Functional Ca2+ Channels between Channel Clusters are Necessary for the Propagation of IP3R-Mediated Ca2+ Waves


Abstract
1. Introduction
2. Results
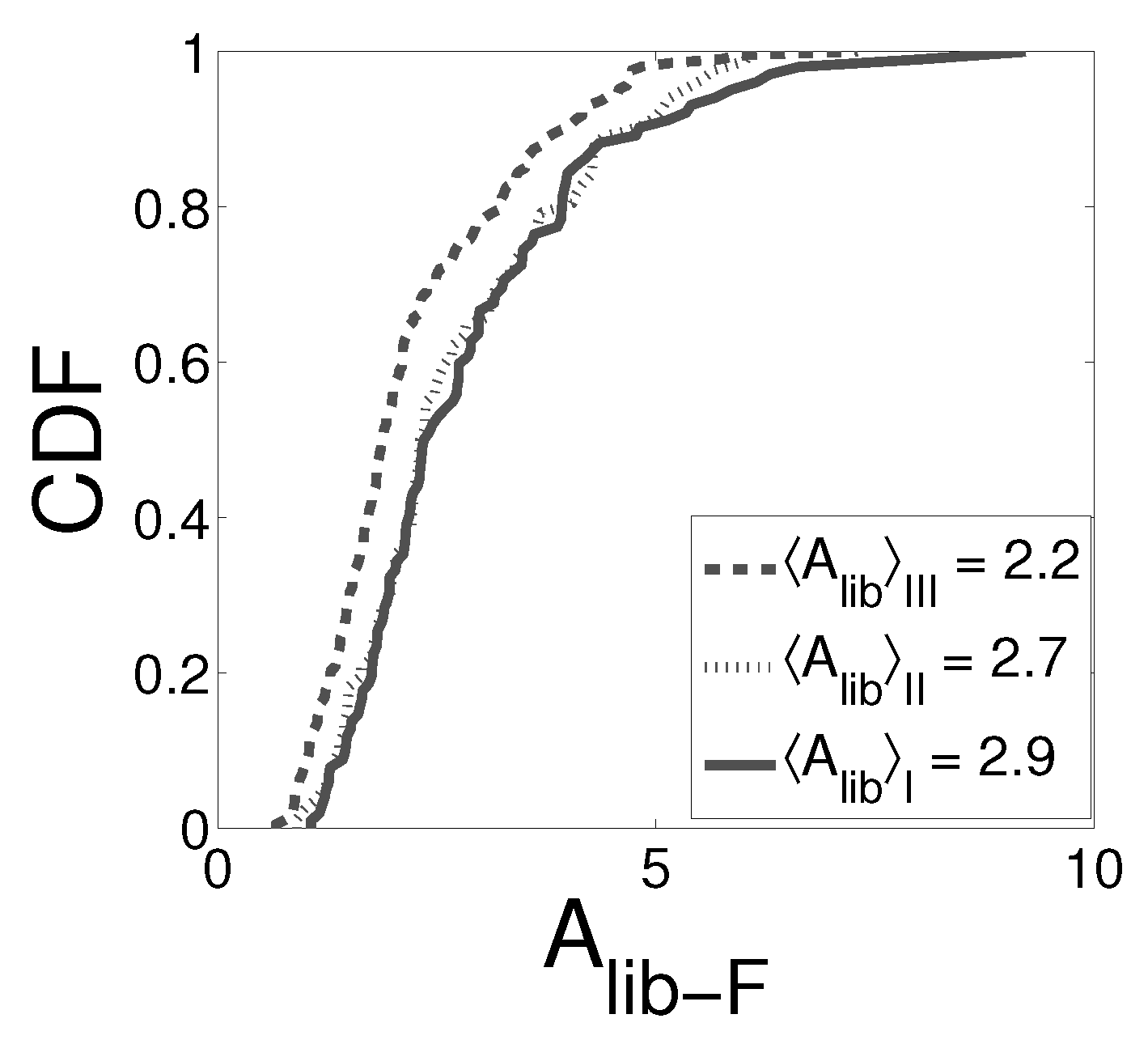
2.1. Experimental Results
2.2. Probabilistic Model to Analyze the Differences Observed in the Experimental Event Size Distributions for Different Values of
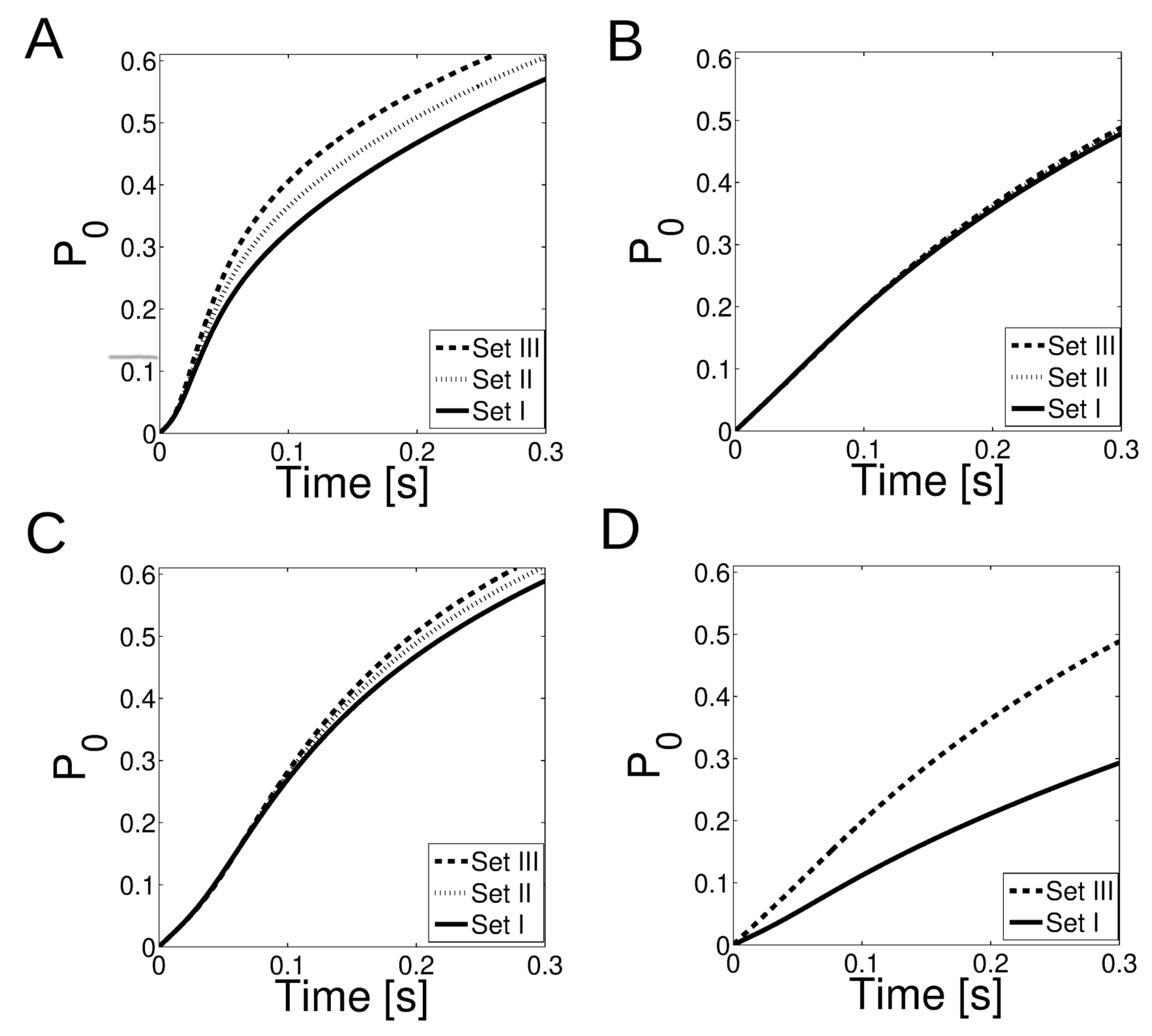
2.3. Numerical Simulations to Estimate the Probability That a Release Event from One (Primary) Cluster Induces the Release of Ca from Another (Secondary) Cluster
2.4. Combining the Estimates Derived from the Experiments and from the Numerical Simulations to Interpret the Changes Observed Experimentally
2.5. Changes in Basal Calcium Concentration, [Ca]
3. Discussion
4. Materials and Methods
4.1. Oocyte Preparation
4.2. Confocal Microscopy
4.3. Image Analysis
4.4. Basal Calcium Estimation
4.5. Numerical Simulations
Author Contributions
Funding
Conflicts of Interest
References
- Berridge, M.J.; Bootman, M.D.; Lipp, P. Calcium—A life and death signal. Nature 1998, 395, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Bootman, M.D.; Collins, T.J.; Peppiatt, C.M.; Prothero, L.S.; MacKenzie, L.; Smet, P.D.; Travers, M.; Tovey, S.C.; Seo, J.T.; Berridge, M.J.; et al. Calcium signalling—An overview. Semin. Cell Dev. Biol. 2001, 12, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Choe, C.U.; Ehrlich, B.E. The Inositol 1,4,5-Trisphosphate Receptor (IP3R) and Its Regulators: Sometimes Good and Sometimes Bad Teamwork. Sci. Signal. 2006, 2006, re15. [Google Scholar] [CrossRef] [PubMed]
