Identification of Fungi in the Debitterizing Water of Apricot Kernels and Their Preliminary Evaluation on Degrading Amygdalin



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Morphology Observation of the Isolated Strains
2.3. Pretreatment of the Samples and Extraction of Genomic DNA
2.4. PCR Amplification, Recovery and Purification of the PCR Products
2.5. Preparation of the Medium and Strain Culture
2.6. Determination of Amygdalin by HPLC
2.7. Degradation Investigation of the Amygdalin Caused by the Three Strains
2.8. Measurement of the Beta-Glucosidase (β-GC) Activity
2.9. Statistical Analysis
3. Results and Discussion
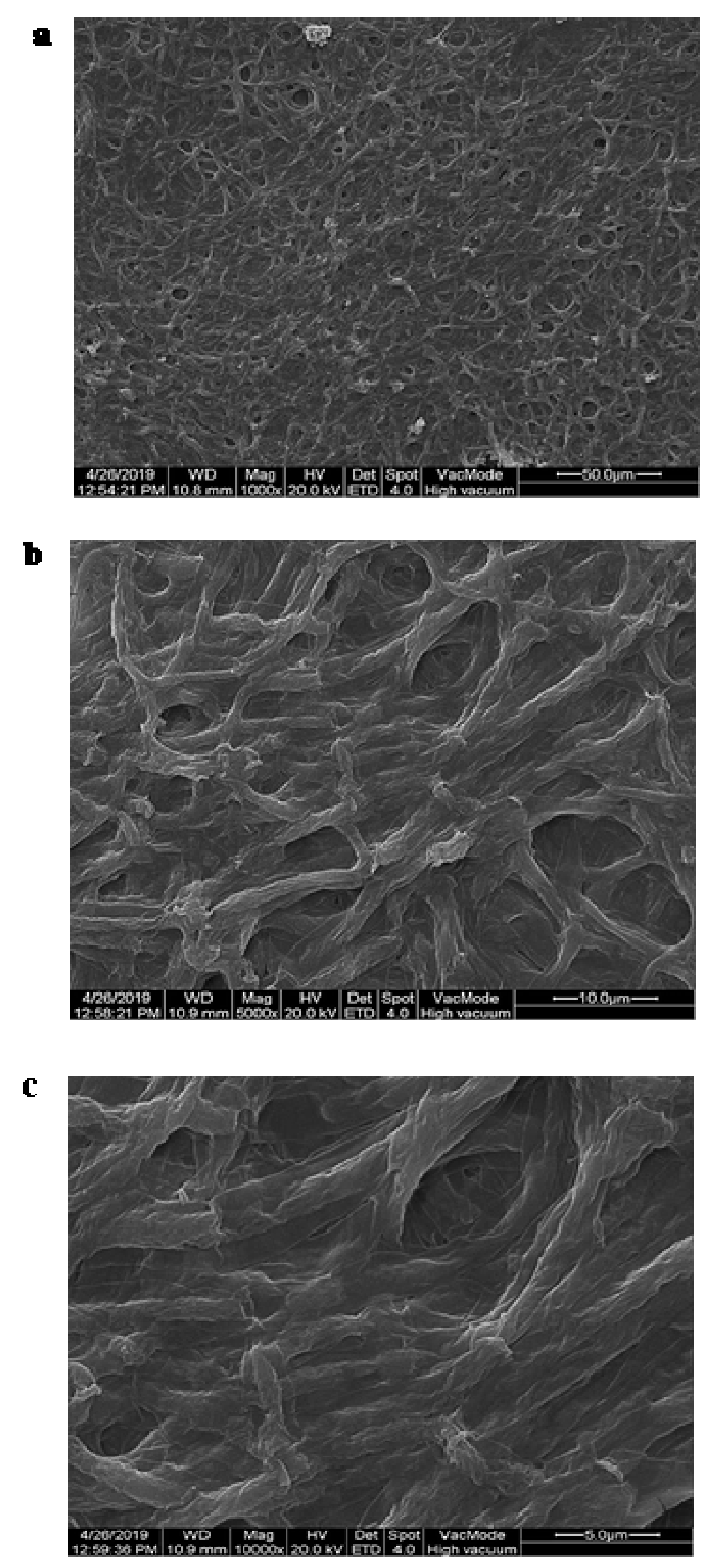
3.1. Morphology Characterization of Strains
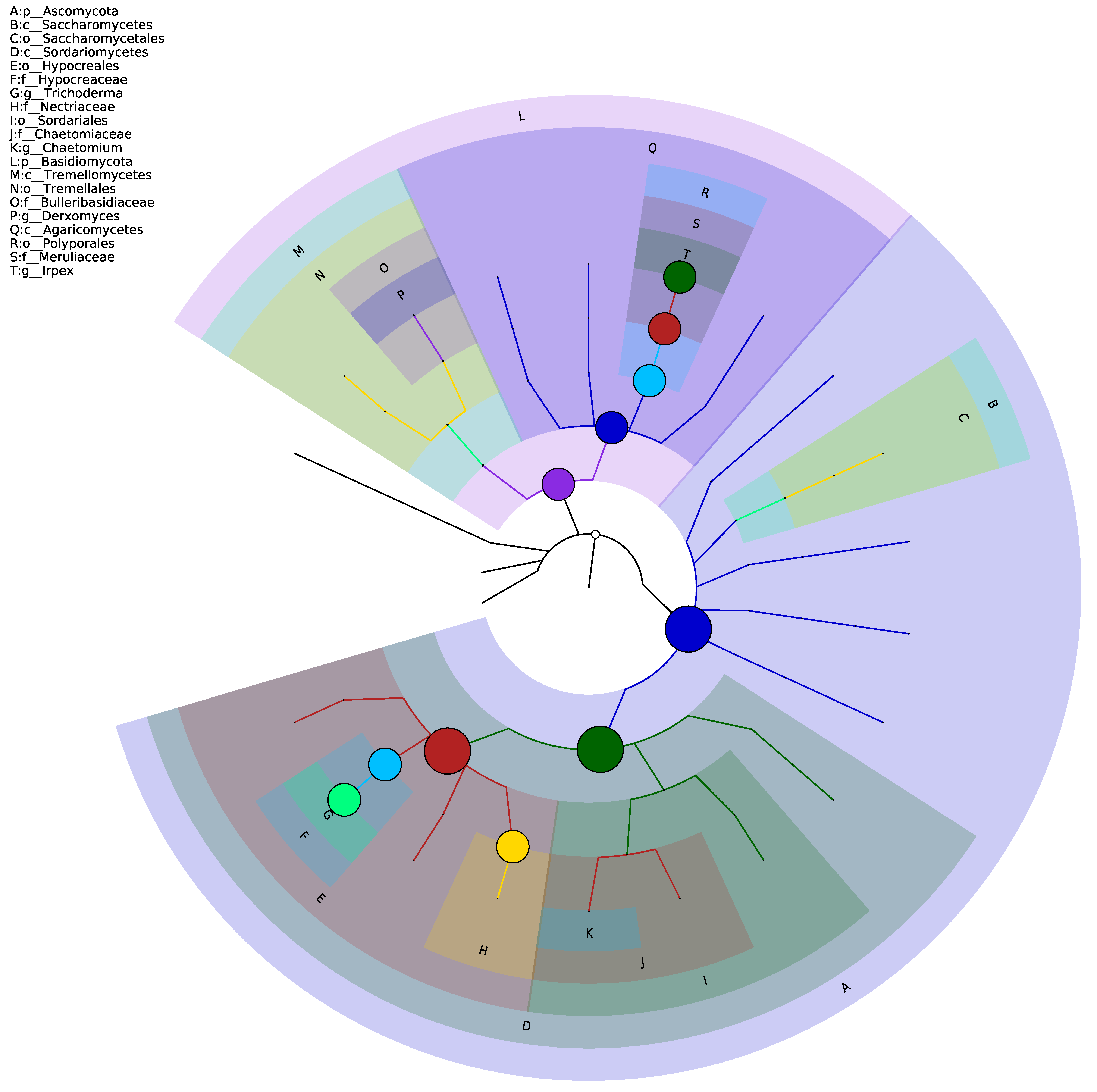
3.2. Diversity Analysis of the Fungal in the Samples
