Abstract
Background: Gram-positive bacteria, once considered incapable of producing extracellular vesicles (EVs) due to their thick peptidoglycan layer, are now known to secrete EVs that transport virulence factors and modulate host immunity. These EVs contribute to bacterial pathogenicity by facilitating biofilm formation, immune evasion, and inflammation. Granulicatella adiacens, an oral commensal associated with infective endocarditis, represents a clinically relevant model to study EV-mediated virulence. Objectives: This study’s aim was to investigate whether the proteomic composition and immunomodulatory activity of G. adiacens EVs differ between biofilm and planktonic lifestyles, thereby contributing to distinct pathogenic behaviours. Methods: EVs isolated from G. adiacens CCUG 27809 cultures were characterized using nano LC-ESI-MS/MS, followed by comprehensive bioinformatic and cytokine assays. Results: Quantitative proteomic profiling identified 1017 proteins, revealing distinct signatures between biofilm- and planktonic-derived EVs. Principal component analysis showed clear segregation between the two states, with biofilm EVs enriched in proteins linked to stress adaptation, adhesion, and structural integrity, while planktonic EVs exhibited growth- and metabolism-related proteins. A total of 114 virulence-associated proteins were identified, including several novel candidates. Functionally, EVs from both conditions significantly induced pro-inflammatory cytokines IL-8 and IL-1β in a dose-dependent manner (p < 0.05), whereas IL-17 remained unchanged. Conclusions: G. adiacens EVs exhibit lifestyle-dependent proteomic and immunomodulatory differences, underscoring their role in host–pathogen interactions and endocardial infection. These findings provide a foundation for future mechanistic and in vivo studies exploring EV-mediated virulence and potential therapeutic modulation.
1. Introduction
Transient bacteremia following dental procedures can allow oral microorganisms to enter the bloodstream, occasionally leading to infective endocarditis [1,2]. Among these, Granulicatella species—facultatively anaerobic, Gram-positive cocci—have emerged as opportunistic pathogens associated with oral and systemic infections, notably infective endocarditis [3,4,5]. Although G. adiacens is part of the normal oral microbiota [6], it has been implicated in dental caries, endodontic infections, and periodontitis [7,8,9], suggesting that under specific conditions it can shift from commensalism to pathogenicity.
Bacterial extracellular vesicles (EVs) have emerged as crucial mediators of intercellular communication and virulence. Gram-negative bacteria are well known to produce outer membrane vesicles (OMVs) derived from membrane blebbing [10,11], but recent studies confirm that Gram-positive species also release EVs despite their thick peptidoglycan walls [12,13,14] and recent reviews have updated mechanisms of biogenesis, composition, and roles of Gram-positive EVs in pathogenesis and host modulation [15,16]. These Gram-positive EVs can carry a range of components—including enzymes, toxins, lipids, and nucleic acids—and have been implicated in modulating host immune responses, promoting biofilm formation, and enhancing bacterial survival under stress [14,17]. Furthermore, external interventions such as antimicrobial agents [18,19,20] and photobiomodulation (e.g., low-level laser therapy) [21], can alter oral microbiota composition and host responses, potentially influencing EV production and signalling, underscoring their clinical significance.
In line with proteomic differences between biofilm and planktonic cells [22,23], proteomic analyses comparing EVs from biofilm and planktonic conditions have highlighted quantitative differences in composition that likely reflect the physiological conditions [24]. In the context of oral cavity, previous studies have profiled the secretome and EV proteome of G. adiacens [25,26], yet the impact of bacterial lifestyle—biofilm versus planktonic growth—on EV composition and virulence potential remains unexplored. Biofilm formation is known to enhance microbial resistance and virulence [27,28,29]; therefore, EVs released under these conditions may differ functionally and structurally from those produced in planktonic cultures.
This study aims to test the hypothesis that EVs released by G. adiacens during biofilm growth differ proteomically and functionally from those produced in planktonic conditions, contributing to distinct mechanisms of virulence and host immune modulation. To address this, we isolated and characterized EVs from both lifestyles using high-resolution quantitative proteomics and evaluated their cytokine-inducing capacity in vitro. This comparative approach provides the first insight into lifestyle-specific EV-mediated pathogenic pathways in G. adiacens.
2. Materials and Methods
2.1. Bacterial Strain and Culture Conditions
The reference strain Granulicatella adiacens CCUG 27809 was used for all experiments. This well-characterized strain was grown on chocolate blood agar (CBA) supplemented with 0.001% pyridoxal hydrochloride and incubated at 37 °C under 5% CO2 for 48 h. A loopful of colonies was subsequently inoculated into brucella broth containing 0.001% pyridoxal hydrochloride and incubated under identical conditions for another 48 h. Three independent biological replicates were prepared for all experiments.
2.2. Biofilm and Planktonic Cultures
Biofilm and planktonic cultures were prepared following a previously described method with minor modifications [30]. Bacterial cells from agar plates were suspended in brucella broth and washed once (5000× g, 5 min) to ensure purity. The cell pellet was resuspended in 1 mL of fresh broth, and the optical density (OD600) was adjusted to 1.
For biofilm formation, 100 µL of this standardized suspension was inoculated into each well of a 24-well plate containing 900 µL of brucella broth. Planktonic cultures were initiated by adding 100 µL of the same suspension to 900 µL of broth in sterile microfuge tubes. Wells and tubes containing only brucella broth served as negative controls. Cultures were incubated at 37 °C under 5% CO2 for 24 h.
2.3. Isolation of Extracellular Vesicles (EVs)
EVs were isolated using a differential centrifugation and filtration protocol [25] with modifications. Broth cultures were centrifuged at 5000× g for 10 min to remove cells (Eppendorf, Hamburg, Germany). The supernatant was filtered through a 0.22 µm syringe filter (Millipore, Darmstadt, Germany) to eliminate residual bacteria and ultracentrifuged at 125,000× g for 3 h at 4 °C (Beckman, Indianapolis, IN, USA). The resulting pellet was resuspended in sterile phosphate-buffered saline (PBS) and stored at −20 °C until use.
2.4. Scanning Electron Microscopy (SEM)
EV samples were fixed in PBS containing 3% glutaraldehyde for 2 h at room temperature, followed by overnight storage at 4 °C. Post-fixation was carried out with 1% osmium tetroxide for 2 h. Samples were dehydrated through a graded acetone series (30–100%), critical-point dried, mounted on aluminum stubs with carbon tape, sputter-coated with gold, and stored in a desiccator. Imaging was performed using a Zeiss Leo Supra 50 VP field emission SEM (Carl Zeiss, Oberkochen, Germany).