- Foskett, J.K.; White, C.; Cheung, K.H.; Mak, D.O.D. Inositol Trisphosphate Receptor Ca2+ Release Channels. Physiol. Rev. 2007, 87, 593–658. [Google Scholar] [CrossRef] [PubMed]
- Fabiato, A. Calcium-induced release of calcium from the cardiac sarcoplasmic reticulum. Am. J. Physiol. 1983, 245, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.P.; Callamara, N.; Marchant, J.S.; Parker, I. A continuum of InsP3-mediated elementary Ca2+ signalling events in Xenopus oocyte. J. Physiol. 1998, 509, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.F.; Parker, I. Imaging the quantal substructure of single IP3R channel activity during Ca2+ puffs in intact mammalian cells. Proc. Natl. Acad. Sci. USA 2009, 106, 6404–6409. [Google Scholar] [CrossRef] [PubMed]
- Solovey, G.; Dawson, S.P. Intra-cluster percolation of calcium signals. PLoS ONE 2010, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Keizer, J.; Smith, G.D.; Ponce-Dawson, S.; Pearson, J.E. Saltatory Propagation of Ca2+ Waves by Ca2+ Sparks. Biophys. J. 1998, 75, 595–600. [Google Scholar] [CrossRef]
- Piegari, E.; Lopez, L.F.; Dawson, S.P. Using two dyes to observe the competition of Ca2+ trapping mechanisms and their effect on intracellular Ca2+ signals. Phys. Biol. 2018, 15, 066006. [Google Scholar] [CrossRef] [PubMed]
- Paredes, R.M.; Etzler, J.C.; Watts, L.T.; Zheng, W.; Lechleiter, J.D. Chemical calcium indicators. Methods 2008, 46, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Dargan, S.L.; Parker, I. Buffer kinetics shape the spatiotemporal patterns of IP3-evoked Ca2+ signals. J. Physiol. 2003, 553, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Dargan, S.L.; Schwaller, B.; Parker, I. Spatiotemporal patterning of IP3-mediated Ca2+ signals in Xenopus oocytes by Ca2+-binding proteins. J. Physiol. 