
Harnessing the Potential of Symbiotic Endophytic Fungi and Plant Growth-Promoting Rhizobacteria to Enhance Soil Quality in Saline Soils

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Isolation of Bacteria
2.2. Purification of Bacteria
2.3. Assessment of Some PGPR Properties of the Isolates
2.3.1. Qualitative Assessment of Insoluble Phosphate Solubilization Potential in NBRIP Medium
2.3.2. Quantitative Assessment of Insoluble Phosphate Solubilization Potential in Broth NBRIP Medium
2.3.3. Qualitative Assessment of Mica Solubilization Potential
2.3.4. Auxin Production
2.3.5. Siderophore Production
2.3.6. Salinity Assay
2.4. Selection of Superior Rhizobacteria for Phenotypic and Molecular Identification
2.4.1. Phenotypic Identification of the Selected Isolates
2.4.2. Molecular Identification and Phylogenetic Tree
2.5. Pot Experiment
2.5.1. Plant Harvest and Chemical Analyses
2.5.2. Statistical Analysis
3. Results and Discussion
3.1. In-Vitro Trials
3.1.1. PGPR-Specific Tests
3.1.2. Phenotype Identification
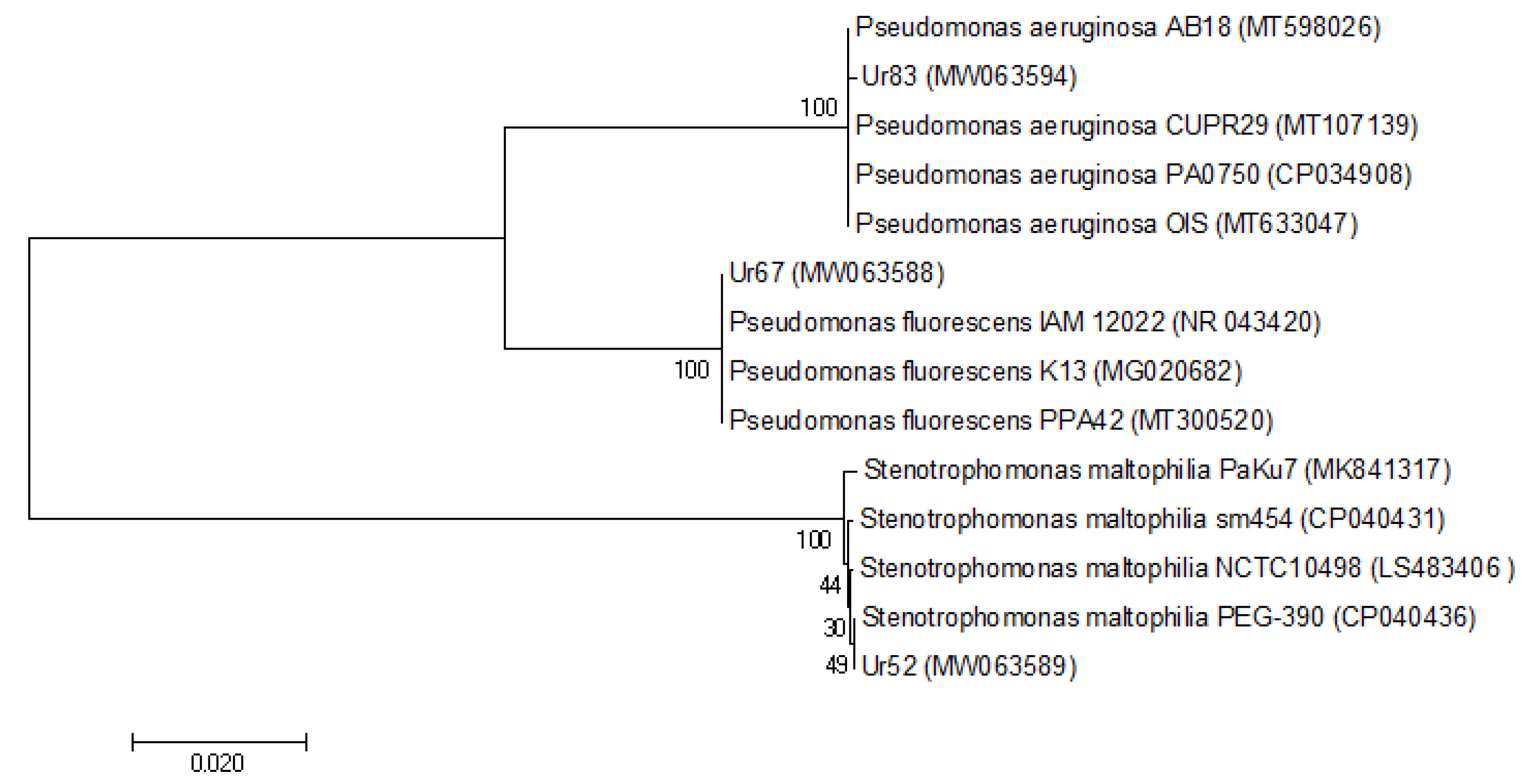
3.1.3. Molecular and Genotypic Identification of the Superior Isolates
3.2. Greenhouse Trials
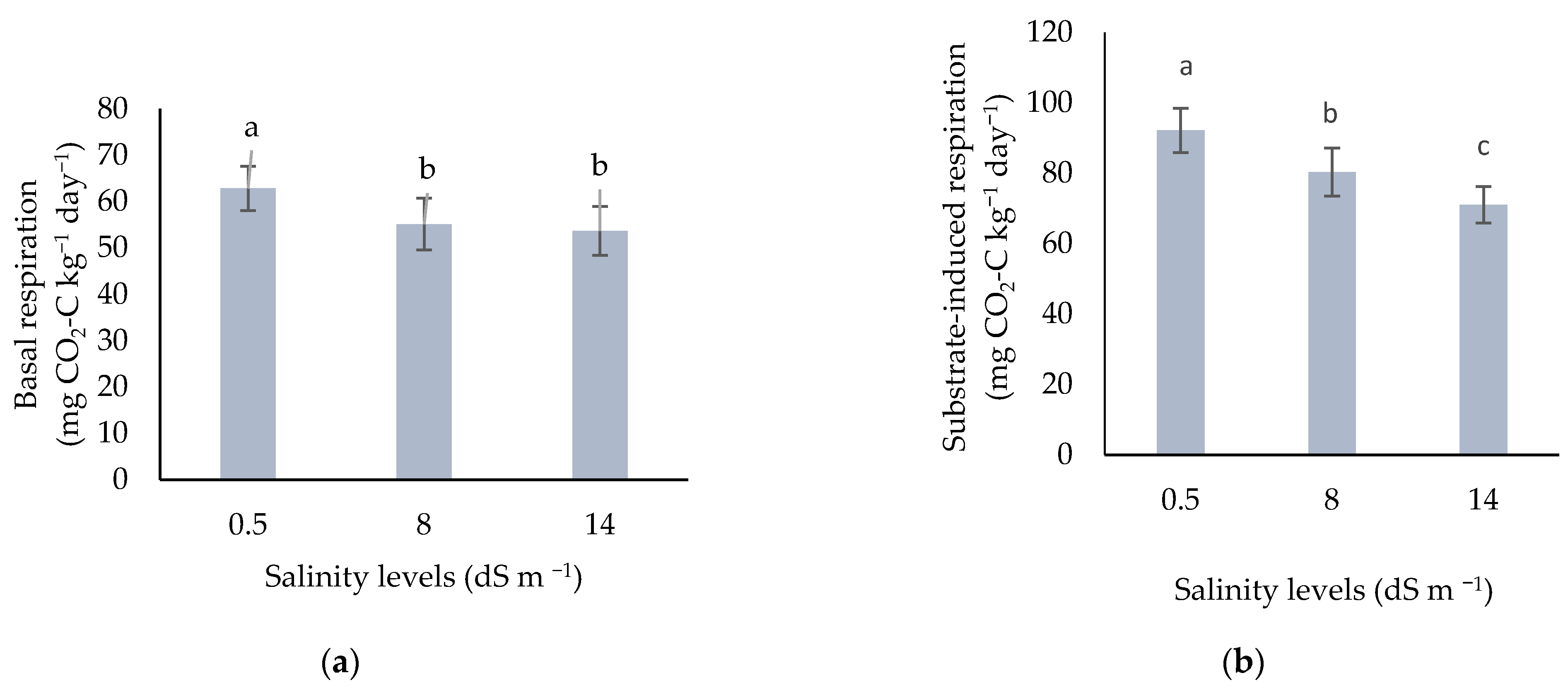
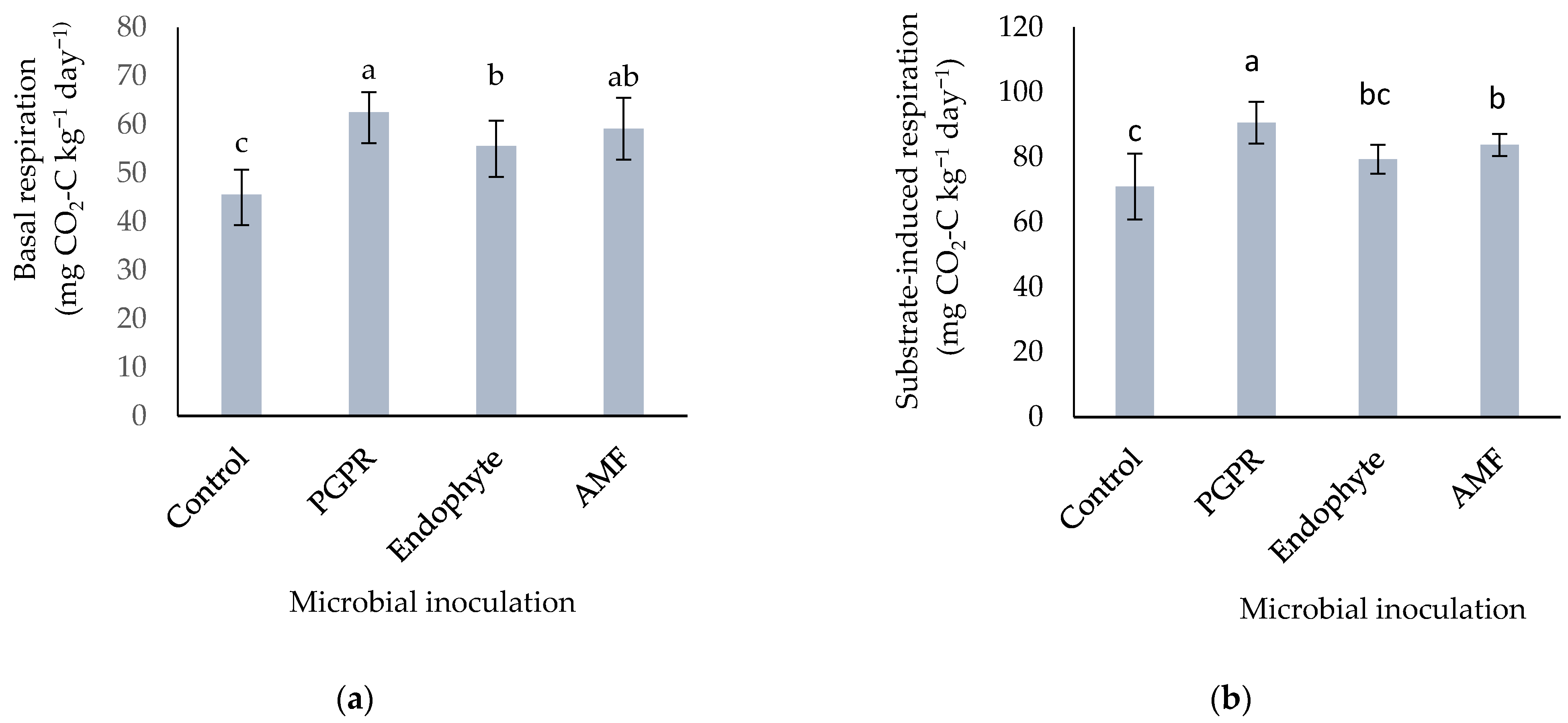
3.2.1. Basal and Substrate-Induced Respiration
3.2.2. Microbial Biomass Carbon
3.2.3. Metabolic Quotient and Carbon Availability
3.2.4. Acid and Alkaline Phosphatase Enzyme
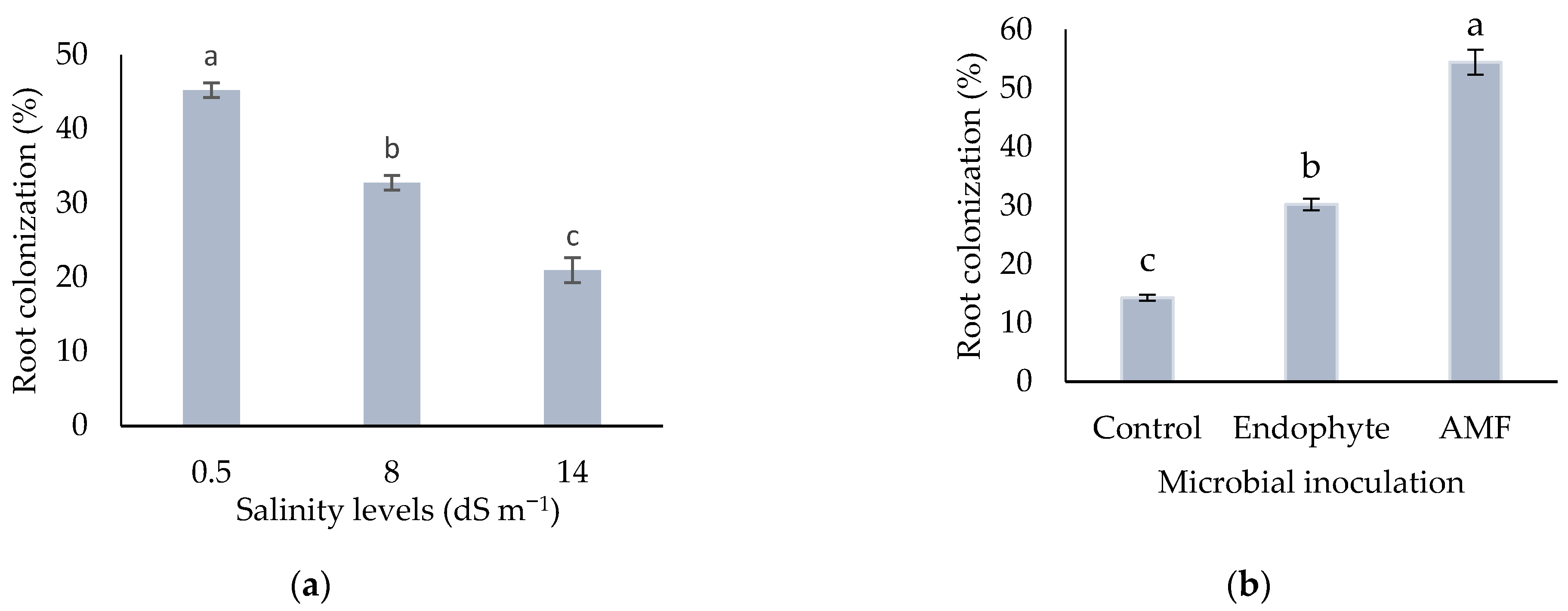
3.2.5. Root Colonization Percentage
3.2.6. Shoot Dry Weight
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Karaki, G.N. Nursery inoculation of tomato with arbuscular mycorrhizal fungi and subsequent performance under irrigation with saline water. Sci. Hortic. 2006, 109, 1–7. [Google Scholar] [CrossRef]
- Barin, M.; Aliasgharzad, N.; Olsson, P.A.; Sadeghiani, M.H.R.; Moghddam, M. The abundance of arbuscular mycorrhizal fungi in relation to soil salinity around Lake Urmia in northern Iran. Pedobiologia 2013, 56, 225–232. [Google Scholar] [CrossRef]