2.5. Protein Quantification and SDS-PAGE
Protein concentrations were determined using the Quick Start™ Bradford Microplate Assay (Bio-Rad, Hercules, CA, USA). Equal protein quantities were denatured in 2× Laemmli buffer (100 °C, 5 min), centrifuged (5000× g, 5 min), and separated by SDS-PAGE (12% gels, 120 V). Silver staining was used for visualization, and images were captured using a ChemiDoc™ MP Imaging System (Bio-Rad, USA).
2.6. Sample Preparation for Mass Spectrometry
A total of 25 µg of each EV protein sample was reduced with 5 mM TCEP, alkylated with 50 mM iodoacetamide, and digested overnight with trypsin (1:50 enzyme–substrate, 37 °C). Peptides were purified using C18 silica cartridges, dried under vacuum, and resuspended in buffer A (2% acetonitrile, 0.1% formic acid).
2.7. LC-MS/MS Analysis
Peptides were analyzed on an Easy-nLC 1000 system coupled to an Orbitrap Exploris 240 mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA). Peptides (1 µg) were loaded onto a Picofrit column (1.8 µm resin, 15 cm) and eluted with a 0–38% gradient of buffer B (80% acetonitrile, 0.1% formic acid) at 500 nL/min for 96 min. The instrument was operated in data-dependent acquisition mode (top 20) with a dynamic exclusion of 30 s. MS spectra were acquired at 60 K resolution (m/z 375–1500), and MS2 spectra at 15 K resolution.
Raw data have been deposited in the PRIDE repository (Project ID: PXD059541; username: reviewer_pxd059541@ebi.ac.uk; password: gyAZOgM9BoHt).
2.8. Data Processing
Raw files were analyzed using Proteome Discoverer v2.5 against the G. adiacens UniProt database. Search parameters included a precursor mass tolerance of 10 ppm and fragment tolerance of 0.02 Da. Trypsin/P was specified as the digestion enzyme, with carbamidomethyl (C) as a fixed modification and oxidation (M) and N-terminal acetylation as variable modifications. Peptide-spectrum matches and protein-level false discovery rates (FDR) were controlled at 1%.
2.9. Data Pre-Processing and Quality Control
Proteins were retained only if quantified in ≥2 of 3 biological replicates under at least one condition (biofilm or planktonic). Abundance values were log2-transformed and missing data imputed using the “MinProb” method, assuming left-censored missingness. This approach models low-abundance proteins while minimizing variance inflation.
2.10. Differential Protein Expression Analysis
Differentially expressed proteins between biofilm and planktonic EVs were identified using the limma package (R/Bioconductor), which applies empirical Bayes moderation to improve statistical power. Adjusted p-values (Benjamini–Hochberg correction) < 0.05 and |log2 fold change| > 1.5 were considered significant.
2.11. Data Visualization and Functional Analysis
Principal component analysis (PCA), volcano plots, and hierarchical clustering were performed using R (v4.2) packages FactoMineR, EnhancedVolcano, and ComplexHeatmap, respectively. Gene Ontology (GO) enrichment was carried out in ExpressAnalyst using hypergeometric testing with FDR correction (adjusted p < 0.05). Protein–protein interaction networks were generated using STRING v12.5 (confidence > 0.7) to identify functional modules and hub proteins.
2.12. Cytokine Quantification
Peripheral blood mononuclear cells (PBMCs) were stimulated with EVs, and cytokines (IL-8, IL-1β, IL-17) were quantified using Quantikine® ELISA kits (R&D Systems, Minneapolis, MN, USA) following the manufacturer’s instructions. Absorbance was measured at 450 nm with wavelength correction at 570 nm (iMark™ Microplate Reader, Bio-Rad). All assays were performed in duplicate, and cytokine concentrations were calculated from standard curves.
2.13. Statistical Analysis
For proteomics data, differential abundance testing was performed as described above. Cytokine data, which were non-normally distributed, were compared using the nonparametric Mann–Whitney U test (p < 0.05). Statistical analyses were performed using SPSS v25.0.
2.14. Ethical Approval
This study was approved by the ethical committee of the Health Sciences Center, Kuwait University (DR/EC/3413, Date: 31 October 2018), and has been carried out in full accordance with the World Medical Association Declaration of Helsinki. The blood donor received written information about the nature and purposes of the study and a written informed consent was obtained upon his/her approval to participate.
3. Results
3.1. EV Preparation from G. adiacens Biofilm and Planktonic Cells
Following previously established protocols in our laboratory, vesicles were isolated from bacterial cultures of the reference strain G. adiacens CCUG 27809. Determination of the protein concentration in EV preparations revealed values ranging from 400 to 600 µg across three independent isolations. SDS–PAGE analysis (Figure 1A) showed distinct banding patterns between vesicles derived from biofilm and planktonic growth. In silico analysis of the protein sequences using a 2D-gel analysis tool indicated that most cytoplasmic proteins had isoelectric points (pI) between 4.5 and 6.0, whereas most secreted proteins were in the pI range of 8.0 to 9.6 (Figure 1B). Scanning electron microscopy (SEM) of EV preparations showed no observable morphological differences between the two sample types in terms of size or abundance (Figure 1C).
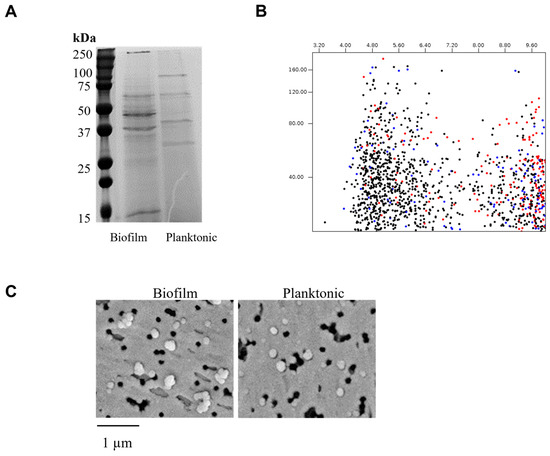
Figure 1.
Analysis of G. adiacens extracellular vesicles. Vesicle preparations containing standard amounts of protein were run on an SDS-PAGE gel electrophoresis (Panel (A)) and in silico 2D gel visualization (Panel (B)). Scanning electron microscopy was performed for the vesicle preparations (Panel (C)).
3.2. Differential Protein Expression in Vesicles Produced Under Biofilm and Planktonic Conditions
To define molecular adaptations of G. adiacens to different growth modes, quantitative proteomic profiling was performed on extracellular vesicles (EVs) isolated from biofilm and planktonic cultures. The volcano plot (Figure 2A) highlighted proteins significantly upregulated in biofilm (blue) and planktonic (red) conditions, while grey points represented non-significant changes. Principal component analysis (PCA) revealed strong separation between the two states, with the first component explaining ~75% of total variance. Pearson’s correlation heatmaps (Figure 2B) demonstrated large co-regulated protein modules, indicating that the biofilm–planktonic transition involves coordinated regulation of functional networks rather than random protein-level changes.