2004, 556, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Piegari, E.; Sigaut, L.; Ponce Dawson, S. Ca2+ images obtained in different experimental conditions shed light on the spatial distribution of IP3 receptors that underlie Ca2+ puffs. Cell Calcium 2015, 57, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Solovey, G.; Fraiman, D.; Dawson, S.P. Mean field strategies induce unrealistic nonlinearities in calcium puffs. Front. Physiol. 2011, 2, 1–11. [Google Scholar] [CrossRef]
- Callamaras, N.; Parker, I. Phasic characteristic of elementary Ca2+ release sites underlies quantal responses to IP3. EMBO J. 2000, 19, 3608–3617. [Google Scholar] [CrossRef] [PubMed]
- Sigaut, L.; Barella, M.; Espada, R.; Ponce, M.L.; Dawson, S.P. Custom-made modification of a commercial confocal microscope to photolyze caged compounds using the conventional illumination module and its application to the observation of Inositol 1,4,5-trisphosphate-mediated calcium signals. J. Biomed. Opt. 2011, 16, 066013. [Google Scholar] [CrossRef] [PubMed]
- Piegari, E.; Lopez, L.; Perez Ipiña, E.; Ponce Dawson, S. Fluorescence fluctuations and equivalence classes of Ca2+ imaging experiments. PLoS ONE 2014, 9, e95860. [Google Scholar] [CrossRef] [PubMed]
- Youngt, G.W.D.E.; Keizer, J. A single pool. Nature 1992, 89, 9895–9899. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | [Fluo-4] () | [Rhod-2] () | [EGTA] () |
|---|---|---|---|
| Set I | 36 | 90 | 90 |
| Set II | 36 | 36 | 90 |
| Set III | 36 | 0 | 90 |
| Parameter | Abbreviature | Values |
|---|---|---|
| Number of IPRs in the source | 1, 10, 50 | |