- Esmaili, E.; Kapourchal, S.A.; Malakouti, M.J.; Homaee, M. Interactive effect of salinity and two nitrogen fertilizers on growth and composition of sorghum. Plant Soil Environ. 2008, 54, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Barin, M.; Aliasgharzad, N.; Olsson, P.A.; Rasouli-Sadaghiani, M.H. Salinity-induced differences in soil microbial communities around the hypersaline Lake Urmia. Soil Res. 2015, 53, 494–504. [Google Scholar] [CrossRef]
- Hammer, E.C.; Nasr, H.; Pallon, J.; Olsson, P.A.; Wallander, H. Elemental composition of arbuscular mycorrhizal fungi at high salinity. Mycorrhiza 2011, 21, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Rietz, D.N.; Haynes, R.J. Effects of irrigation-induced salinity and sodicity on soil microbial activity. Soil Biol. Biochem. 2003, 35, 845–854. [Google Scholar] [CrossRef]
- Yang, R.; Qin, Z.; Wang, J.; Xu, S.; Zhao, W.; Zhang, X.; Huang, Z. Salinity changes root occupancy by arbuscular mycorrhizal fungal species. Pedobiologia 2020, 81, 150665. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Kucharski, M.; Kucharski, J. Applicability of biochemical indices to quality assessmnet of soil pulluted with heavy metals. J. Elem. 2013, 18, 723–732. [Google Scholar]
- Brendecke, J.W.; Axelson, R.D.; Ian, L.; Pepper, I.L. Soil microbial activity as an indicator of soil fertility—Long term effects of municipal sewage-sludge on an arid soil. Soil Biol. Biochem. 1993, 25, 751–758. [Google Scholar] [CrossRef]
- Zarei, M.J.; Hajinia, S.; Karimi, N.; Goltapeh, E.M.; Rejali, F.; Varma, A. Effect of Piriformospora indica and Azospirillum strains from saline or non-saline soil on mitigation of the effects of NaCl. Soil Biol. Biochem. 2012, 45, 139–146. [Google Scholar] [CrossRef]
- Stefanoski, D.C.; Figueiredo, C.C.D.; Santos, G.G.; Marchão, R.L. Selecting soil quality indicators for different soil management systems in the Brazilian Cerrado. Pesqui. Agropecu. Bras. 2016, 51, 1643–1651. [Google Scholar] [CrossRef] [Green Version]