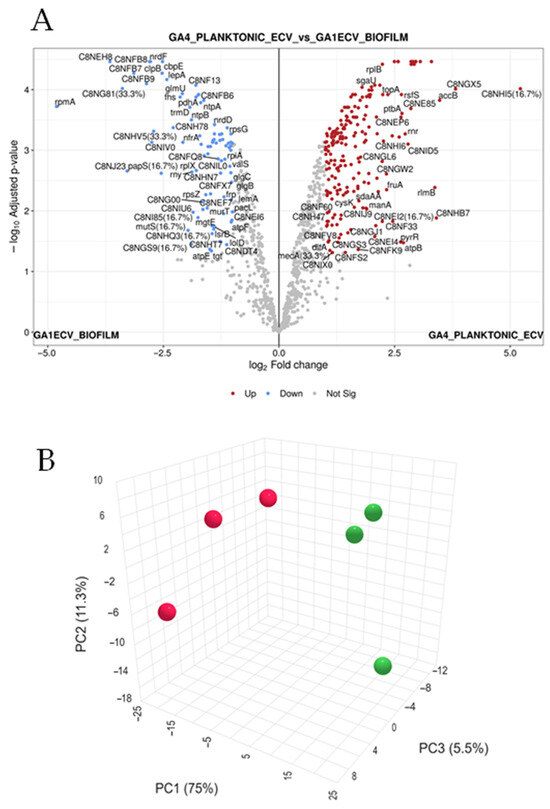
Figure 2.
Global proteomic landscape of G. adiacens EVs from biofilm and planktonic cultures. This figure presents a global differential proteomic analysis, highlighting the distinct molecular signatures of extracellular vesicles (EVs) from G. adiacens grown in biofilm versus planktonic states. (A) Volcano plot illustrating the magnitude of protein expression change (x-axis: log2-FC) versus statistical significance (y-axis: −log10 adjusted p-value). Proteins significantly upregulated in planktonic EVs are shown as red points, those upregulated in biofilm EVs are blue points, and non-significantly changed proteins are grey points. Significance was determined by an adjusted p-value < 0.05 and an absolute log2-FC > 1.5. Key differentially expressed proteins are labelled. (B) Three-dimensional Principal Component Analysis (PCA) plot visualizing the global proteomic variance between samples. The plot demonstrates clear spatial separation of the biofilm (red spheres) and planktonic (green spheres) sample clusters. The first three principal components (PC1, PC2, and PC3) account for 75%, 11.3%, and 5.5% of the total variance, respectively, indicating distinct proteomic profiles. Statistical analysis was performed using a linear model with Benjamini–Hochberg correction for multiple testing. Red dots indicate biofilms while the green ones indicate planktonic samples.
Mass spectrometry identified 1017 proteins in biofilm-derived EVs and 1048 in planktonic EVs. Consistency across biological replicates was confirmed by line plots showing log2-transformed intensity profiles for significantly upregulated and downregulated (Figure S1A) proteins, both displaying uniform expression trends. Proteomic analysis revealed distinct functional profiles between biofilm and planktonic vesicles. Biofilm-associated EVs were enriched in proteins related to cell wall and peptidoglycan synthesis (e.g., Lipid II isoglutaminyl synthase subunits GatD and MurT, peptidoglycan glycosyltransferase, LPXTG-motif cell wall anchor proteins), ATP synthase components, and energy metabolism enzymes. These trends suggest an active structural and metabolic adaptation supporting matrix formation and sessile survival. In contrast, planktonic vesicles showed upregulation of proteins involved in translation, redox metabolism, and cell division (e.g., ribosomal proteins, FtsZ, FtsA, redoxin family enzymes), reflecting rapid growth and motility.
Gene Set Enrichment Analysis (GSEA) further supported these distinctions (Figure S1B). Pathways related to cell wall metabolism (Enrichment Score = 0.106) and stress response (Enrichment Score = 0.083) were enriched in biofilm EVs, whereas transport systems (Enrichment Score = –0.373) and transcription machinery (Enrichment Score = –0.517) were enriched in planktonic EVs.
3.3. Differentially Expressed Proteins Exhibit Consistent and Robust Expression Patterns
The robustness of the proteomic signatures was confirmed through several quality-control analyses. Unsupervised hierarchical clustering of Z-score normalized abundances of significantly differentially expressed proteins segregated samples strictly according to their growth condition (Figure 3). All biofilm replicates clustered together, distinct from planktonic replicates, demonstrating systematic and reproducible proteomic divergence.
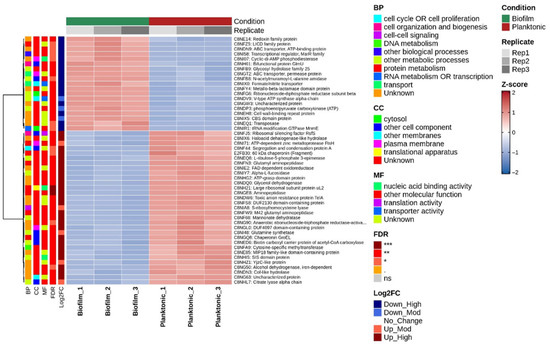
Figure 3.
Hierarchical clustering of differentially expressed EV proteins. This figure provides a detailed examination of the expression patterns of significantly altered proteins, confirming the robustness and reproducibility of the proteomic data. A, Heatmap displaying the Z-score normalized, log2-transformed abundance of significantly differentially expressed proteins. Rows represent individual proteins and columns represent biological replicates (n = 3 per condition). The color key indicates relative abundance (blue: low abundance; red: high abundance). Unsupervised hierarchical clustering (Ward.D2 method, Euclidean distance) was applied to both samples (columns) and proteins (rows). The column dendrogram demonstrates a clear distinction between biofilm and planktonic samples, signifying their disparate EV proteomes. Annotations for Gene Ontology (GO) biological process, false discovery rate (FDR), and log2FC are provided. *: FDR < 0.05, **: FDR < 0.01, ***: FDR < 0.001, n.s.: not significant.
Row-wise clustering revealed two major co-regulated protein blocks: one consistently upregulated in biofilms (red) and the other downregulated (blue). Sample-to-sample correlation analysis (Figure S2A) showed strong within-group correlations (r ≈ 1.0) and low between-group correlations, confirming excellent reproducibility. Violin plots of log2-transformed protein abundances showed consistent distributions across all samples, ruling out normalization bias (Figure S2B).
Representative scatter plots (Figure S3) for select proteins showed statistically significant differences (p < 0.01) between biofilm (blue) and planktonic (orange) replicates, validating the robustness of the differential expression patterns.
