
Effects of Growth Parameters on the Analysis of Aspergillus flavus Volatile Metabolites

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Growth Media
2.2. Fungal Isolates and Growth
2.3. Selection of SPME Fibers
2.4. Effects of Growth Parameters on the MVOCs’ Production
2.5. GC-MS Analysis
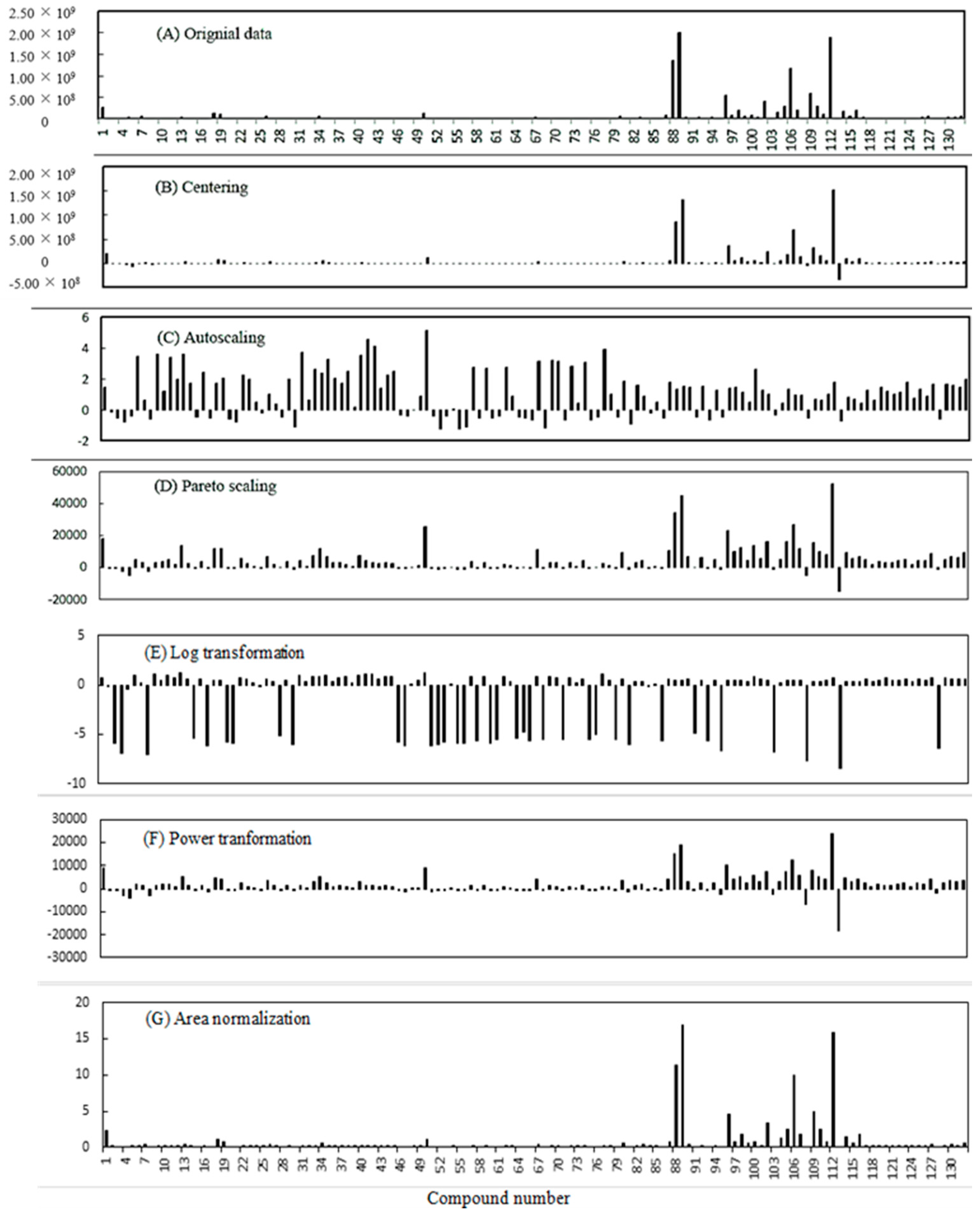
2.6. GC-MS MVOCs Data Manipulation
2.7. Data Analysis
3. Results and Discussion
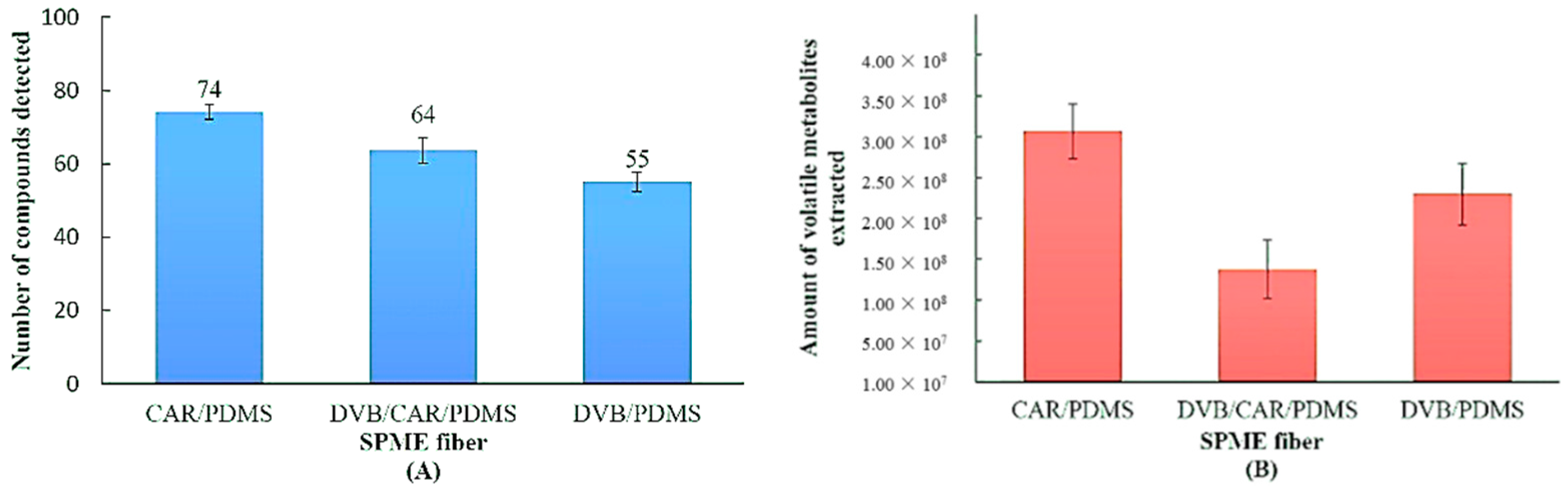
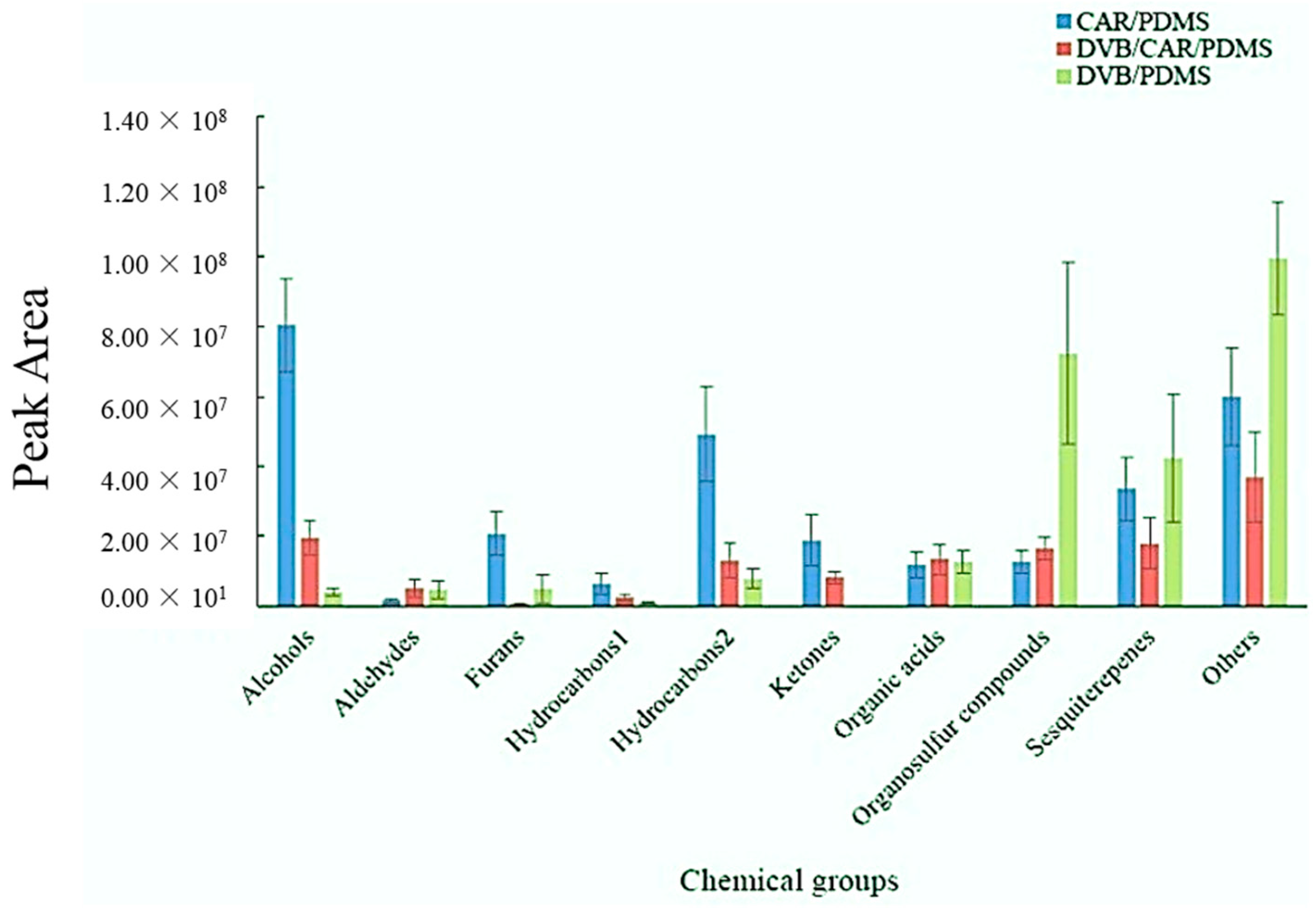
3.1. Evaluation of SPME Fiber on Metabolic Profiling
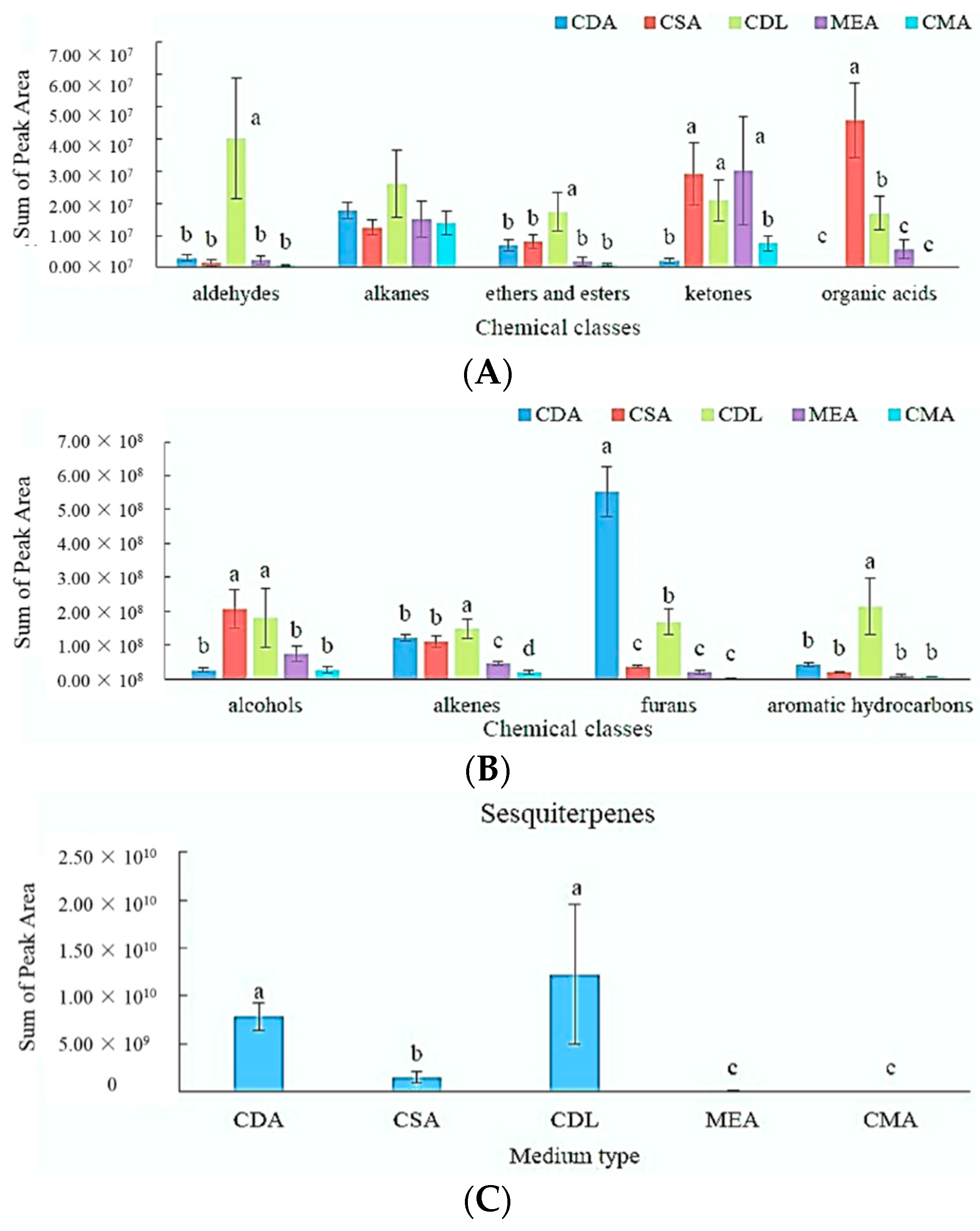
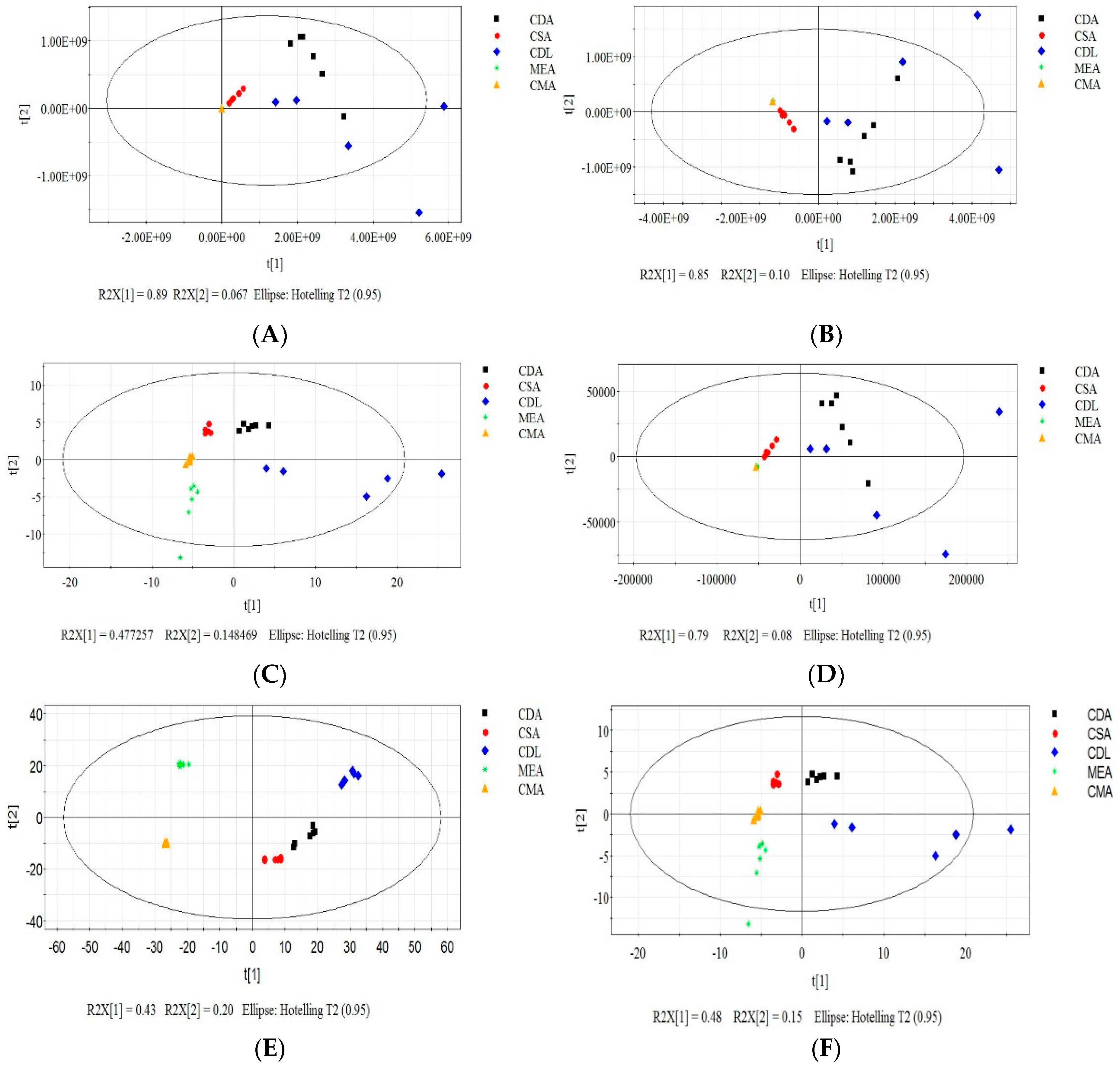
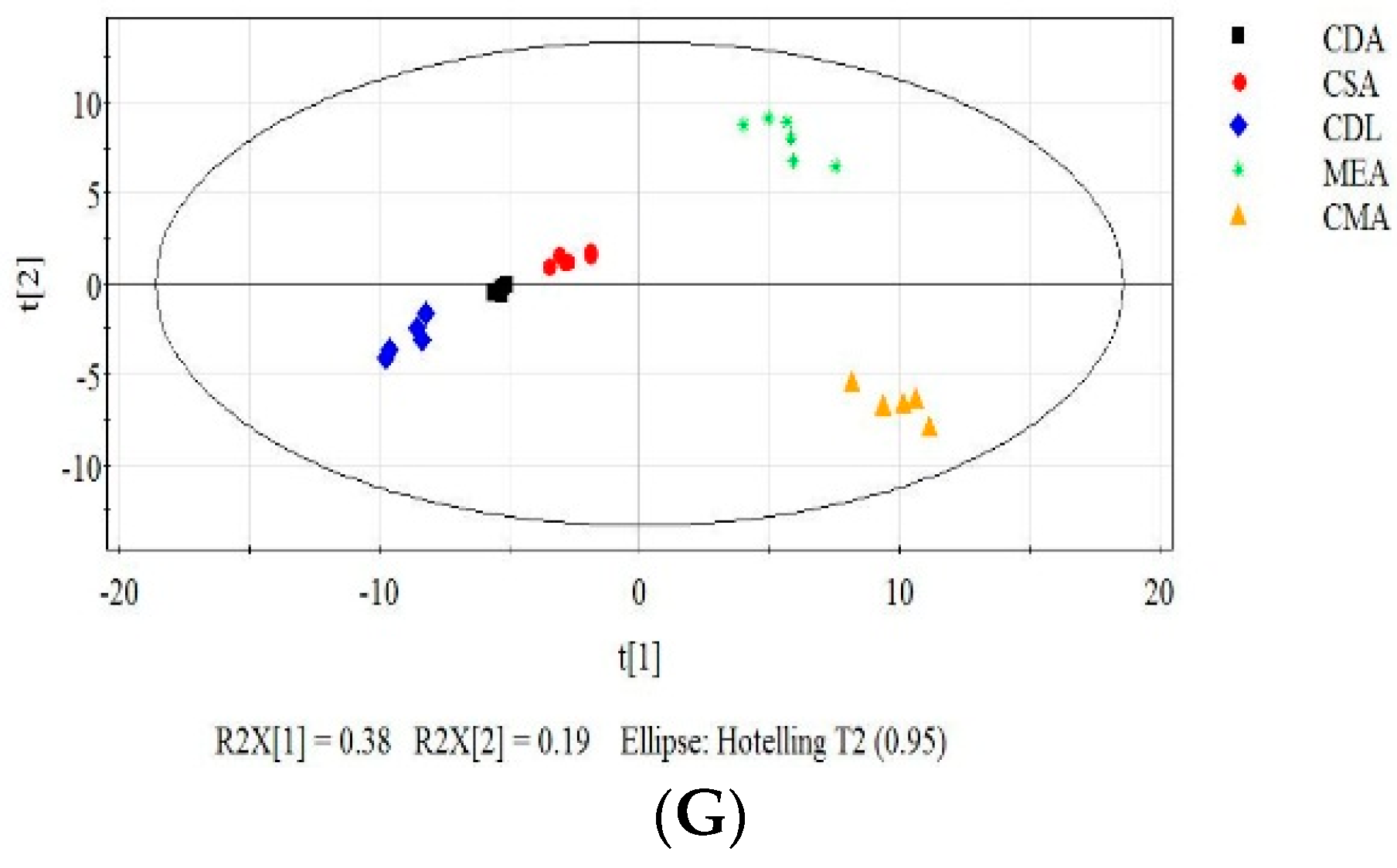
3.2. Effect of the Growth Substrates on MVOCs’ Production
3.3. Effect of the Concentration of Spore Suspension on MVOCs’ Production
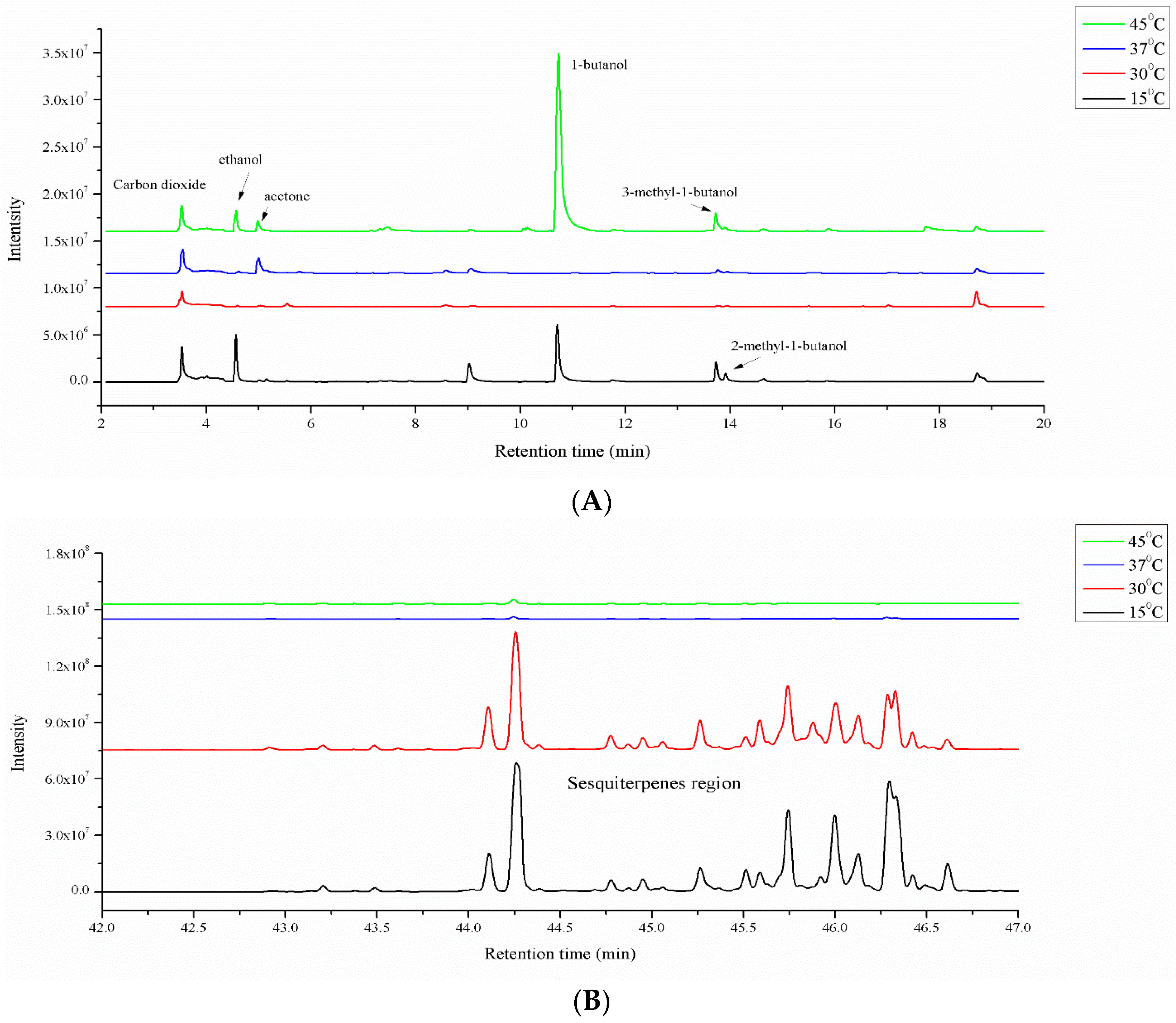
3.4. Effect of Temperature on MVOCs’ Production
3.5. Effect of Data Pretreatment Methods
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gourama, H.; Bullerman, L.B. Aspergillus flavus and Aspergillus parasiticus: Aflatoxigenic fungi of concern in foods and feeds: A review. J. Food Prot. 1995, 58, 1395–1404. [Google Scholar]
- Lamb, M.C.; Sternitzke, D.A. Cost of aflatoxin to the farmer, buying point, and sheller segments of the southeast united states peanut industry. Pean. Sci. 2001, 28, 59–63. [Google Scholar] [CrossRef]
- Korpi, A.; Järnberg, J.; Pasanen, A.-L. Microbial volatile organic compounds. Crit. Rev. Toxicol. 2009, 39, 139–193. [Google Scholar] [CrossRef] [PubMed]
- Ryan, T.J.; Beaucham, C. Dominant microbial volatile organic compounds in 23 US homes. Chemosphere 2013, 90, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Moularat, S.; Hulin, M.; Robine, E.; Annesi-Maesano, I.; Caillaud, D. Airborne fungal volatile organic compounds in rural and urban dwellings: Detection of mould contamination in 94 homes determined by visual inspection and airborne fungal volatile organic compounds method. Sci. Total. Environ. 2011, 409, 2005–2009. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Kimura, T.; Tanaka, H.; Kaneko, S.; Ichii, S.; Kiuchi, M.; Suzuki, T. Analysis of volatile metabolites emitted by soil-derived fungi using head space solid-phase microextraction/gas chromatography/mass spectrometry: I. Aspergillus fumigatus, Aspergillus nidulans, Fusarium solani and Penicillium paneum. Surf. Interface Anal. 2012, 44, 694–698. [Google Scholar] [CrossRef]
- Kuske, M.; Romain, A.-C.; Nicolas, J. Microbial volatile organic compounds as indicators of fungi. Can an electronic nose detect fungi in indoor environments? Build. Environ. 2005, 40, 824–831. [Google Scholar] [CrossRef]
- Van Lancker, F.; Adams, A.; Delmulle, B.; de Saeger, S.; Moretti, A.; van Peteghem, C.; de Kimpe, N. Use of headspace SPME-GC-MS for the analysis of the volatiles produced by indoor molds grown on different substrates. J. Environ. Monitor. 2008, 10, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Jurjevic, Z.; Rains, G.C.; Wilson, D.M.; Lewis, W. Volatile metabolites associated with one aflatoxigenic and one nontoxigenic Aspergillus flavus strain grown on two different substrates. Phytopathol. Mediterr. 2009, 47, 266–271. [Google Scholar]