- Paz-Ferreiro, J.; Fu, S. Biological Indices for Soil Quality Evaluation: Perspectives and Limitations. Land Degrad. Dev. 2013, 27, 14–25. [Google Scholar] [CrossRef]
- MomoWobeng, N.B.; Banfield, C.C.; Megueni, C.; MarieMapongmetsem, P.; Dippold, M.A. Impact of legumes on soil microbial activity and C cycle functions in two contrasting Cameroonian agroecological zones. Pedobiologia 2020, 81, 150662. [Google Scholar]
- Bradford, M.A.; McCulley, R.L.; Crowther, T.W.; Oldfield, E.E.; Wood, S.A.; Fierer, N. Cross-biome patterns in soil microbial respiration predictable from evolutionary theory on thermal adaptation. Nat. Ecol. Evol. 2019, 3, 223–231. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, X. Soil Respiration and the Environment; Academic Press: San Diego, CA, USA, 2006; p. 328. [Google Scholar]
- Lin, Y.J.; Gao, F.; Zhang, J.L.; Zhou, L.Y.; Zhang, X.M.; Li, B.L.; Zhao, H.J.; Li, X.D. Soil microbial biomass and respiration rate under effects of different planting patterns of peanut. Ying Yong Sheng Tai Xue Bao 2010, 21, 2323–2328. [Google Scholar]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leiros, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Bertini, S.C.B.; Azevedo, L.C.B.; de Carvalho Mendes, I.; Cardoso, E.J.B.N. Hierarchical partitioning for selection of microbial and chemical indicators of soil quality. Pedobiologia 2014, 57, 293–301. [Google Scholar] [CrossRef]
- Rao, D.L.N.; Pathak, H. Ameliorative influence of organic matter on biological activity of salt-affected soils. Arid. Land Res. Manag. 1996, 10, 311–319. [Google Scholar] [CrossRef]
- Batra, L.; Manna, M.C. Dehydrogenase activity and microbial biomass carbon in salt-affected soils of semiarid and arid regions. Arid. Land Res. Manag. 1997, 11, 295–303. [Google Scholar] [CrossRef]
- Tripathi, S.; Kumari, S.; Chakraborty, A.; Gupta, A.; Chakrabarti, K.; Bandyapadhyay, B.K. Microbial biomass and its activities in salt-affected coastal soils. Biol. Fertil. Soils 2006, 42, 273–277. [Google Scholar] [CrossRef]