3.4. Protein Overlaps and Distribution Between Vesicles from Biofilm and Planktonic Conditions
Protein overlap analysis (Figure 4) highlighted substantial similarity between the two proteomes: 96% (979 proteins) were shared between biofilm and planktonic EVs, while 22 and 17 proteins were unique to biofilm and planktonic conditions, respectively (Figure 4A). This suggests that transitions between growth states primarily reflect quantitative modulation of a shared proteome rather than qualitative replacement.
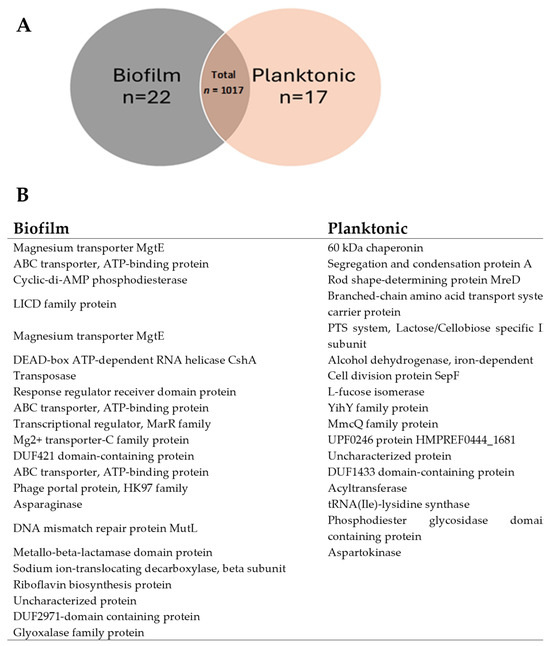
Figure 4.
Protein Overlap and Distribution Between BIOFILM and PLANKTONIC States in G. adiacens. This figure dissects the composition of the biofilm and planktonic EV proteomes, highlighting both shared and unique protein components. (A) Venn diagram illustrating the overlap in protein expression between biofilm (GA1ECV_biofilm, red) and planktonic (GA4_planktonic_ECV, blue) conditions. Of the total 1017 proteins, 979 (96%) are shared between the two conditions, while 22 (2.16%) are unique to biofilm and 17 (1.67%) to planktonic. The count of proteins in each category is color-coded to indicate protein abundance, with higher counts shown in darker blue. (B) Table listing a selection of functionally relevant proteins unique to either the biofilm or planktonic state. Biofilm-unique proteins include factors associated with environmental adaptation and stress, such as the magnesium transporter MgtE and the DNA mismatch repair protein MutL. Planktonic-unique proteins include factors related to active proliferation, such as the cell division protein SepF and a 60 kDa chaperonin. These unique proteins may function as specialized molecular determinants for each distinct mode of existence. The overlap between biofilm and planktonic states was determined using a Venn diagram, and the protein counts were visualized in a bar plot to show the number of shared and unique proteins between conditions.
The biofilm-unique proteins included magnesium transporters (MgtE), ABC transporters, cyclic-di-AMP phosphodiesterase, and metallo-β-lactamase—proteins likely contributing to biofilm stability and stress resilience. Planktonic-specific proteins, such as the 60-kDa chaperonin, alcohol dehydrogenase, and SepF, were associated with growth and division. These findings identify biofilm-specific molecular targets (e.g., MgtE, ABC transporters) that may inform anti-biofilm strategies.
3.5. Gene Ontology and Functional Annotation of Differentially Expressed Proteins
Network analysis (Figure 5A) revealed a central “Metabolic Process” hub linking major branches such as proteolysis, carbohydrate derivative metabolism, and organic acid metabolism—indicating coordinated regulation of energy and matrix-related processes. Biofilm vesicles emphasized protein degradation and cell envelope processes, while planktonic vesicles favoured biosynthesis and cellular development.
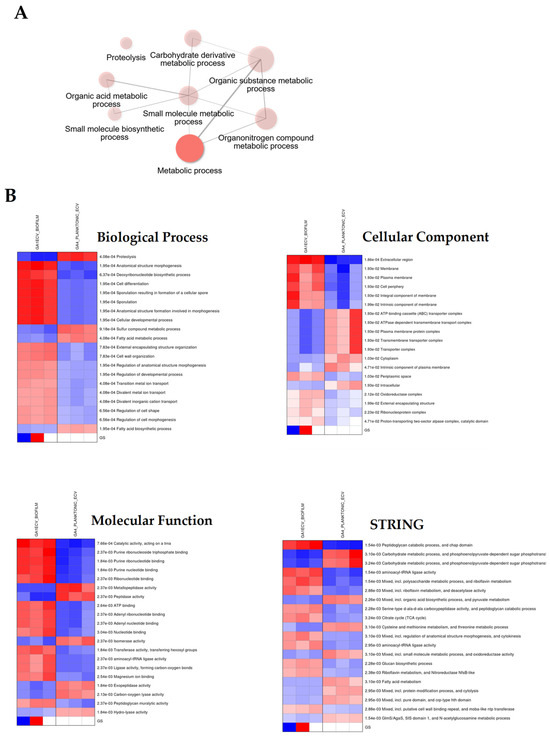
Figure 5.
Gene Ontology and Functional Annotation of Differentially Expressed Proteins in G. adiacens. Panel (A): Network diagram illustrating the enrichment of gene ontology (GO) biological processes (BP) related to metabolic process. Proteins that are differentially expressed between biofilm (GA1ECV_biofilm, red) and planktonic (GA4_planktonic_ECV, blue) conditions are linked to specific biological processes. The sizes of the nodes correspond to the number of proteins associated with each process, with the largest node representing metabolic process. other enriched terms include proteolysis, small molecule metabolic process, and carbohydrate derivative metabolic process. Panel (B): Heatmaps of the top enriched GO terms for biofilm and planktonic conditions across three categories: The color intensity corresponds to the level of enrichment, with darker red and blue indicating more significant terms. Gene Ontology enrichment analysis was performed to identify biological processes, cellular components, and molecular functions enriched in differentially expressed proteins between the biofilm and planktonic conditions. A p-value threshold of 0.05 was used for statistical significance, and the results were visualized using network diagrams and heatmaps. Proteins were clustered based on their GO term associations, and hierarchical clustering was applied to group related terms.
Heatmaps (Figure 5B) showed distinct enrichment patterns: biofilms exhibited elevated activity in proteolysis and membrane-associated processes, whereas planktonic cells demonstrated higher activity in metabolic and growth-related functions.
3.6. Proteins with Predicted Virulence Potential
Table 1 lists 114 proteins with predicted virulence potential, several of which are well-documented in bacterial pathogenicity. Of the 22 biofilm-unique proteins, six were predicted to be virulent, compared to none in the planktonic-unique set. These included Type VII secretion system components (e.g., YukD, EsaA), LPXTG-motif anchor proteins, metallo-β-lactamase, and cell wall-binding repeat proteins. High-scoring ABC transporters and several uncharacterized proteins were also identified, suggesting potential new virulence factors warranting further investigation. Overall, biofilm EVs were enriched in surface-associated and secretion-related virulence determinants.

