1. Introduction
Assuming that during the process of evolution, more and more complex organisms, characterized by an increased awareness of life, emerge gradually, then, in accordance with Charles Darwin’s conclusion, as a result of evolution, “innumerable transitional forms must have existed” [
1]. Surprisingly, Charles Darwin questioned his theory of evolution by asking a rhetorical question: “why do we not find them (i.e., transitional forms) imbedded in countless numbers in the crust of the earth?” [
1]. Despite the large differences between multicellular organisms, there is an absence (or scarcity) of intermediate transitional forms, i.e., individual organisms appear in the fossil record suddenly without evidence of intermediate transitional forms [
2]. Moreover, every appearance of more complex organisms is discontinuous and unpredictable [
3]. Based on the evidence of absence (or scarcity) of transitional fossils in the fossil record, a new punctuated equilibrium theory has been formulated [
4,
5]. According to this theory, most changes occur suddenly during the formation of the species [
4,
5,
6]. After an abrupt appearance of a species in the fossil record, the population undergoes stabilization (showing slight evolutionary changes throughout most of its geological history) [
4]. The state of slight (or lack of) evolutionary morphological changes has been termed ‘stasis’. The occurrence of long stasis states may be the reason why some researchers are of the opinion that so far, modern science has not provided a satisfactory empirical explanation for the increasing complexity of living organisms throughout evolution [
7]. It appears that the evolution of organisms occurs mostly as microevolution in separated genome attractors (i.e., organism-kind genome attractors) that ensure stable evolutionary changes of organisms [
8]. The term ‘attractor’ means a system configuration toward which the system strives over time. Once the attractor is attained, the system is stable enough to return to its original state when eventual perturbations disappear [
9]. Genome attractors are places where adaptation to the environment occurs through natural selection, as a result of which organisms become better adapted to the environment while being stabilized. From this point of view, organism-kind genome attractors (i.e., genome attractors of organism-kind) can be considered as genome attractors that allow the stabilization of configurations of features that are typical for given organisms (for example, the favored configuration of features typical for humans is stabilized during the entrapment of organisms in human-kind genome attractor). This new attempt is in accordance with the current interest of researchers for ‘organisms as attractors in phase space’ [
10]. Macroevolution can be caused by the occurrence of the genome instability, resulting in the organisms leaving current genome attractors and attaining new genome attractors. Stable evolution in genome attractors (as long periods of microevolution) and a change of genome attractors (as relatively short periods of macroevolution) may indicate the discontinuous nature of the evolution process [
8]. Genome attractors, due to allowing the stable evolution of organisms, can be termed ‘oases of life’. Existence of the instability area between genome attractors results in big evolutionary distances between organisms that are trapped in different genome attractors and may also explain the lack (or a very small number) of intermediate transitional forms.
This article can be considered as a continuation of previously published articles in which unified cell bioenergetics (UCB) and new attempts to establish methods that can be used to examine the evolution of organisms (including the evolution of transformed cells) have been presented [
8,
11,
12,
13,
14,
15]. These new methods include the use of artificial neural network (ANN) and a semihomologous approach to recognize evolution. In the previously published articles, the neural network has been trained using cytochrome b sequences that have been used as phylogenetic probes that represent whole organisms [
8,
11]. It has been presented that the taught neural network is able to recognize the evolution of unknown (i.e., unseen before) organisms. The analysis of the results has led to the conclusion that during evolution, organisms are trapped in local organism-kind genome attractors.
In view of the presented information, the main aim of this article has been formulated as a presentation of new research in order to confirm the discontinuous character of evolution. The variability of selected groups of organisms has been checked using a semihomologous approach in order to examine the influence of evolution on the stability of organisms trapped in organism-kind genome attractors. Moreover, it has been pointed out that in accordance with unified cell bioenergetics (UCB), disturbances (perturbations) of cell bioenergetics can be considered as a main driver of evolution.
In this article, the evolution of organisms is recognized using an artificial neural network, but unlike the previous articles, cytochrome c sequences have been used to train and then to recognize the evolution of organisms. Cytochrome c is an omnipresent and essential protein that is highly conserved across the spectrum of organisms. As estimated on the basis of fossil evidence, the number of amino acid differences in cytochrome c between different lineages varies linearly in time [
16,
17]. These features along with the small size of cytochrome c (molecular weight about 12 kDa) make it suitable for use to study cladistics [
18].