- Wong, V.N.; Dalal, R.C.; Greene, R.S. Salinity and sodicity effects on respiration and microbial biomass of soil. Biol. Fertil. Soils 2008, 44, 943–953. [Google Scholar] [CrossRef]
- Yan, N.; Marschner, P. Response of soil respiration and microbial biomass to changing EC in saline soils. Soil Biol. Biochem. 2013, 65, 322–328. [Google Scholar] [CrossRef]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Kaur, G.; Reddy, M.S. Role of phosphate-solubilizing bacteria in improving the soil fertility and crop productivity in organic farming. Arch. Agron. Soil Sci. 2014, 60, 549–564. [Google Scholar] [CrossRef]
- Schaad, N.W.; Jones, J.B.; Chun, W. Laboratory Guide for the Identification of Plant Pathogenic Bacteria, 3rd ed.; American Phytopathological Society, APS Press: Saint Paul, MN, USA, 2001. [Google Scholar]
- Diep, C.N.; Hieu, T.N. Phosphate and potassium solubilizing bacteria from weathered materials of denatured rock mountain, Ha Tien, Kiên Giang province, Vietnam. Am. J. Life Sci. 2013, 1, 88–89. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Tallapragada, P.; Seshachala, U. Phosphate-solubilizing microbes and their occurrence in the rhizospheres of Piper betel in Karnataka, India. Turk. J. Biol. 2012, 36, 25–35. [Google Scholar]
- Cotteni, A. Soil and Plant Testing as a Basis of Fertilizer Recommendations; Food and Agriculture Organization of the United Nations: Rome, Italy, 1980; pp. 64–100. [Google Scholar]
- Hu, X.F.; Chen, J.; Guo, J.F. Two phosphate and potassium solubilizing bacteria isolated from Tiannu Mountain, Zhejiang, China. World J. Microbiol. Biotechnol. 2006, 22, 983–990. [Google Scholar] [CrossRef]