Table 1.
Predicted virulent proteins.
3.7. Proinflammatory Potential of EV Preparations
The proinflammatory effects of the EV preparations were evaluated using quantitative ELISA kits (R&D Systems). As shown in Figure 6, significant differences (p < 0.05) were observed in IL-8 and IL-1β levels between biofilm- and planktonic-derived EVs, with a dose-dependent increase in both cytokines. IL-17 levels did not differ significantly between the two conditions.
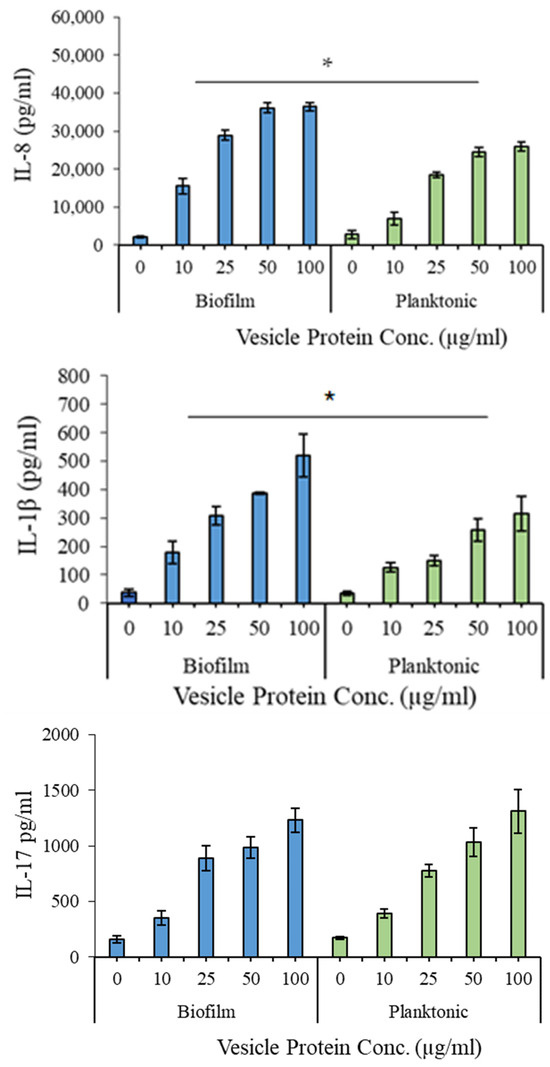
Figure 6.
Proinflammatory potential of the extracellular vesicles produced by G. adiacens in biofilm and planktonic growth. Vesicle preparations at the concentrations 10, 25, 50, and 100 µg/mL were used for stimulating human PBMCs. Cytokine induction was considered significantly different at * p < 0.05.
4. Discussion
The present study provides the first comprehensive characterization of extracellular vesicles (EVs) produced by G. adiacens under biofilm and planktonic growth conditions, revealing distinct proteomic and immunostimulatory profiles associated with each lifestyle. These findings support the growing evidence that bacterial EVs are key mediators of virulence, intercellular communication, and environmental adaptation [11,12,14] and recent studies further highlight their diverse immunomodulatory roles and clinical potential in Gram-positive infections [31,32]. While earlier work examined protein secretion in Granulicatella species [25], this is the first report describing proteomic differences in EVs derived from biofilm versus planktonic G. adiacens, offering novel insights into its pathogenic potential.
SDS-PAGE analysis revealed subtle but reproducible differences in EV protein profiles between biofilm and planktonic cultures, suggesting condition-specific variation in protein cargo. In silico 2D-gel mapping indicated that cytoplasmic proteins clustered predominantly in the acidic pI range (4.5–6.0), whereas secreted proteins were enriched in the alkaline range (8.0–9.6), consistent with trends observed in other Gram-positive bacteria. SEM imaging showed no morphological differences between biofilm- and planktonic-derived vesicles, implying that their functional divergence arises primarily from molecular composition rather than structural variation.
Proteomic analysis identified 1017 proteins, with 96% shared between both conditions. Nevertheless, differential abundance patterns and a subset of condition-specific proteins (22 biofilm-unique, 17 planktonic-unique) suggest specialized adaptations. This near-complete overlap implies that the shift between these lifestyles relies primarily on quantitative changes (up- or down-regulation) in a shared proteome rather than a largescale protein replacement. However, unique proteins in each state, though small in number—could play pivotal roles in biofilm formation or planktonic lifestyle, acting as specialized molecular switches for these distinct modes of existence. Biofilm-specific proteins such as magnesium transporter (mgtE) and Cyclic-di-AMP phosphodiesterase have been shown to play important roles in the regulation of type III secretion system in different bacteria [33,34]. Further, deletion of mutL, a protein involved in DNA mutation repair was found to affect adhesion and biofilm formation leading to an overall reduced virulence [35]. Proteins with metallo-beta lactamase domain can confer resistance to a broad range of beta-lactam antibiotics. Its presence in biofilm vesicles indicates a mechanism for antibiotic inactivation [36].
Biofilm EVs were enriched in proteins linked to cell-wall biogenesis and metabolic persistence, such as GatD, MurT, and LPXTG-anchored surface proteins. The prominence of MurT/GatD pathway components—known to be essential for cell-wall amidation and antibiotic tolerance in other pathogens including S. aureus and M. tuberculosis [37,38,39]—suggests a similar role in maintaining biofilm integrity and survival. Elevated levels of stress-response chaperones (DnaK, trigger factor) and ribosome hibernation factors further align with the biofilm’s demand for long-term stability under nutrient-limited conditions.
Conversely, planktonic EVs showed enrichment in ribosomal proteins, FtsZ, and transcriptional regulators (SigA, Rho), consistent with rapid proliferation and metabolic flexibility. The presence of redox-active enzymes such as glycerol dehydrogenase implies enhanced energy metabolism to sustain a free-living state. PCA and hierarchical clustering analyses further confirmed clear proteomic segregation between biofilm and planktonic EVs, reflecting extensive physiological reprogramming during the biofilm–planktonic transition.
Gene Ontology (GO) enrichment analyses revealed that biofilm EVs are functionally biased toward proteolysis and carbohydrate metabolism, likely supporting matrix remodelling and energy production, whereas planktonic EVs favoured metabolic versatility and cell development pathways. Notably, 114 virulence-associated proteins—including Type VII secretion components (YukD, EsaA), cell-wall hydrolases, and antibiotic resistance determinants (metallo-β-lactamase)—were identified, highlighting the potential contribution of G. adiacens EVs to host–pathogen interactions and disease progression.