| Distance to the Ca source | d | (0.4–1.5) m |
| Number of sensing IPRs | 1, 5 | |
| Rhod-2 concentration | 0, 90 M | |
| Velocity of propagation | V | 10, 20 m/s |
| Parameter | Value | Units |
|---|---|---|
| Free Calcium | ||
| 220 | ||
| 0.05–0.1 | ||
| Calcium dye Fluo-4-dextran | ||
| 15 | ||
| 240 | ||
| 180 | ||
| 36 | ||
| Calcium dye Rhod-2-dextran | ||
| 15 | ||
| 70 | ||
| 130 | ||
| 0, 36, 90 | ||
| Exogenous buffer EGTA | ||
| 80 | ||
| 5 | ||
| 0,75 | ||
| 90 | ||
| Endogenous immobile buffer | ||
| 0 | ||
| 400 | ||
| 800 | ||
| 300 | ||
| Pump | ||
| 0.1 | ||
| 0.9 | ||
| Source | ||
| 1, 10, 50 | - | |
| 20 | ms | |
| 0.1 | pA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piegari, E.; Ponce Dawson, S. Functional Ca2+ Channels between Channel Clusters are Necessary for the Propagation of IP3R-Mediated Ca2+ Waves. Math. Comput. Appl. 2019, 24, 61. https://doi.org/10.3390/mca24020061
Piegari E, Ponce Dawson S. Functional Ca2+ Channels between Channel Clusters are Necessary for the Propagation of IP3R-Mediated Ca2+ Waves. Mathematical and Computational Applications. 2019; 24(2):61. https://doi.org/10.3390/mca24020061
Chicago/Turabian StylePiegari, Estefanía, and Silvina Ponce Dawson. 2019. "Functional Ca2+ Channels between Channel Clusters are Necessary for the Propagation of IP3R-Mediated Ca2+ Waves" Mathematical and Computational Applications 24, no. 2: 61. https://doi.org/10.3390/mca24020061
APA StylePiegari, E., & Ponce Dawson, S. (2019). Functional Ca2+ Channels between Channel Clusters are Necessary for the Propagation of IP3R-Mediated Ca2+ Waves. Mathematical and Computational Applications, 24(2), 61. https://doi.org/10.3390/mca24020061