- Bent, E.; Tuzan, S.; Chanway, C.P.; Enebak, S. Alteration in plant growth and in root hormone levels of lodgepole pines inoculated with rhizobacteria. Can. J. Microbiol. 2001, 47, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Nakbanpote, W.; Panitlurtumpai, N.; Sangdee, A.; Sakulpone, N.; Sirisom, P.; Pimthong, A. Salt-tolerant and plant growth-promoting bacteria isolated from Zn/Cd contaminated soil: Identification and effect on rice under saline conditions. J. Plant Interact. 2014, 9, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Llop, P.; Caruso, P.; Cubero, J.; Morente, C.; López, M.M. A simple extraction procedure for efficient routine detection of pathogenic bacteria in plant material by polymerase chain reaction. J Microbiol. Methods 1999, 37, 23–31. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Yamasato, K. Phylogeny of spore-forming lactic acid bacteria based on 16S rRNA gene sequences. FEMS Microbiol. Lett. 1994, 115, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H. (Eds.) Methods of Soil Analysis, Part 3: Chemical Methods; John Wiley and Sons: Hoboken, NJ, USA, 2020; Volume 14. [Google Scholar]
- Hajiboland, R.; Aliasgharzadeh, N.; Farsad Laiegh, S.H.; Poschenieder, C.H. Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant Soil 2010, 331, 313–327. [Google Scholar] [CrossRef]
- Anderson, J.P.E.; Page, A.L.; Miller, R.H.; Keeney, D.R. (Eds.) Soil Respiration. In Methods of Soil Analysis: Microbiological and Chemical Properties, Part 2, 2nd ed.; SSSA: Madison, WI, USA, 1982; pp. 831–872. [Google Scholar]
- Nakamoto, T.; Wakahara, S. Development of substrate induced respiration (SIR) method combined with selective inhibition for estimating fungal and bacterial biomass in humic andosols. Plant Prod. Sci. 2004, 7, 70–76. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Ladd, J.N. Microbial biomass in soil measurement and turnover. Soil Biochem. 1981, 5, 415–471. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Martens, R. Methoden zur Quantitative Bestimmung und Charakterisierung der Mikrobiellen Biomasse in Böden. Eigenverlag des Institutes für Bodenbiologie der FAL Braunschweig 1991. Available online: https://www.openagrar.de/receive/timport_mods_00005888 (accessed on 7 October 2021).