Unified cell bioenergetics allows the universal interpretation of several main cell bioenergetic effects (inter alia, the Pasteur, Crabtree, Kluyver, glucose effects) that can occur during bioprocesses and certain diseases (diabetes, cancer, heart diseases) based on the intramitochondrial NADH (mtNADH) level [
19,
20,
21,
22,
23,
24,
25]. Mitochondria control both the life and death of cells [
19,
26]. Moreover, it is known that in almost every disease, an increased lactate concentration is caused by a shift in mitochondrial activity [
27,
28]. In addition, in accordance with UCB, a common feature of some diseases is an increased level of mtNADH that leads to an increase in the lactate production rate (LPR) [
19,
21]. It is known that the reactive oxygen species (ROS) formation rate increases exponentially with NADH concentration when most electron donors are in a non-reducing state [
29]. In accordance with existing theories, organism evolution driving forces include random genetic mutations and natural selection [
30,
31]. A moderate ROS level may affect a number of cell biological processes through transcriptional regulation, but a high ROS level can result in severe oxidative damage to DNA, proteins, and biolipid membranes [
19,
32,
33,
34]. Viewed in this light, a moderate ROS level can stimulate organism evolution without a change of genome attractors, but bioenergetic problems (which lead to, among others, a higher level of mtNADH and an increase in LPR) can cause the occurrence of specific diseases (including cancer) and also can lead to changes of genome attractors as a result of high ROS formation rate [
8,
19]. From this point of view, ROS can act as a factor stimulating organism evolution also by random genetic mutations.
This article is organized as follows: firstly, the methods and theoretical bases are listed, including a description of the neural network implementation, the basis of unified cell bioenergetics, and the semihomologous approach. Secondly, selected aspects of the evolution of organisms are examined and discussed. Finally, the research conclusions are presented.
3. Results and Discussion
The “is evolution a continuous or discontinuous process?” question has intrigued scientists for many years [
40]. This question was fascinating in Charles Darwin’s time, who argued that during the process of evolution, more and more complex organisms emerge gradually (see Introduction), and this question remains still relevant nowadays, when the increasing computational power of computers gives new opportunities to search for an answer to this question. Different computational methods for generating phylogenetic trees can be used to support the search for this answer. The exemplary phylogenetic tree construction methods include Neighbor Joining, Maximum Parsimony, Maximum Likelihood, and Bayesian Inference methods [
41,
42,
43]. The number of phylogenetic trees that can be created depends mainly on the number of analyzed organisms, and it grows rapidly while increasing the number of organisms (for example, for 50 organisms, the number of possible rooted trees is bigger than the number of atoms in the universe) [