- Moularat, S.; Robine, E. Process for determining mycotoxin production from a specific chemical fingerprint. Google Patents US 20130244272 A1, 2012. [Google Scholar]
- Polizzi, V.; Adams, A.; Malysheva, S.V.; de Saeger, S.; van Peteghem, C.; Moretti, A.; Picco, A.M.; de Kimpe, N. Identification of volatile markers for indoor fungal growth and chemotaxonomic classification of Aspergillus species. Fungal Biol. 2012, 116, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Faubert, P.; Hagen, M.; zu Castell, W.; Polle, A.; Schnitzler, J.-P.; Rosenkranz, M. Volatile profiles of fungi—Chemotyping of species and ecological functions. Fungal Genet. Biol. 2013, 54, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lavine, B.K.; Mirjankar, N.; LeBouf, R.; Rossner, A. Prediction of mold contamination from microbial volatile organic compound profiles using head space gas chromatography/mass spectrometry. Microchem. J. 2012, 103, 119–124. [Google Scholar] [CrossRef]
- Griffin, J.L.; Nicholls, A.W. Metabolomics as a functional genomic tool for understanding lipid dysfunction in diabetes, obesity and related disorders. Pharmgenomics Pers. Med. 2006, 7, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Quinones, M.P.; Kaddurah-Daouk, R. Metabolomics tools for identifying biomarkers for neuropsychiatric diseases. Neurobiol. Dis. 2009, 35, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Kaddurah-Daouk, R.; Kristal, B.S.; Weinshilboum, R.M. Metabolomics: A global biochemical approach to drug response and disease. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 653–683. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Applications of metabolomics in drug discovery and development. Drugs. R D 2008, 9, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Viant, M.R.; Tjeerdema, R.S. Metabolomics: Methodologies and applications in the environmental sciences. J. Pestic. Sci. 2006, 31, 245–251. [Google Scholar] [CrossRef]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef] [PubMed]
- Holmquist, G.; Walker, H.; Stahr, H. Influence of Temperature, pH, Water Activity and Antifungal Agents on Growth of Aspergillus Flavus and A. Parasiticus. J. Food Sci. 1983, 48, 778–782. [Google Scholar] [CrossRef]
- Polizzi, V.; Adams, A.; de Saeger, S.; van Peteghem, C.; Moretti, A.; de Kimpe, N. Influence of various growth parameters on fungal growth and volatile metabolite production by indoor molds. Sci. Total Environ. 2012, 414, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Grintzalis, K.; Vernardis, S.I.; Klapa, M.I.; Georgiou, C.D. Role of Oxidative Stress in Sclerotial Differentiation and Aflatoxin B1 Biosynthesis in Aspergillus Flavus. Appl. Environ. Microbiol. 2014, 80, 5561–5571. [Google Scholar] [CrossRef] [PubMed]
- López-Malo, A.; Alzamora, S.M.; Argaiz, A. Effect of vanillin concentration, pH and incubation temperature on Aspergillus flavus, Aspergillus niger, Aspergillus ochraceus and Aspergillus parasiticus growth. Food Microbiol. 1997, 14, 117–124. [Google Scholar] [CrossRef]