- Liu, H.; Lv, W.; Wang, S.; Luo, C.; Zhang, Z.; Wang, Z.; Jiang, L.; Cui, X. Decreased soil substrate availability with incubation time weakens the response of microbial respiration to high temperature in an alpine meadow on the Tibetan Plateau. J. Soils Sediments 2019, 19, 255–262. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Brit. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Moira, E.K.; Henderson, M.E.K.; Duff, R.B. The release of metallic and silicates ions from minerals, rocks and soils by fungal activity. J. Soil Sci. 1963, 14, 237–245. [Google Scholar]
- Barin, M.; Sadeghi, S.; Rasouli-Sadaghiani, M.H.; Sepehr, E.; Dovlti, B.; Vahedi, R. Influence of K-solubilizing fungi on potassium release from silicate minerals and some growth indexes on corn (Zea mays L.). Appl. Soil Res. 2018, 3, 29–41. [Google Scholar]
- Hashemnejad, F.; Barin, M.; Khezri, M.; Ghoosta, Y.; Hammer, E.C. Isolation and identification of insoluble zinc-solubilising bacteria and evaluation of their ability to solubilise various zinc minerals. J. Soil Sci. Plant Nutr. 2021, 21, 1–9. [Google Scholar] [CrossRef]
- Wu, P.S.; Lu, C.Y.; Chang, L.Y.; Hsueh, P.R.; Lee, P.I.; Chen, J.M.; Lee, C.Y.; Chan, P.C.; Chang, P.Y.; Yang, T.T.; et al. Stenotrophomonas maltophilia bacteremia in pediatric patients—A 10-year analysis. J. Microbiol. Immunol. Infect. 2006, 39, 144–149. [Google Scholar]
- Deredjian, A.; Alliot, N.; Blanchard, L.; Brothier, E.; Anane, M.; Cambier, P.; Jolivet, C.; Khelil, M.N.; Nazaret, S.; Saby, N.; et al. Occurrence of Stenotrophomonas maltophilia in agricultural soils and antibiotic resistance properties. Res. Microbiol. 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Messiha, N.A.S.; van Diepeningen, A.D.; Farag, N.S.; Abdallah, S.A.; Janse, J.D.; van Bruggen, A.H.C. Stenotrophomonas maltophilia: A new potential biocontrol agent of Ralstonia solanacearum, causal agent of potato brown rot. Eur. J. Plant Pathol. 2007, 118, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Sitaraman, R. Pseudomonas spp. as models for plant-microbe interactions. Front. Plant Sci. 2015, 6, 787. [Google Scholar] [CrossRef] [Green Version]
- Taurian, T.; Anzuay, M.S.; Angelini, J.G.; Tonelli, M.L.; Ludueña, L.; Pena, D.; Ibáñez, F.; Fabra, A. Phosphate-solubilizing peanut associated bacteria: Screening for plant growth-promoting activities. Plant Soil 2010, 329, 421–431. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Sardinha, M.; Müller, T.; Schmeisky, H.; Joergensen, R.G. Microbial performance in soils along a salinity gradient under acidic conditions. Appl. Soil Ecol. 2003, 23, 237–244. [Google Scholar] [CrossRef]
- Aghababaei, F.; Raiesi, F.; Hosseinpur, A. The combined effects of earthworms and arbuscular mycorrhizal fungi on microbial biomass and enzyme activities in a calcareous soil spiked with cadmium. Appl. Soil Ecol. 2014, 75, 33–42. [Google Scholar] [CrossRef]