Functionally, EVs from both growth states triggered significant induction of IL-8 and IL-1β in human PBMCs, with biofilm-derived EVs eliciting a stronger proinflammatory response. This selective cytokine activation is consistent with previous observations that biofilm bacteria provoke heightened innate immune signalling [12,40]. Interestingly, IL-17 levels remained unchanged, suggesting a targeted immune modulation rather than broad activation. IL-17 is an important cytokine in the mucosal immunity. The observed similarity in IL-17 stimulation by biofilm and planktonic EVs is likely because they share similar outer envelope components, such as bacterial cell wall elements and surface proteins, which are known to induce this key mucosal immunity cytokine [41,42]. The observed cytokine patterns are relevant to G. adiacens pathogenesis, given the roles of IL-8 and IL-1β in infective endocarditis and oral infections [43,44]. While TNF-α and IL-6 were not assessed in this study, future cytokine panels may include these key inflammatory mediators to better define the immunomodulatory spectrum of G. adiacens EVs.
Extensive research on bacterial membrane vesicles has established that a diverse array of well-characterized toxins and non-toxin virulence factors are secreted via vesicle pathway [45,46,47]. In contrast to their soluble counterparts, vesicle-associated virulence factors are uniquely shielded from degradation by host proteases [11]. Furthermore, vesicles facilitate the targeted delivery of these factors as concentrated packages to host cells and tissues, thereby amplifying damage at specific sites. Similar to oral bacteria implicated in infective endocarditis—such as Aggregatibacter actinomycetemcomitans [48], Kingella kingae [49], and others [25,50,51]—it is plausible that G. adiacens similarly utilizes its extracellular vesicles, which are enriched with numerous putative virulence proteins, in the pathogenesis of this infection.
5. Limitations and Future Directions
This study was conducted using EVs derived from the G. adiacens reference strain CCUG 27809 to ensure experimental reproducibility. The other limitation is that functional control experiments using heat- or protease-treated EVs were not performed in this study and will be addressed in future work to confirm that cytokine induction is protein-mediated. Future investigations should validate these findings using multiple clinical isolates to assess strain-specific variability. Additionally, while the proteomic and immunostimulatory characterizations provide a strong foundation, further in-depth analyses—including mechanistic studies with modified or digested EVs and in vivo infection models—are warranted to establish causal relationships between EV composition and virulence. Further research exploring the molecular mechanisms through which EVs mediate host–pathogen communication could also elucidate novel therapeutic targets.
6. Conclusions
This work demonstrates that G. adiacens produces extracellular vesicles with distinct molecular and immunological characteristics depending on its growth mode. Biofilm-derived EVs exhibit enrichment in structural and stress-response proteins, whereas planktonic EVs reflect a metabolically active state. The presence of multiple virulence-associated proteins and proinflammatory activity underscores the potential contribution of EVs to G. adiacens pathogenicity. This is consistent with recent reviews that discuss EV-mediated modulation of antibiotic tolerance and the potential diagnostic/therapeutic implications of bacterial EVs in clinical infections [15,32]. By elucidating these vesicular adaptations, our study provides a foundation for future mechanistic and translational research aimed at understanding and mitigating infections caused by this understudied organism.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/dj13120557/s1, Figure S1. Global proteomic landscape of G. adiacens EVs from biofilm and planktonic cultures. Figure S2. Quality assessment of differentially expressed EV proteins. Figure S3. Expression levels of proteins.
Author Contributions
Conceptualization, M.K.; methodology, M.K.; software, M.K.; validation, M.K.; formal analysis, M.K.; investigation, M.K.; resources, M.K.; data curation, M.K.; writing—original draft preparation, M.K.; writing—review and editing, M.K. and S.A.; visualization, M.K. and S.A.; supervision, M.K.; project administration, M.K.; funding acquisition, M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Research Administration of Kuwait University (Grant # DB01/19).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the ethical committee of the Health Sciences Centre, Kuwait University (DR/EC/3413, Date: 31 October 2018), and has been carried out in full accordance with the World Medical Association Declaration of Helsinki.
Informed Consent Statement
Informed consent was obtained from the subject who participated in the study. The blood donor received written information about the nature and purposes of the study and a written informed consent was obtained upon his/her approval to participate.
Data Availability Statement
Raw mass spectrometry data have been deposited in the PRIDE repository (Project ID: PXD059541; username: reviewer_pxd059541@ebi.ac.uk; password: gyAZOgM9BoHt).
Acknowledgments
We wholeheartedly thank Hamad Yadikar at the department of Biological Sciences, College of Science, Kuwait University for kind help with proteomics data interpretation and manuscript review and comments. We also thank Krishna Girija and Febine Mathew for their hard work and excellent technical support for this project. We are grateful to Jessy Mathew at the Electron Microscopy Facility, Faculty of Medicine, Kuwait University. Our sincere thanks to the Research Administration of Kuwait University for their generous funding to this project. The study was conducted at the Oral Microbiology Research Laboratory, College of Dentistry, Kuwait University (SRUL 01/14).
Conflicts of Interest
No potential conflicts of interest were reported by the authors.