- Joffe, A.; Lisker, N. Effects of light, temperature, and pH value on aflatoxin production in vitro. Appl. Environ. Microbiol. 1969, 18, 517–518. [Google Scholar]
- Calenic, B.; Miricescu, D.; Greabu, M.; Andrey, V.K.; Troppmair, J.; Ruzsanyi, V.; Amann, A. Oxidative stress and volatile organic compounds: Interplay in pulmonary, cardio-vascular, digestive tract systems and cancer. Open Chem. 2015, 13, 1020–1030. [Google Scholar] [CrossRef]
- Sunesson, A.; Vaes, W.; Nilsson, C.; Blomquist, G.; Andersson, B.; Carlson, R. Identification of volatile metabolites from five fungal species cultivated on two media. Appl. Environ. Microbiol. 1995, 61, 2911–2918. [Google Scholar] [PubMed]
- Betancourt, D.A.; Krebs, K.; Moore, S.A.; Martin, S.M. Microbial volatile organic compound emissions from Stachybotrys chartarum growing on gypsum wallboard and ceiling tile. BMC Microbiol. 2013, 13, 283. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.O.; Frisvad, J.C. Comparison of different methods for collection of volatile chemical markers from fungi. J. Microbiol. Methods 1995, 24, 135–144. [Google Scholar] [CrossRef]
- Camara, M.; Gharbi, N.; Cocco, E.; Guignard, C.; Behr, M.; Evers, D.; Orlewski, P. Fast screening for presence of muddy/earthy odorants in wine and in wine must using a hyphenated gas chromatography-differential ion mobility spectrometry (GC/DMS). Int. J. Ion. Mobil. Spectrom. 2011, 14, 39–47. [Google Scholar] [CrossRef]
- Demyttenaere, J.C.R.; Moriña, R.M.; Sandra, P. Monitoring and fast detection of mycotoxin-producing fungi based on headspace solid-phase microextraction and headspace sorptive extraction of the volatile metabolites. J. Chromatogr. A 2003, 985, 127–135. [Google Scholar] [CrossRef]
- Jeleń, H. Use of solid phase microextraction (SPME) for profiling fungal volatile metabolites. Lett. Appl. Microbiol. 2003, 36, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Wood-Jones, A.; Wang, W.; Vanlangenberg, C.; Jones, D.; Gower, J.; Simmons, P.; Baird, R.; Mlsna, T. Monitoring MVOC profiles over time from isolates of Aspergillus flavus using SPME GC-MS. J. Agric. Chem. Environ. 2014, 3, 48–63. [Google Scholar] [CrossRef]
- Kluger, B.; Zeilinger, S.; Wiesenberger, G.; Schöfbeck, D.; Schuhmacher, R. Detection and identification of fungal microbial volatile organic compounds by HS-SPME-GC-MS. In Laboratory Protocols in Fungal Biology; Springer: Berlin, Geramny, 2013; pp. 455–465. [Google Scholar]
- Deshmukh, Y.; Khare, P.; Patra, D.; Nadaf, A.B. HS-SPME-GC-FID method for detection and quantification of Bacillus cereus ATCC 10702 mediated 2-acetyl-1-pyrroline. Biotechnol. Prog. 2014, 30, 1356–1363. [Google Scholar] [CrossRef] [PubMed]
