion in human rheumatoid arthritis synovial fibroblasts: Involvement of ERK, JNK, AP-1, and NF-κB. J. Cell. Physiol. 2010, 224, 516–526. [Google Scholar] [CrossRef]
- Chen, C.; Xie, N.; Ling, J.; Du, Y.; Gu, H. Proteomic analysis of the effects of CSF-1 and IL-1? on dental follicle cells. Mol. Med. Rep. 2016, 14, 2405–2414. [Google Scholar] [CrossRef]
- Cabrera, S.M.; Henschel, A.M.; Hessner, M.J. Innate inflammation in type 1 diabetes. Transl. Res. 2016, 167, 214–227. [Google Scholar] [CrossRef]
- Perry, V. Innate Inflammation in Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a009373. [Google Scholar] [CrossRef]
- De França Landim, E.V.; de Carvalho, M.G.F.; de Carvalho, R.A.; Granville-Garcia, A.F.; de Freitas Lima, R.; de Brito Costa, E.M.M. Assessment of Gingival Mucosa of Infant Rats during Teething. J. Contemp. Dent. Pract. 2013, 14, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Cerri, P.S.; Pereira-Júnior, J.A.; Biselli, N.B.; Sasso-Cerri, E. Mast cells and MMP-9 in the lamina propria during eruption of rat molars: Quantitative and immunohistochemical evaluation. J. Anat. 2010, 217, 116–125. [Google Scholar] [CrossRef] [PubMed]
- De Pizzol Júnior, J.; Sasso-Cerri, E.; Cerri, P. Matrix Metalloproteinase-1 and Acid Phosphatase in the Degradation of the Lamina Propria of Eruptive Pathway of Rat Molars. Cells 2018, 7, 206. [Google Scholar] [CrossRef] [PubMed]
- Nissinen, L.; Kähäri, V. Matrix metalloproteinases in inflammation. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2014, 1840, 2571–2580. [Google Scholar] [CrossRef]
- Franco, C.; Patricia, H.R.; Timo, S.; Claudia, B.; Marcela, H. Matrix metalloproteinases as regulators of periodontal inflamma-tion. Int. J. Mol. Sci. 2017, 18, 440. [Google Scholar] [CrossRef] [PubMed]
- Pari, A.; Ilango, P.; Subbareddy, V.; Katamreddy, V.; Parthasarthy, H. Gingival Diseases in Childhood—A Review. J. Clin. Diagn. Res. 2014, 8, ZE01. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makino, Y.; Fujikawa, K.; Matsuki-Fukushima, M.; Inoue, S.; Nakamura, M. Role of Innate Inflammation in the Regulation of Tissue Remodeling during Tooth Eruption. Dent. J. 2021, 9, 7. https://doi.org/10.3390/dj9010007
Makino Y, Fujikawa K, Matsuki-Fukushima M, Inoue S, Nakamura M. Role of Innate Inflammation in the Regulation of Tissue Remodeling during Tooth Eruption. Dentistry Journal. 2021; 9(1):7. https://doi.org/10.3390/dj9010007
Chicago/Turabian StyleMakino, Yusuke, Kaoru Fujikawa, Miwako Matsuki-Fukushima, Satoshi Inoue, and Masanori Nakamura. 2021. "Role of Innate Inflammation in the Regulation of Tissue Remodeling during Tooth Eruption" Dentistry Journal 9, no. 1: 7. https://doi.org/10.3390/dj9010007
APA StyleMakino, Y., Fujikawa, K., Matsuki-Fukushima, M., Inoue, S., & Nakamura, M. (2021). Role of Innate Inflammation in the Regulation of Tissue Remodeling during Tooth Eruption. Dentistry Journal, 9(1), 7. https://doi.org/10.3390/dj9010007