References
- Lockhart, P.B.; Brennan, M.T.; Thornhill, M.; Michalowicz, B.S.; Noll, J.; Bahrani-Mougeot, F.K.; Sasser, H.C. Poor oral hygiene as a risk factor for infective endocarditis-related bacteremia. J. Am. Dent. Assoc. 2009, 140, 1238–1244. [Google Scholar] [CrossRef]
- Perez-Chaparro, P.J.; Meuric, V.; De Mello, G.; Bonnaure-Mallet, M. Bacteremia of oral origin. Rev. Stomatol. Chir. Maxillofac. 2011, 112, 300–303. [Google Scholar] [CrossRef]
- Adam, E.L.; Siciliano, R.F.; Gualandro, D.M.; Calderaro, D.; Issa, V.S.; Rossi, F.; Caramelli, B.; Mansur, A.J.; Strabelli, T.M.V. Case series of infective endocarditis caused by Granulicatella species. Int. J. Infect. Dis. 2015, 31, 56–58. [Google Scholar] [CrossRef]
- Kanasi, E.; Dewhirst, F.; Chalmers, N.; Kent, R., Jr.; Moore, A.; Hughes, C.; Pradhan, N.; Loo, C.; Tanner, A.C.R. Clonal analysis of the microbiota of severe early childhood caries. Caries Res. 2010, 44, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Shimoyama, Y.; Ishikawa, T.; Kodama, Y.; Tajika, S.; Kimura, S. Contribution of different adherent properties of Granulicatella adiacens and Abiotrophia defectiva to their associations with oral colonization and the risk of infective endocarditis. J. Oral Sci. 2020, 62, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the normal bacterial flora of the oral cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef] [PubMed]
- Asikainen, S.; Dogan, B.; Turgut, Z.; Paster, B.J.; Bodur, A.; Oscarsson, J. Specified species in gingival crevicular fluid predict bacterial diversity. PLoS ONE 2010, 5, e13589. [Google Scholar] [CrossRef]
- Hsiao, W.W.L.; Li, K.L.; Liu, Z.; Jones, C.; Fraser-Liggett, C.M.; Fouad, A.F. Microbial transformation from normal oral microbiota to acute endodontic infections. BMC Genom. 2012, 13, 345. [Google Scholar] [CrossRef]
- Siqueira, J.F., Jr.; Rocas, I.N. Catonella morbi and Granulicatella adiacens: New species in endodontic infections. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontology 2006, 102, 259–264. [Google Scholar] [CrossRef]
- Ellis, T.N.; Kuehn, M.J. Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiol. Mol. Biol. Rev. 2010, 74, 81–94. [Google Scholar] [CrossRef]
- Kuehn, M.J.; Kesty, N.C. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes Dev. 2005, 19, 2645–2655. [Google Scholar] [CrossRef] [PubMed]
- Avila-Calderón, E.D.; Araiza-Villanueva, M.G.; Cancino-Diaz, J.C.; López-Villegas, E.O.; Sriranganathan, N.; Boyle, S.M.; Contreras-Rodríguez, A. Roles of bacterial membrane vesicles. Arch. Microbiol. 2015, 197, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.-H.; Choi, C.W.; Park, E.C.; Lee, S.-Y.; Kim, S.I. Isolation and proteomic characterization of bacterial extracellular membrane vesicles. Curr. Protein Pept. Sci. 2014, 15, 719–731. [Google Scholar] [CrossRef]
- Liu, Y.; Defourny, K.A.Y.; Smid, E.J.; Abee, T. Gram-Positive Bacterial Extracellular Vesicles and Their Impact on Health and Disease. Front. Microbiol. 2018, 9, 1502. [Google Scholar] [CrossRef] [PubMed]
- Sangiorgio, G.; Nicitra, E.; Bivona, D.; Bonomo, C.; Bonacci, P.; Santagati, M.; Musso, N.; Bongiorno, D.; Stefani, S. Interactions of Gram-Positive Bacterial Membrane Vesicles and Hosts: Updates and Future Directions. Int. J. Mol. Sci. 2024, 25, 2904. [Google Scholar] [CrossRef]
- Xu, Y.; Xie, C.; Liu, Y.; Qin, X.; Liu, J. An update on our understanding of Gram-positive bacterial membrane vesicles: Discovery, functions, and applications. Front. Cell. Infect. Microbiol. 2023, 13, 1273813. [Google Scholar] [CrossRef]
- Toyofuku, M.; Cárcamo-Oyarce, G.; Yamamoto, T.; Eisenstein, F.; Hsiao, C.-C.; Kurosawa, M.; Gademann, K.; Pilhofer, M.; Nomura, N.; Eberl, L. Prophage-triggered membrane vesicle formation through peptidoglycan damage in Bacillus subtilis. Nat. Commun. 2017, 8, 481. [Google Scholar] [CrossRef]
- Brookes, Z.L.; Belfield, L.A.; Ashworth, A.; Casas-Agustench, P.; Raja, M.; Pollard, A.J.; Bescos, R. Effects of chlorhexidine mouthwash on the oral microbiome. J. Dent. 2021, 113, 103768. [Google Scholar] [CrossRef]
- Baroudi, K.; Dagli, R.; Dagli, N.; Darwish, S. Oral Microbial Shift: Factors affecting the Microbiome and Prevention of Oral Disease. J. Contemp. Dent. Pract. 2016, 17, 90–96. [Google Scholar] [CrossRef]
- Kleine Bardenhorst, S.; Hagenfeld, D.; Matern, J.; Prior, K.; Harks, I.; Eickholz, P.; Lorenz, K.; Kim, T.-S.; Kocher, T.; Meyle, J.; et al. The role of the oral microbiota in the causal effect of adjunctive antibiotics on clinical outcomes in stage III-IV periodontitis patients. Microbiome 2024, 12, 220. [Google Scholar] [CrossRef]
- Alhazmi, Y.A.; Aljabri, M.Y.; Raafat, S.N.; Gomaa, S.M.; Shamel, M. Exploring the Effects of Low-Level Laser Therapy on the Cytocompatibility and Osteo/Odontogenic Potential of Gingival-Derived Mesenchymal Stem Cells: Preliminary Report. Appl. Sci. 2023, 13, 8490. [Google Scholar] [CrossRef]
- De Angelis, M.; Siragusa, S.; Campanella, D.; Di Cagno, R.; Gobbetti, M. Comparative proteomic analysis of biofilm and planktonic cells of Lactobacillus plantarum DB200. Proteomics 2015, 15, 2244–2257. [Google Scholar] [CrossRef] [PubMed]
- Dumitrache, A.; Klingeman, D.M.; Natzke, J.; Rodriguez, M., Jr.; Giannone, R.J.; Hettich, R.L.; Davison, B.H.; Brown, S.D. Specialized activities and expression differences for Clostridium thermocellum biofilm and planktonic cells. Sci Rep. 2017, 7, 43583. [Google Scholar] [CrossRef] [PubMed]
- Marinacci, B.; D’aMbrosio, C.; Vitale, I.; Di Sotto, A.; Cairone, F.; Spano, M.; Carradori, S.; Scaloni, A.; Gullì, M.; Puca, V.; et al. Biochemical and functional properties of vesicles from planktonic and biofilm phenotypes of Limosilactobacillus reuteri DSM 17938. Sci. Rep. 2025, 15, 18889. [Google Scholar] [CrossRef]
- Alkandari, S.A.; Bhardwaj, R.G.; Ellepola, A.; Karched, M. Proteomics of extracellular vesicles produced by Granulicatella adiacens, which causes infective endocarditis. PLoS ONE 2020, 15, e0227657. [Google Scholar] [CrossRef]
- Karched, M.; Bhardwaj, R.G.; Tiss, A.; Asikainen, S. Proteomic Analysis and Virulence Assessment of Granulicatella adiacens Secretome. Front. Cell. Infect. Microbiol. 2019, 9, 104. [Google Scholar] [CrossRef]