41]. Since it is impossible to analyze each of the possible trees for a large number of organisms, for this reason, it is also impossible to uncover the real truth about evolution when the number of organisms is high. In the future, the use of quantum computers, which will provide much more computing power and storage capacity compared to today’s digital computers, should bring an understanding of some evolution aspects unknown today, giving the possibility of analyzing a much larger number of phylogenetic trees. Looking for a solution to the question (“is evolution a continuous or discontinuous process?”) and still waiting for the evolution of quantum computers technology, in this article, the evolution of organisms is examined not by using tree-generating computational methods but by one of the artificial intelligence methods, i.e., artificial neural networks. The use of artificial neural networks allows the evolution of organisms to be examined through recognition (contrary to computational methods for generating phylogenetic trees). In order to gain recognition ability, artificial neural networks must be taught using a set of patterns. In this study, before teaching the neural networks, the organisms (that cytochrome c sequences have been used to teach the three and four-layer neural networks) have been ordered from the most primitive organisms to the most developed organisms. The preliminary order of the organisms has been established by calculating the evolutionary distances between each of the organisms used to teach the neural networks and
Homo sapiens (#20). Evolutionary distances (with Poisson correction and the selected pairwise deletion option) have been calculated as a distance matrix using the MEGA-X program [
44]. Additionally, the semihomologous approach has been used to check variability of homologous (“R”), semihomologous (i.e., sum of the “#” and “
$” (i.e., “# +
$”)), and “-” positions. Before using the dotPicker program (with an implemented semihomologous approach), each sequence of cytochrome c has been aligned with a
Homo sapiens cytochrome c sequence using the ClustalW program (implemented as the option in the MEGA-X program), and then, a pairwise deletion algorithm has been executed. The semihomologous approach allowed establishing the final order of the organisms used to teach ANN. This was important because after the calculations, it turned out that for some organisms, the evolutionary distances were the same, i.e., for Fly (#9) and Spider (#10) (evolutionary distance equal to 0.2499417445), for Horse (#14), Cat (#15), and Dog (#16) (0.1106655679), and for Gray whale (#17) and Domestic sheep (#18) (0.1000834586) (see
Table A1 in
Appendix A). In the semihomologous approach, the organism that has higher number of homologous positions is closer to
Homo sapiens (in the considered case). If the number of homologous positions is the same, then the organism that has the higher number of semihomologous positions is closer to
Homo sapiens. Using the semihomologous approach, it was possible to establish the order of the organisms in an unambiguous way. The final order of the organisms, i.e., the established order of the organisms from the most primitive (characterized by the biggest evolutionary distance in relation to
Homo sapiens) to the most developed organism in relation to
Homo sapiens (characterized by the smallest evolutionary distance in relation to
Homo sapiens) is presented in
Table 1. The results of calculations (i.e., evolutionary distances with Poisson correction between
Homo sapiens and organisms used to teach the three and four-layer neural networks, the number of homologous positions (i.e., “R” positions in [%]), semihomologous positions (i.e., sum of “#” and “
$” positions in [%]), and the other positions (i.e., “-” positions in [%])) are presented in
Table A1 (see
Appendix A) and in
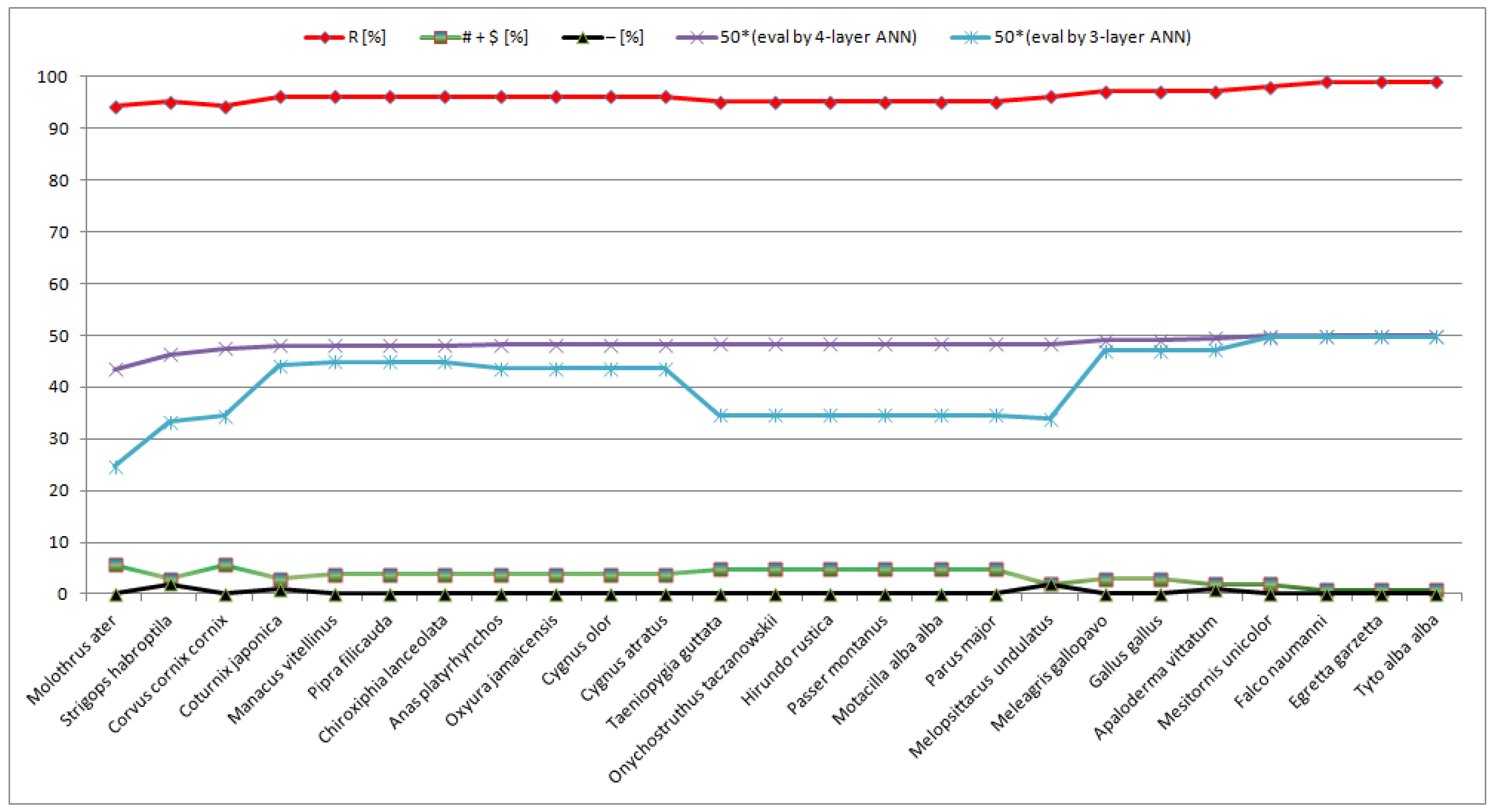
Figure 1.