- Shirey, R.E. Optimization of extraction conditions and fiber selection for semivolatile analytes using solid-phase microextraction. J. Chromatogr. Sci. 2000, 38, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Stoppacher, N.; Kluger, B.; Zeilinger, S.; Krska, R.; Schuhmacher, R. Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride by HS-SPME-GC-MS. J. Microbiol. Methods 2010, 81, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, M.T.; Mayahi, S.; Denning, D.W. A study on Aspergillus species in houses of asthmatic patients from Sari City, Iran and a brief review of the health effects of exposure to indoor Aspergillus. Environ. Monit. Assess. 2010, 168, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, K.; Larsen, K.; Simkus, M. Volatile metabolites from indoor molds grown on media containing wood constituents. Environ. Sci. Pollut. Res. 2003, 10, 206–208. [Google Scholar] [CrossRef]
- Radványi, D.; Gere, A.; Jókai, Z.; Fodor, P. Rapid evaluation technique to differentiate mushroom disease-related moulds by detecting microbial volatile organic compounds using HS-SPME-GC-MS. Anal. Bioanal. Chem. 2015, 407, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Zscheppank, C.; Wiegand, H.; Lenzen, C.; Wingender, J.; Telgheder, U. Investigation of volatile metabolites during growth of Escherichia coli and Pseudomonas aeruginosa by needle trap-GC-MS. Anal. Bioanal. Chem. 2014, 406, 6617–6628. [Google Scholar] [CrossRef] [PubMed]
- Koek, M.M.; Muilwijk, B.; van der Werf, M.J.; Hankemeier, T. Microbial metabolomics with gas chromatography/mass spectrometry. Anal. Chem. 2006, 78, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Matysik, S.; Herbarth, O.; Mueller, A. Determination of microbial volatile organic compounds (MVOCs) by passive sampling onto charcoal sorbents. Chemosphere 2009, 76, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Syhre, M.; Scotter, J.M.; Chambers, S.T. Investigation into the production of 2-Pentylfuran by Aspergillus fumigatus and other respiratory pathogens in vitro and human breath samples. Med. Mycol. 2008, 46, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, K.; Nielsen, K.F.; Din, S.U. Patterns of volatile metabolites and nonvolatile trichothecenes produced by isolates of Stachybotrys, Fusarium, Trichoderma, Trichothecium and Memnoniella. Environ. Sci. Pollut. Res. 2003, 10, 162–166. [Google Scholar] [CrossRef]
- Meruva, N.; Penn, J.; Farthing, D. Rapid identification of microbial VOCs from tobacco molds using closed-loop stripping and gas chromatography/time-of-flight mass spectrometry. J. Ind Microbiol. Biotechnol. 2004, 31, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Ruzsanyi, V.; Baumbach, J.; Eiceman, G. Detection of the mold markers using ion mobility spectrometry. Int. J. Ion. Mobil. Spectrom. 2003, 6, 53–57. [Google Scholar]
- Tiebe, C.; Hübert, T.; Koch, B.; Ritter, U.; Stephan, I. Investigation of gaseous metabolites from moulds by ion mobility spectrometry (IMS) and gas chromatography-mass spectrometry (GC-MS). Int. J. Ion Mobil. Spectrom. 2010, 13, 17–24. [Google Scholar] [CrossRef]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Bro, R.; Smilde, A.K. Centering and scaling in component analysis. J. Chemom. 2003, 17, 16–33. [Google Scholar] [CrossRef]
- Kvalheim, O.M.; Brakstad, F.; Liang, Y. Preprocessing of analytical profiles in the presence of homoscedastic or heteroscedastic noise. Anal. Chem. 1994, 66, 43–51. [Google Scholar] [CrossRef]