- Rasul, G.; Appuhn, A.; Müller, T.; Joergensen, R.G. Salinity-induced changes in the microbial use of sugarcane filter cake added to soil. Appl. Soil Ecol. 2006, 31, 1–10. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biol. Biochem. 1990, 22, 251–255. [Google Scholar] [CrossRef]
- Karimi, A.; Khodaverdiloo, H.; Rasooli Sadaghiani, M.H.; Khajavi, S. Changes of Microbial Indices of Inoculated Fungi and Bacteria at Hyoscyamus niger L. Rhizosphere at Different Levels of Soil Lead (Pb) Pollution. Iran. J. Soil Water Res. 2018, 49, 59–69. [Google Scholar]
- Marulanda, A.; Porcel, R.; Barea, J.M.; Azcón, R. Drought tolerance and antioxidant activities in lavender plants colonized by native drought-tolerant or drought-sensitive Glomus species. Microb. Ecol. 2007, 54, 543–552. [Google Scholar] [CrossRef]
- Rabie, G.H.; Almadini, A.M. Role of bioinoculants in development of salt-tolerance of Vicia faba plants under salinity stress. Afr. J. Biotechnol. 2005, 4, 210–222. [Google Scholar]
- Barin, M.; Aliasgharzad, N.; Samadi, A. Influence of mycorrhization on the mineral nutrition and yield of tomato under sodium chloride and salts mixture induced salinity levels. Iran. J. Soil Waters Sci. 2006, 20, 94–105. [Google Scholar]
- Zahir, Z.A.; Ghani, U.; Naveed, M.; Nadeem, S.M.; Asghar, H.N. Comparative effectiveness of Pseudomonas and Serratia sp. containing ACC-deaminase for improving growth and yield of wheat (Triticum aestivum L.) under salt-stressed conditions. Arch. Microbiol. 2009, 191, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Stajkovic, O.; Delic, D.; Josic, D.; Kuzmanovic, D.; Rasulic, N.; Knezevic-Vukcevic, J. Improvement of common bean growth by co-inoculation with Rhizobium and plant growth-promoting bacteria. Rom. Biotechnol. Lett. 2011, 16, 5919–5926. [Google Scholar]
- Hajinia, S.; Zare, M.J.; Mohammadi Goltapeh, E.; Rajali, F. Evaluation of the usefulness of endophytic fungus Piriformospora indica and Azospirillum sp. in increasing the tolerance of wheat cultivar Sardari (Triticum aestivum) to salinity stress. J. Environ. Stresses Crop Sci. 2011, 4, 21–31. [Google Scholar]
- Ahmadi Geshlagi, S.; Aliasgharzad, N.; Tavassoli, A. Evaluation of nutrients uptake and yield of mycorrhizal corn under salt stress condition. Water Soil Sci. 2014, 25, 79–89. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
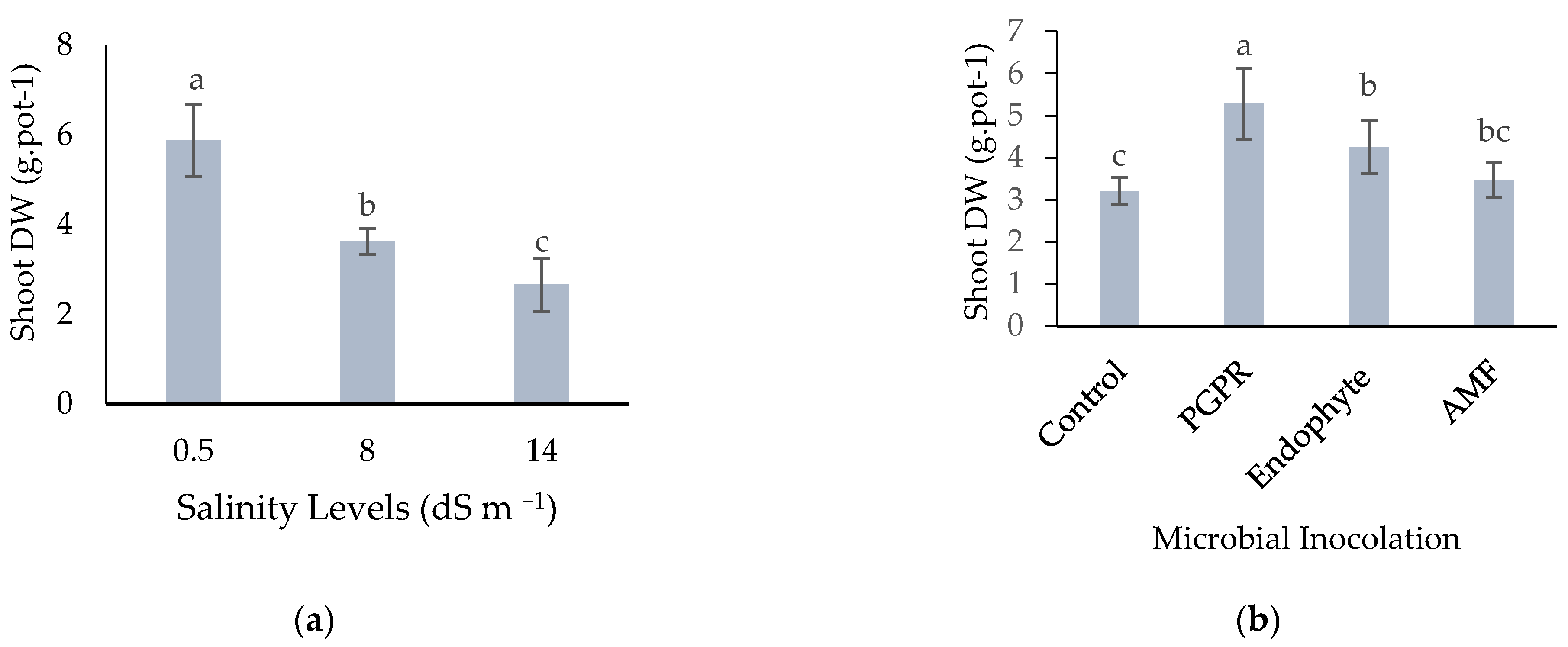{kind=link}
| Soil Texture | pH | EC (dS m−1) | Calcium Carbonate Equivalence (CCE) (%) | Organic Carbon (%) | Available Phosphorus (mg kg−1) | Available Potassium (mg kg−1) |
|---|---|---|---|---|---|---|
| Loam-clay | 7.69 | 0.50 | 19.50 | 0.50 | 7.40 | 194 |
| Isolate | Insoluble P Solubilizing Potential (cm) | Insoluble P Compound Solubilizing Potential in Broth Medium (μg mL−1) | Insoluble K Solubilizing Potential (cm) | Auxin Production | Siderophore Production | Salinity Tolerance (Up to 10%) |