An important and interesting observation is that decreasing evolutionary distances in relation to
Homo sapiens is associated with increasing the number of homologous positions (i.e., “R” positions) and decreasing both the number of semihomologous positions (i.e., “# +
$” positions) and the other positions (i.e., “-” positions) (
Figure 1). It can be concluded that during decreasing evolutionary distances, positions with two or three point mutations in the codons of compared amino acids (i.e., “-” positions) undergo changes to semihomologous positions (i.e., positions with one-point mutation in the codons of compared amino acids), and semihomologous positions undergo changes to homologous positions.
In the next step of the work, five groups of organisms have been examined: hominoids, Old World Monkeys, New World Monkeys, birds, and fish. Cytochrome c sequences of the selected organisms from these groups have been recognized using the taught three and four-layer neural networks. Additionally, the semihomologous approach has been applied to a deeper examination of the genetic characteristics of organisms belonging to these groups.
Selected representatives of hominoids, Old World Monkeys and New World Monkeys, have been examined in relation to
Homo sapiens. The results of calculations (i.e., recognized evolutionary similarities between
Homo sapiens and the selected representatives of hominoids, Old World Monkeys, and New World Monkeys by the three and four-layer neural networks and the number of homologous positions (i.e., “R” positions in [%]), semihomologous positions (i.e., sum of “#” and “
$” positions in [%]) and the other positions (i.e., “-” positions in [%])) are presented in
Table A2 (see
Appendix A) and in
Figure 2.
The organisms presented in
Figure 2 and in
Table A2 (see
Appendix A) have been set in the order of increasing evaluation by the four-layer neural network, i.e., in the order of increasing evolutionary similarities in relation to
Homo sapiens. From
Figure 2 and the results presented in
Table A2 (see
Appendix A), it is visible that the use of the neural networks (both the three and four-layer networks) allows the separation of New World Monkeys (for this group of organisms, the recognized by the four-layer neural network evolutionary similarity to
Homo sapiens is in the range [0.94334, 0.99716]; see
Table A2) from the other organisms (i.e., Old World Monkeys (for which the recognized evolutionary similarity to
Homo sapiens is in the range [0.99784, 0.99813]) and hominoids (for which the recognized evolutionary similarity to
Homo sapiens is in the range [0.99895, 0.99924]). It should be noted that the maximum evolutionary similarity recognized by the neural networks is equal to 1 and the minimum evolutionary similarity is equal to 0. The range of recognized evolutionary similarities (i.e., [0,1]) results from the specificity of the neural networks teaching (see
Section 2.1). Based on the previous published works, the use of sequences of cytochrome b allows also a more clear separation Old World Monkeys from hominoids [
8,
11]. It was possible due to the greater length of cytochrome b sequences (comparing to sequences of cytochrome c), to more clearly recognize the pattern of evolution. The separation of New World Monkeys from Old World Monkeys and Old World Monkeys from hominoids may indicate that the evolution of these groups of organisms takes place in different genome attractors. Additionally, to check this conclusion, a semihomologous approach has been applied. In this case, the use of the semihomologous approach also indicates a clear separation of NWM from the other organisms (i.e., OWM and hominoids) (
Table 3).
Selected representatives of birds have been checked in relation to Eagle (#13,
Aquila chrysaetos chrysaetos). The results of calculations (i.e., recognized evolutionary similarities between
Aquila chrysaetos chrysaetos and the selected representatives of birds by the three and four-layer neural networks and the number of homologous positions (i.e., “R” positions in [%]), semihomologous positions (i.e., sum of “#” and “
$” positions in [%]), and the other positions (i.e., “-” positions in [%])) are presented in
Table A3 (see
Appendix A) and in
Figure 3.
The organisms presented in
Figure 3 and in
Table A3 (see
Appendix A) have been set in the order of increasing evaluation by the four-layer neural network, i.e., in the order of increasing evolutionary similarities in relation to
Aquila chrysaetos chrysaetos (#13). From
Figure 3 and the results presented in
Table A3 (see
Appendix A), it is visible that the use of the neural networks (both the three and four-layer networks) allows the recognition of the bird-kind genome attractor (characterized by high average evaluations (i.e., high average evolutionary similarities) by the three and four-layer neural networks equal to 0.82162 and 0.96594, respectively). Additionally, to check this conclusion, the semihomologous approach has been applied. In this case, the use of the semihomologous approach also indicates the separation of birds (i.e., the big average number of “R” positions equal to 96.30 [%]) from the other organisms. Based on the results (
Figure 3 and
Table A3), a conclusion can be drawn that is presented as Remark 1.