- Thorn, R.M.; Reynolds, D.M.; Greenman, J. Multivariate analysis of bacterial volatile compound profiles for discrimination between selected species and strains in vitro. J. Microbiol. Methods 2011, 84, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Claeson, A.S.; Nordin, S.; Sunesson, A.L. Effects on perceived air quality and symptoms of exposure to microbially produced metabolites and compounds emitted from damp building materials. Indoor Air 2009, 19, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Pinzari, F.; Fanelli, C.; Canhoto, O.; Magan, N. Electronic nose for the early detection of moulds in libraries and archives. Indoor Built Environ. 2004, 13, 387–395. [Google Scholar] [CrossRef]
- Reddy, T.; Viswanathan, L.; Venkitasubramanian, T. Factors affecting aflatoxin production by Aspergillus parasiticus in a chemically defined medium. Microbiology 1979, 114, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, P.; Gullberg, J.; Nordström, A.; Kusano, M.; Kowalczyk, M.; Sjöström, M.; Moritz, T. A strategy for identifying differences in large series of metabolomic samples analyzed by GC/MS. Anal. Chem. 2004, 76, 1738–1745. [Google Scholar] [CrossRef] [PubMed]
- Pawliszyn, J. Handbook of Solid Phase Microextraction; Elsevier: Philadelphia, PA, USA, 2011. [Google Scholar]
- Rega, B.; Fournier, N.; Guichard, E. Solid phase microextraction (SPME) of orange juice flavor: Odor representativeness by direct gas chromatography olfactometry (D-GC-O). J. Agric. Food Chem. 2003, 51, 7092–7099. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.O.; Frisvad, J.C. Characterization of volatile metabolites from 47 Penicillium taxa. Mycol. Res. 1995, 99, 1153–1166. [Google Scholar] [CrossRef]
- Kahlos, K.; Kiviranta, J.L.; Hiltunen, R.V. Volatile constituents of wild and in vitro cultivated Gloeophyllum odoratum. Phytochemistry 1994, 36, 917–922. [Google Scholar] [CrossRef]
- Wheatley, R.; Hackett, C.; Bruce, A.; Kundzewicz, A. Effect of substrate composition on production of volatile organic compounds from Trichoderma spp. inhibitory to wood decay fungi. Int. Biodeterior. Biodegrad. 1997, 39, 199–205. [Google Scholar] [CrossRef]
- Gao, P.; Korley, F.; Martin, J.; Chen, B.T. Determination of unique microbial volatile organic compounds produced by five Aspergillus species commonly found in problem buildings. Am. Ind. Hyg. Assoc. J. 2002, 63, 135–140. [Google Scholar] [CrossRef]
- Schindler, A.F.; Palmer, J.G.; Eisenberg, W.V. Aflatoxin production by Aspergillus flavus as related to various temperatures. J. Appl. Microbiol. 1967, 15, 1006–1009. [Google Scholar]
- Karunaratne, A.; Bullerman, L.B. Interactive effects of spore load and temperature on aflatoxin production. J. Food Prot. 1990, 53, 227–236. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Substrate Name | Abbreviation | pH | Media Concentration | Ingredients |
|---|---|---|---|---|
| Corn meal agar | CMA | 6.0 ± 0.2 | 17 g/L | 2 g corn meal, infusion from solid, 15 g agar |