|---|---|---|---|---|---|---|
| Ur83 | 3 | 24.04 | 4.16 | − | + | + |
| Ur67 | 3.4 | 26.29 | 3.3 | + | + | + |
| Ur52 | 1.2 | 20.16 | 3.5 | + | − | + |
| Isolate | Gram Staining | Obligatory Aerobic Growth | Mobility | Oxidase | Catalase | Florescent Pigment Production | Levan Production | Endospore Production | Gelatine Hydrolysis | Citrate |
|---|---|---|---|---|---|---|---|---|---|---|
| Ur83 | − | + | + | + | + | + | + | − | + | + |
| Ur67 | − | + | + | + | + | + | + | − | + | + |
| Ur52 | − | + | + | + | + | − | − | − | + | + |
| Salinity Level (dS m−1) | Microbial Biomass Carbon (mg CO2-C g−1) | Acid Phosphatase Enzyme (μg PNP g−1 h−1) | Alkaline Phosphatase Enzyme (μg PNP g−1 h−1) | Carbon Availability (μg PNP g−1 h−1) |
|---|---|---|---|---|
| 0.5 | 238.7 ± 37.28 a | 19.76 ± 2.16 a | 25.91 ± 2.19 a | 0.73 ± 0.038 a |
| 8 | 188.1 ± 38.42 b | 13.39 ± 1.88 b | 22.67 ± 2.25 b | 0.67 ± 0.039 b |
| 14 | 141.9 ± 29.49 c | 7.97 ± 1.9 c | 17.19 ± 2.01 c | 0.66 ± 0.026 a |
| Microbial Inoculation | Microbial Biomass Carbon (mg CO2-C g−1) | Acid Phosphatase Enzyme (μg PNP g−1 h−1) | Alkaline Phosphatase Enzyme (μg PNP g−1 h−1) | Metabolic Quotient (μg PNP g−1 h−1) | Carbon Availability (μg PNP g−1 h−1) |
|---|---|---|---|---|---|
| Control | 130.53 ± 34.3 c | 6.88 ± 1.3 c | 19.04 ± 2.2 b | 0.37 ± 0.03 a | 0.63 ± 0.02 b |
| PGPR | 239.06 ± 36.3 a | 21.17 ± 2.2 a | 24.64 ± 2.0 a | 0.27 ± 0.04 b | 0.70 ± 0.03 a |
| Endophytic | 177.46 ± 39.5 b | 11.77 ± 1.9 b | 20.56 ± 2.3 b | 0.33 ± 0.05 ab | 0.70 ± 0.04 a |
| AMF | 211.2 ± 30.1 a | 15.01± 2.5 b | 23.45 ± 2.1 a | 0.28 ± 0.03 b | 0.71 ± 0.06 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rouydel, Z.; Barin, M.; Rasouli-Sadaghiani, M.H.; Khezri, M.; Vetukuri, R.R.; Kushwaha, S. Harnessing the Potential of Symbiotic Endophytic Fungi and Plant Growth-Promoting Rhizobacteria to Enhance Soil Quality in Saline Soils. Processes 2021, 9, 1810. https://doi.org/10.3390/pr9101810
Rouydel Z, Barin M, Rasouli-Sadaghiani MH, Khezri M, Vetukuri RR, Kushwaha S. Harnessing the Potential of Symbiotic Endophytic Fungi and Plant Growth-Promoting Rhizobacteria to Enhance Soil Quality in Saline Soils. Processes. 2021; 9(10):1810. https://doi.org/10.3390/pr9101810
Chicago/Turabian StyleRouydel, Zahra, Mohsen Barin, Mir Hassan Rasouli-Sadaghiani, Maryam Khezri, Ramesh Raju Vetukuri, and Sandeep Kushwaha. 2021. "Harnessing the Potential of Symbiotic Endophytic Fungi and Plant Growth-Promoting Rhizobacteria to Enhance Soil Quality in Saline Soils" Processes 9, no. 10: 1810. https://doi.org/10.3390/pr9101810
APA StyleRouydel, Z., Barin, M., Rasouli-Sadaghiani, M. H., Khezri, M., Vetukuri, R. R., & Kushwaha, S. (2021). Harnessing the Potential of Symbiotic Endophytic Fungi and Plant Growth-Promoting Rhizobacteria to Enhance Soil Quality in Saline Soils. Processes, 9(10), 1810. https://doi.org/10.3390/pr9101810