Remark 1. During evolution in this organism-kind genome attractor (indicated by increasing evolutionary similarities recognized by the four-layer neural network), the number of homologous positions (i.e., “R” positions) increases and the number of semihomologous positions (i.e., “# + $” positions) decreases, while the number of “-” positions remains small. It can be concluded that during evolution, semihomologous positions undergo changes to homologous positions, gradually increasing the number of homologous positions.
Selected representatives of fish have been checked in relation to Cod (#11,
Gadus morhua). The results of calculations (i.e., recognized evolutionary similarities between
Gadus morhua and the selected representatives of fishes by the three and four-layer neural networks and the number of homologous positions (i.e., “R” positions in [%]), semihomologous positions (i.e., sum of “#” and “
$” positions in [%]), and the other positions (i.e., “-” positions in [%])) are presented in
Table A4 (see
Appendix A) and in
Figure 4.
The organisms presented in
Figure 4 and in
Table A4 (see
Appendix A) have been set in the order of increasing evaluation by the four-layer neural network, i.e., in the order of increasing evolutionary similarities in relation to
Gadus morhua (#11). From
Figure 4 and the results presented in
Table A4 (see
Appendix A), it is visible that the use of the neural networks (both the three and four-layer networks) allows recognition of the fish-kind genome attractor (characterized by high average evaluations (i.e., high average evolutionary similarities) by the three and four-layer neural networks equal to 0.85739 and 0.92734, respectively). Additionally, to check this conclusion, the semihomologous approach has been applied. In this case, the use of the semihomologous approach also indicates the separation of fishes (i.e., the big average number of “R” positions equal to 93.04 [%]) from the other organisms. Oscillations of the number of “R”, “# +
$”, and “-” positions can indicate a bigger variability of fishes in the fish-kind genome attractor, but the conclusion presented as Remark 1 is also valid in this case.
Standard deviation applied to the number of homologous (i.e., “R”), semihomologous (i.e., sum of “#” and “
$” (i.e., “# +
$”)), and “-” positions in the semihomologous approach has been calculated in order to check the variability of the selected organisms in their organism-kind genome attractors (
Table 4).
The higher standard deviations obtained for fish indicate that the numbers of “R”, “# +
$”, and “-” positions are spread out over a wider range (among the checked organisms) for these organisms (
Table 4). This can also indicate that for the checked organisms, the variability of fishes in the fish-kind genome attractor is the highest. The evolutionary relationships recognized by the artificial neural networks are also characterized by the largest differences between the results obtained using the three and four-layer networks for fish. The variability of birds in the bird-kind genome attractor is smaller than the variability of fishes in the fish-kind genome attractor. Taking into account standard deviations of the number of “R” positions, the variability of hominoids in the hominoid-kind genome attractor is the smallest (
Table 4). Based on these results, it can be concluded that more developed organisms show greater persistence (stability) in their organism-kind genome attractors. Since the real challenge is to find unambiguous intermediate transitional forms, for this reason, it can be concluded that organisms have been trapped in their organism-kind genome attractors eons ago. Being trapped in genome attractors, organisms undergo perturbations that drive microevolution. Looking at the multi-level organization of nature, most biologists are typically thinking that cause and effect events come from a lower (molecular) level of hierarchy of organization [
45]. These events, in the form of perturbations, travel up this hierarchy [
45]. Taking into account this point of view and in accordance with unified cell bioenergetics (UCB), the effects of bioenergetic disturbances (perturbations) can be depicted as shown in
Figure 5.
In accordance with UCB, small disturbances (at molecular level) of cell bioenergetics can lead to small disturbances related, among others, to the Pasteur, Crabtree, Kluyver, and glucose effects; i.e., small disturbances can cause reversible effects (see
Figure 5a). For example, these disturbances can be caused by a moderate ROS level that may affect a number of cell biological processes through transcriptional regulation (see Introduction). That means that small disturbances of cell bioenergetics can stimulate organism evolution without a change of genome attractor (i.e., can stimulate microevolution). However, huge or long-lasting (for example age-related) disturbances of cell bioenergetics can lead to specific diseases (including Alzheimer’s disease and cancer transformation) (see
Figure 5b) [
21,
28]. The propagation of disturbances caused by a high level of ROS (resulting in severe oxidative damage to DNA, see Introduction) can also lead to changes of genome attractors by transformed cells [
8].




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