| Czapek solution agar | CSA | 7.3 ± 0.2 | 49 g/L | 30 g saccharose, 2.0 g NaNO3, 1.0 g K2HPO4, 0.5 g MgSO4, 0.5 g KCl, 0.01g FeSO4, and 15 g agar |
| Chemical defined agar | CDA | - | 60.28 g/L | 30 g sucrose, 10 g asparagine, 3.5 g (NH4)2SO4, 1 g KH2PO4, 500 mg MgSO4, 200 mg CaCl2, 10 mg ZnSO4·7H2O, 5 mg MnCl2, 2 mg FeSO4, and 15 g agar |
| Chemical defined liquid | CDL | - | 45.28 g/L | 30 g sucrose, 10 g asparagine, 3.5 g (NH4)2SO4, 1 g KH2PO4, 500 mg MgSO4, 200 mg CaCl2, 10 mg ZnSO4·7H2O, 5 mg MnCl2, 2 mg FeSO4 |
| Malt extract agar | MEA | 4.7 ± 0.2 | 33.6 g/L | 12.75 g maltose, 2.75 g dextrin, 2.35 g glycerol, 0.78 g peptone and 15.0 g agar |
| Effect Factors | Growth Parameters | |
|---|---|---|
| Media | CMA, CSA, CDA, CDL and MEA | |
| Concentration of spore suspension (spores/mL) | High | 1.2 × 107 |
| Medium | 2.4 × 106 | |
| Low | 4.8 × 105 | |
| Toothpick | - | |
| Temperature (°C) | 15, 30, 37, 45 | |
| No. | Compound Name | R.T. (min) b | Peak Area RSD% | Peak Area% RSD% | ||||
|---|---|---|---|---|---|---|---|---|
| CAR | DVB CAR | DVB | CAR | DVB CAR | DVB | |||
| 4 | 1,4-Pentadiene | 5.543 | 48.8 | 61.9 | - | 40.4 | 40.0 | - |
| 9 | Propanoic acid, 2-methyl-, anhydride | 8.313 | 68.0 | 31.1 | - | 64.1 | 65.4 | - |
| 10 | Furan, 2-methyl- | 8.568 | 38.4 | 20.3 | - | 30.1 | 22.4 | - |
| 11 | 1-Propanol, 2-methyl- | 9.046 | 30.4 | 32.6 | - | 29.7 | 30.8 | - |
| 15 | 1-Butanol, 3-methyl- | 13.825 | 52.6 | 51.3 | - | 57.7 | 74.6 | - |
| 16 | 1-Butanol, 2-methyl- | 13.943 | 37.8 | 34.3 | - | 41.8 | 32.3 | - |
| 18 | Toluene | 15.501 | 13.8 | 15.9 | - | 25.1 | 26.2 | - |
| 22 | Styrene | 21.901 | 23.8 | 17.3 | 19.3 | 35.7 | 24.5 | 19.7 |
| 36 | Undecane, 2,6-dimethyl- | 38.682 | 73.5 | 37.2 | 43.5 | 48.0 | 37.0 | 20.0 |
| 52 | 2,4,4,6,6,8,8-Heptamethyl-2-nonene | 43.784 | 26.0 | 27.0 | 31.5 | 15.3 | 40.0 | 11.7 |
| 56 | β-Elemene | 44.256 | 26.8 | 24.7 | 30.2 | 27.1 | 23.8 | 7.8 |
| 64 | α-Selinene | 45.591 | 34.7 | 46.5 | 44.5 | 39.6 | 0.6 | 42.7 |
| 68 | Cedrene | 46.060 | 29.5 | 37.6 | 28.4 | 14.6 | 20.4 | 29.0 |
| 70 | Calamenene | 46.338 | 27.3 | 52.4 | 48.8 | 23.7 | 45.1 | 29.0 |
| 72 | π-Calacorene | 46.627 | 17.0 | 32.6 | 36.5 | 22.8 | 21.7 | 23.1 |
| Average RSD% a | - | 56.7 | 40.9 | 49.8 | 56.7 | 52.08 | 39.5 | |
| No. | MVOC a | Chemical Classes | R.T. (min) | Media (Peak Area × 105) | ||||
|---|---|---|---|---|---|---|---|---|
| CDA | CSA | CDL | MEA | CMA | ||||
| 1 | Ethanol | alcohol | 4.595 | 15.9 ± 7.29 b | 590 ± 239 | 2441 ± 2797 | 50.6 ± 31.8 | 188 ± 92 |
| 2 | Acetone | ketone | 5.022 | - | 282 ± 97 | 142 ± 155 | 266 ± 151 | 71.1 ± 24.7 |
| 4 | Isopropyl Alcohol | alcohol | 5.149 | 4471 ± 601 | 312 ± 66 | - | 78.4 ± 46.3 | 57.03 ± 7.79 |
| 7 | 1,4-Pentadiene | alkene | 5.547 | - | 700 ± 153 | 466 ± 76 | 211 ± 103 | 12.7 ± 2.91 |
| 18 | Furan, 2-methyl- | furan | 8.567 | 131.7 ± 41.4 | 360 ± 45 | 1136 ± 317 | 172 ± 66.2 | 11.71 ± 3.08 |
| 29 | Toluene | aromatic hydrocarbons | 15.506 | 12.9 ± 4.35 | 24.7 ± 7.95 | 101 ± 27.2 | 24.3 ± 3.35 | 8.63 ± 1.71 |
| 36 | Styrene | aromatic hydrocarbons | 21.984 | - | 37.7 ± 8.33 | 91.6 ± 15.7 | 42.3 ± 10.1 | 23.7 ± 10.6 |