- Krzyściak, W.; Jurczak, A.; Kościelniak, D.; Bystrowska, B.; Skalniak, A. The virulence of Streptococcus mutans and the ability to form biofilms. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 499–515. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, W.; Wu, Z.; Lu, C. Reduced virulence is an important characteristic of biofilm infection of Streptococcus suis. FEMS Microbiol. Lett. 2011, 316, 36–43. [Google Scholar] [CrossRef]
- Warrier, A.; Satyamoorthy, K.; Murali, T.S. Quorum-sensing regulation of virulence factors in bacterial biofilm. Futur. Microbiol. 2021, 16, 1003–1021. [Google Scholar] [CrossRef]
- Karched, M.; Bhardwaj, R.G.; Inbamani, A.; Asikainen, S. Quantitation of biofilm and planktonic life forms of coexisting periodontal species. Anaerobe 2015, 35 Pt A, 13–20. [Google Scholar] [CrossRef]
- Effah, C.Y.; Ding, X.; Drokow, E.K.; Li, X.; Tong, R.; Sun, T. Bacteria-derived extracellular vesicles: Endogenous roles, therapeutic potentials and their biomimetics for the treatment and prevention of sepsis. Front. Immunol. 2024, 15, 1296061. [Google Scholar] [CrossRef]
- Peregrino, E.S.; Castañeda-Casimiro, J.; Vázquez-Flores, L.; Estrada-Parra, S.; Wong-Baeza, C.; Serafín-López, J.; Wong-Baeza, I. The Role of Bacterial Extracellular Vesicles in the Immune Response to Pathogens, and Therapeutic Opportunities. Int. J. Mol. Sci. 2024, 25, 6210. [Google Scholar] [CrossRef]
- Chakravarty, S.; Melton, C.N.; Bailin, A.; Yahr, T.L.; Anderson, G.G. Pseudomonas aeruginosa Magnesium Transporter MgtE Inhibits Type III Secretion System Gene Expression by Stimulating rsmYZ Transcription. J. Bacteriol. 2017, 199, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.d.l.P.; Wong, T.Y.; Damron, F.H.; Fernández, J.; Sisti, F. Cyclic di-GMP Regulates the Type III Secretion System and Virulence in Bordetella bronchiseptica. Infect. Immun. 2022, 90, e0010722. [Google Scholar] [CrossRef] [PubMed]
- Płaczkiewicz, J.; Adamczyk-Popławska, M.; Lasek, R.; Bącal, P.; Kwiatek, A. Inactivation of Genes Encoding MutL and MutS Proteins Influences Adhesion and Biofilm Formation by Neisseria gonorrhoeae. Microorganisms 2019, 7, 647. [Google Scholar] [CrossRef]
- Diene, S.M.; Pontarotti, P.; Azza, S.; Armstrong, N.; Pinault, L.; Chabrière, E.; Colson, P.; Rolain, J.-M.; Raoult, D. Origin, Diversity, and Multiple Roles of Enzymes with Metallo-β-Lactamase Fold from Different Organisms. Cells 2023, 12, 1752. [Google Scholar] [CrossRef]
- DeJesus, M.A.; Gerrick, E.R.; Xu, W.; Park, S.W.; Long, J.E.; Boutte, C.C.; Rubin, E.J.; Schnappinger, D.; Ehrt, S.; Fortune, S.M.; et al. Comprehensive Essentiality Analysis of the Mycobacterium tuberculosis Genome via Saturating Transposon Mutagenesis. mBio 2017, 8, e02133-16. [Google Scholar] [CrossRef]
- Figueiredo, T.A.; Ludovice, A.M.; Sobral, R.G. Contribution of peptidoglycan amidation to beta-lactam and lysozyme resistance in different genetic lineages of Staphylococcus aureus. Microb. Drug Resist. 2014, 20, 238–249. [Google Scholar] [CrossRef]
- Thanassi, J.A.; Hartman-Neumann, S.L.; Dougherty, T.J.; Dougherty, B.A.; Pucci, M.J. Identification of 113 conserved essential genes using a high-throughput gene disruption system in Streptococcus pneumoniae. Nucleic Acids Res. 2002, 30, 3152–3162. [Google Scholar] [CrossRef]
- Schwab, A.; Meyering, S.S.; Lepene, B.; Iordanskiy, S.; van Hoek, M.L.; Hakami, R.M.; Kashanchi, F. Extracellular vesicles from infected cells: Potential for direct pathogenesis. Front. Microbiol. 2015, 6, 1132. [Google Scholar] [CrossRef]
- Turner, S.; Raisley, B.; Roach, K.; Bajaña, S.; Munroe, M.E.; James, J.A.; Coggeshall, K.M.; Kovats, S. Gram-Positive Bacteria Cell Wall Peptidoglycan Polymers Activate Human Dendritic Cells to Produce IL-23 and IL-1β and Promote TH17 Cell Differentiation. Microorganisms 2023, 11, 173. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Zimmermann, C.; Kieseier, B.C.; Hartung, H.-P.; Hofstetter, H.H. Bacteria and their cell wall components uniformly co-activate interleukin-17-producing thymocytes. Clin. Exp. Immunol. 2014, 178, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Araujo, I.R.; Ferrari, T.C.; Teixeira-Carvalho, A.; Campi-Azevedo, A.C.; Rodrigues, L.V.; Guimaraes Junior, M.H.; Barros, T.L.S.; Gelape, C.L.; Sousa, G.R.; Nunes, M.C.P.N.; et al. Cytokine Signature in Infective Endocarditis. PLoS ONE 2015, 10, e0133631. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, Y.; Kubota, T.; Sasaguri, K.; Sato, S.; Suzuki, Y.; Kumada, H.; Umemoto, T. Inflammatory cytokine gene expression in human periodontal ligament fibroblasts stimulated with bacterial lipopolysaccharides. Infect. Immun. 1995, 63, 3576–3581. [Google Scholar] [CrossRef]
- Briaud, P.; Carroll, R.K. Extracellular Vesicle Biogenesis and Functions in Gram-Positive Bacteria. Infect. Immun. 2020, 88, 10–1128. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef]
- Toyofuku, M.; Nomura, N.; Eberl, L. Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 2019, 17, 13–24. [Google Scholar] [CrossRef]
- Thay, B.; Damm, A.; Kufer, T.A.; Wai, S.N.; Oscarsson, J. Aggregatibacter actinomycetemcomitans outer membrane vesicles are internalized in human host cells and trigger NOD1- and NOD2-dependent NF-kappaB activation. Infect. Immun. 2014, 82, 4034–4046. [Google Scholar] [CrossRef]
- Maldonado, R.; Wei, R.; Kachlany, S.; Kazi, M.; Balashova, N. Cytotoxic effects of Kingella kingae outer membrane vesicles on human cells. Microb. Pathog. 2011, 51, 22–30. [Google Scholar] [CrossRef]
- Iwabuchi, Y.; Yoshida, H.; Kamei, S.; Uematsu, T.; Saito, M.; Senpuku, H. Formation of Mono-Organismal and Mixed Staphylococcus aureus and Streptococcus mutans Biofilms in the Presence of NaCl. Microorganisms 2025, 13, 1118. [Google Scholar] [CrossRef]
- Yumoto, H.; Hirota, K.; Hirao, K.; Ninomiya, M.; Murakami, K.; Fujii, H.; Miyake, Y. The Pathogenic Factors from Oral Streptococci for Systemic Diseases. Int. J. Mol. Sci. 2019, 20, 4571. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).