| 50 | D-Limonene | aromatic hydrocarbons | 30.733 | - | 34.9 ± 3.24 | 334 ± 548 | 3.76 ± 2.38 | 8.23 ± 2.14 |
| 54 | Dodecane | alkane | 33.053 | 14.1 ± 2.11 | 46.4 ± 16 | 46.5 ± 14.8 | 96.6 ± 99.6 | 30.9 ± 5.98 |
| 63 | 2,2-Dimethylheptane-3,5-dione | ketone | 35.810 | - | 10.1 ± 2.82 | 40.6 ± 25.8 | 10.4 ± 13.1 | 5.38 ± 0.64 |
| 67 | Decanal | aldehyde | 39.186 | 6.65 ± 0.864 | 14.9 ± 8.36 | 238 ± 178 | 11.1 ± 8.25 | 8.58 ± 2.82 |
| 69 | 1,3,7-Octatriene, 3,7-dimethyl-, | alkene | 40.716 | 12.4 ± 2.45 | - | 13.5 ± 11.9 | 13.5 ± 7.87 | - |
| 73 | Heptacosane | alkane | 42.160 | 19.6 ± 6.76 | - | 11.2 ± 10.1 | 53.5 ± 22.2 | 9.73 ± 4.97 |
| 74 | δ-Cadinene | sesquiterpene | 42.380 | - | 38.1 ± 17.2 | 29.7 ± 32.2 | 11 ± 4.47 | 20.7 ± 10.8 |
| 81 | (Z)-2-Hexadecene | alkene | 43.313 | 84.9 ± 22.9 | 19.7 ± 5.31 | - | 33.7 ± 8.69 | 15 ± 7 |
| 82 | trans-α-Bergamotene | sesquiterpene | 43.379 | 282 ± 77 | 38.7 ± 25.8 | 76.7 ± 37.3 | 13.7 ± 5.65 | 16.1 ± 8.52 |
| 83 | α-Cubebene | sesquiterpene | 43.489 | 100 ± 27 | 81.0 ± 18.3 | 542 ± 441 | 188 ± 46.2 | - |
| 84 | 3-Hexadecene, (Z)- | alkene | 43.619 | 61.3 ± 13.8 | 155 ± 56.7 | 68.6 ± 35.1 | 91.6 ± 23.9 | 75.7 ± 34.3 |
| 85 | trans-7-Hexadecene | alkene | 43.781 | - | 83.7 ± 30.9 | 44.2 ± 28.7 | 14.4 ± 5.98 | 37.7 ± 16.7 |
| 87 | Ylangene | sesquiterpene | 43.988 | 10,572 ± 1341 | 96.2 ± 32.1 | 813 ± 345 | 39 ± 20.6 | - |
| 88 | (−)-Aristolene | sesquiterpene | 44.106 | 15,240 ± 2280 | 944 ± 251 | 13,881 ± 6678 | 154 ± 41.4 | 27.3 ± 8.56 |
| 89 | β-Elemene | sesquiterpene | 44.253 | 266 ± 64.2 | 2770 ± 1203 | 18,305 ± 8626 | 15.3 ± 5.53 | 46.5 ± 20.4 |
| 90 | Isoledene | sesquiterpene | 44.383 | - | 74 ± 10.1 | 422 ± 318 | 8.37 ± 6.73 | 6.03 ± 2.1 |
| 92 | β-Humulene | sesquiterpene | 44.502 | - | 89.9 ± 34.4 | 366 ± 194 | 8.16 ± 2.46 | - |
| 95 | α-Farnesene | sesquiterpene | 44.690 | 4354 ± 846 | 63.1 ± 26.8 | 317 ± 208 | - | 7.95 ± 3.88 |
| 96 | α-Gurjunene | sesquiterpene | 44.778 | 647 ± 138 | 187 ± 79.1 | 5032 ± 2941 | 18.3 ± 8.73 | - |
| 97 | β-Cubebene | sesquiterpene | 44.871 | 2067 ± 470 | 61.2 ± 18.4 | 942 ± 514 | - | 5.61 ± 3.25 |
| 101 | Bicyclo[4.4.0]dec-1-ene, 2-isopropyl-5-methyl-9-methylene- | sesquiterpene | 45.110 | 3410 ± 679 | - | 441 ± 269 | 16.7 ± 3.74 | 3.42 ± 1.65 |
| 104 | Valencene | sesquiterpene | 45.511 | 1694 ± 458 | 425 ± 215 | 2675 ± 2334 | 52.3 ± 18.1 | 1.74 ± 0.71 |
| 105 | α-Selinene | sesquiterpene | 45.590 | 10,481 ± 2747 | 536 ± 115 | 3143 ± 2099 | 30 ± 20.2 | - |
| 106 | α-Farnesene | sesquiterpene | 45.742 | 1662 ± 320 | 1079 ± 553 | 13,975 ± 9284 | - | 3.58 ± 1.06 |
| 109 | Cubenene | sesquiterpene | 46.030 | 2869 ± 817 | 504 ± 235 | 9414 ± 6261 | - | 4.51 ± 1.6 |
| 111 | δ-Guaiene | sesquiterpene | 46.181 | 9186 ± 5597 | 100 ± 39.7 | 1255 ± 890 | - | 1.46 ± 0.44 |
| 113 | δ-Cadinene | sesquiterpene | 46.329 | 1503 ± 177 | 3196 ± 1441 | 11,706 ± 8150 | - | 6.42 ± 1.85 |
| 114 | β-Germacrene | sesquiterpene | 46.421 | 606 ± 126 | 335 ± 117 | 2428 ± 1666 | 15.2 ± 5.42 | - |
| 115 | β-Panasinsene | sesquiterpene | 46.489 | 1508 ± 382 | 161 ± 65.9 | 1218 ± 853 | 21.8 ± 3.71 | - |
| 116 | β-Cadinene | sesquiterpene | 46.616 | 164 ± 39.9 | 307 ± 178 | 4222 ± 3500 | 9.23 ± 3.28 | - |
| 122 | Cadina-1(10),6,8-triene | alkene | 47.141 | 60.7 ± 16.9 | 34.9 ± 14 | 299 ± 145 | 12.3 ± 8.27 | - |
| 130 | Germacrene D | sesquiterpene | 47.993 | 52.8 ± 19.9 | 37.8 ± 15.4 | 438 ± 298 | 10.7 ± 4.56 | - |
| 131 | α-Cadinol | alcohol | 48.145 | 272 ± 67 | 23.3 ± 6.58 | 370 ± 256 | - | 2.86 ± 2.72 |
| 132 | Naphthalene, 1,6-dimethyl-4-(1-methylethyl)- | aromatic hydrocarbons | 48.345 | - | 68.7 ± 22.8 | 603 ± 65 | - | 13.85 ± 5 |
| No. | Compound Name | Peak Area (×107) a | p-Value b | |||
|---|---|---|---|---|---|---|
| High | Medium | Low | Toothpick | |||
| 1 | Ethanol | 2.36 ± 0.9 | 2.62 ± 0.2 | 1.56 ± 0.41 | 1.58 ± 0.37 | 0.791 |
| 2 | 1,4-Pentadiene | 6.99 ± 2.39 | 7.98 ± 3.29 | 8.89 ± 2.49 | 0.91 ± 0.13 | 0.010 |
| 3 | 2-methylfuran | 4.4 ± 1.13 | 5.25 ± 0.9 | 4.8 ± 1.24 | 2.26 ± 0.52 | 0.009 |
| 4 | 1-Propanol, 2-methyl- | 5.38 ± 1.09 | 6.02 ± 0.9 | 5.16 ± 1.21 | 9.17 ± 3.09 | 0.045 |
| 5 | 3-methylbutanol | 1.83 ± 0.63 | 2.35 ± 0.56 | 1.93 ± 0.45 | 2.7 ± 1.02 | 0.061 |
| 6 | 2-methylbutanol | 2.07 ± 0.44 | 2.61 ± 0.52 | 2.34 ± 0.51 | 3.12 ± 1.03 | 0.012 |
| 7 | Toluene | 0.66 ± 0.16 | 0.66 ± 0.11 | 0.63 ± 0.07 | 0.84 ± 0.16 | 0.005 |
| 8 | (−)-Aristolene | 19.27 ± 10.45 | 21.66 ± 6.07 | 10.33 ± 1.63 | 15.04 ± 8.68 | 0.597 |
| 9 | β-Elemene | 66.06 ± 35.61 | 86.67 ± 36.71 | 37.35 ± 5.26 | 71.08 ± 41.45 | 0.381 |
| 10 | α-Farnesene | 23.52 ± 18.37 | 31.07 ± 14.74 | 9.61 ± 3.76 | 18.99 ± 10.72 | 0.499 |
| 11 | Cubenene | 19.48 ± 13.56 | 28.81 ± 13.4 | 7.86 ± 3.78 | 27.33 ± 14.48 | 0.363 |
| 12 | δ-Cadinene | 50.58 ± 30.48 | 53.46 ± 11.63 | 26.83 ± 6.6 | 51.12 ± 22.88 | 0.712 |
| 13 | β-Germacrene | 6.22 ± 4.88 | 7.24 ± 2.67 | 2.92 ± 0.83 | 5.12 ± 2.25 | 0.174 |
| 14 | β-Panasinsene | 3.45 ± 3.22 | 2.77 ± 0.55 | 1.46 ± 0.4 | 2.85 ± 1.06 | 0.402 |
| 15 | β-Cadinene | 5.94 ± 4.84 | 5.44 ± 1.18 | 2.65 ± 0.75 | 5.7 ± 2.66 | 0.613 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, D.; She, J.; Gower, J.L.; Stokes, C.E.; Windham, G.L.; Baird, R.E.; Mlsna, T.E. Effects of Growth Parameters on the Analysis of Aspergillus flavus Volatile Metabolites. Separations 2016, 3, 13. https://doi.org/10.3390/separations3020013
Sun D, She J, Gower JL, Stokes CE, Windham GL, Baird RE, Mlsna TE. Effects of Growth Parameters on the Analysis of Aspergillus flavus Volatile Metabolites. Separations. 2016; 3(2):13. https://doi.org/10.3390/separations3020013
Chicago/Turabian StyleSun, Dongdi, Jinyan She, Julie L. Gower, C. Elizabeth Stokes, Gary L. Windham, Richard E. Baird, and Todd E. Mlsna. 2016. "Effects of Growth Parameters on the Analysis of Aspergillus flavus Volatile Metabolites" Separations 3, no. 2: 13. https://doi.org/10.3390/separations3020013
APA StyleSun, D., She, J., Gower, J. L., Stokes, C. E., Windham, G. L., Baird, R. E., & Mlsna, T. E. (2016). Effects of Growth Parameters on the Analysis of Aspergillus flavus Volatile Metabolites. Separations, 3(2), 13. https://doi.org/10.3390/separations3020013