3.3. Distribution of the Fungal Colonies at Different Levels
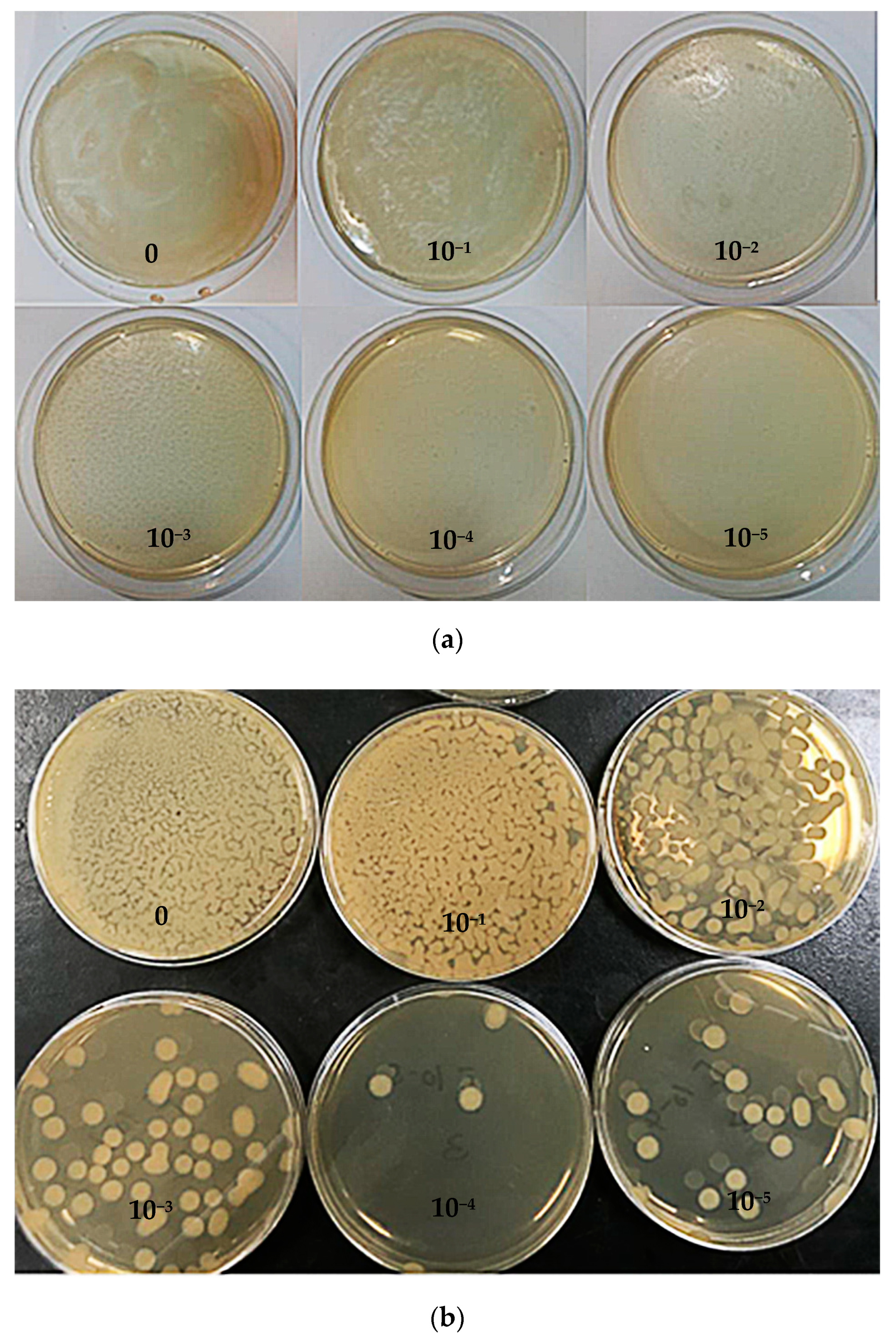
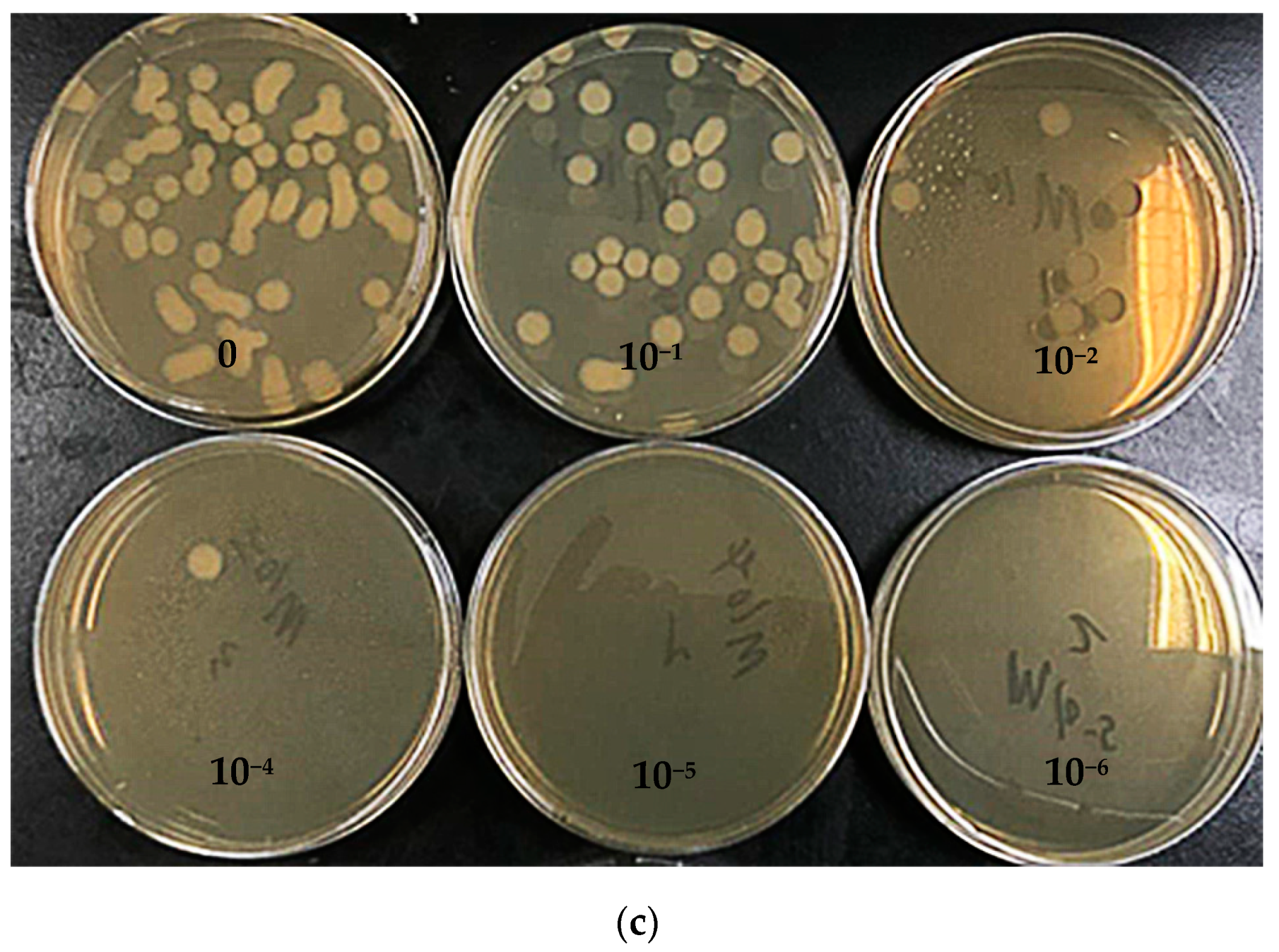
3.4. Colony Count in the Culture Medium
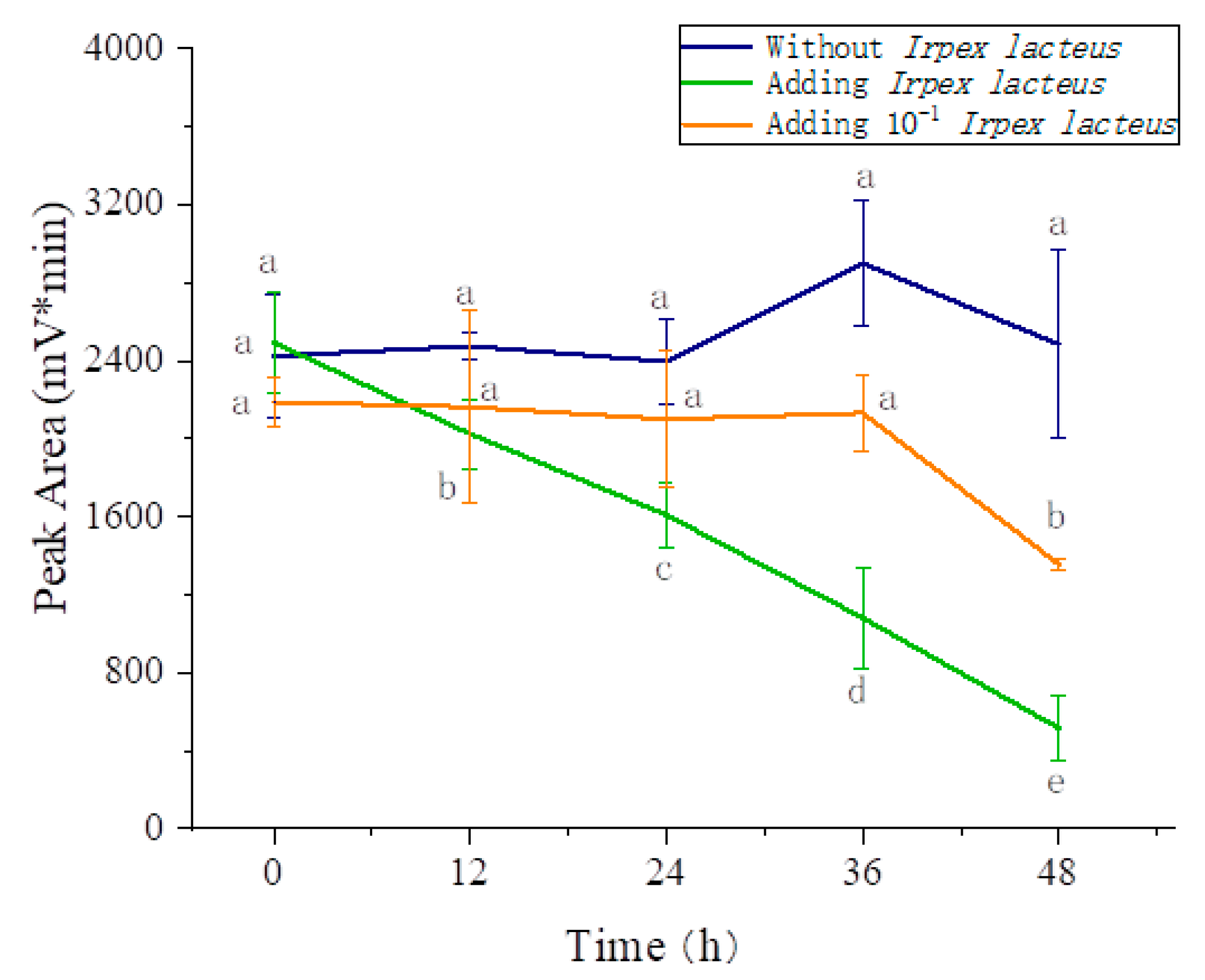
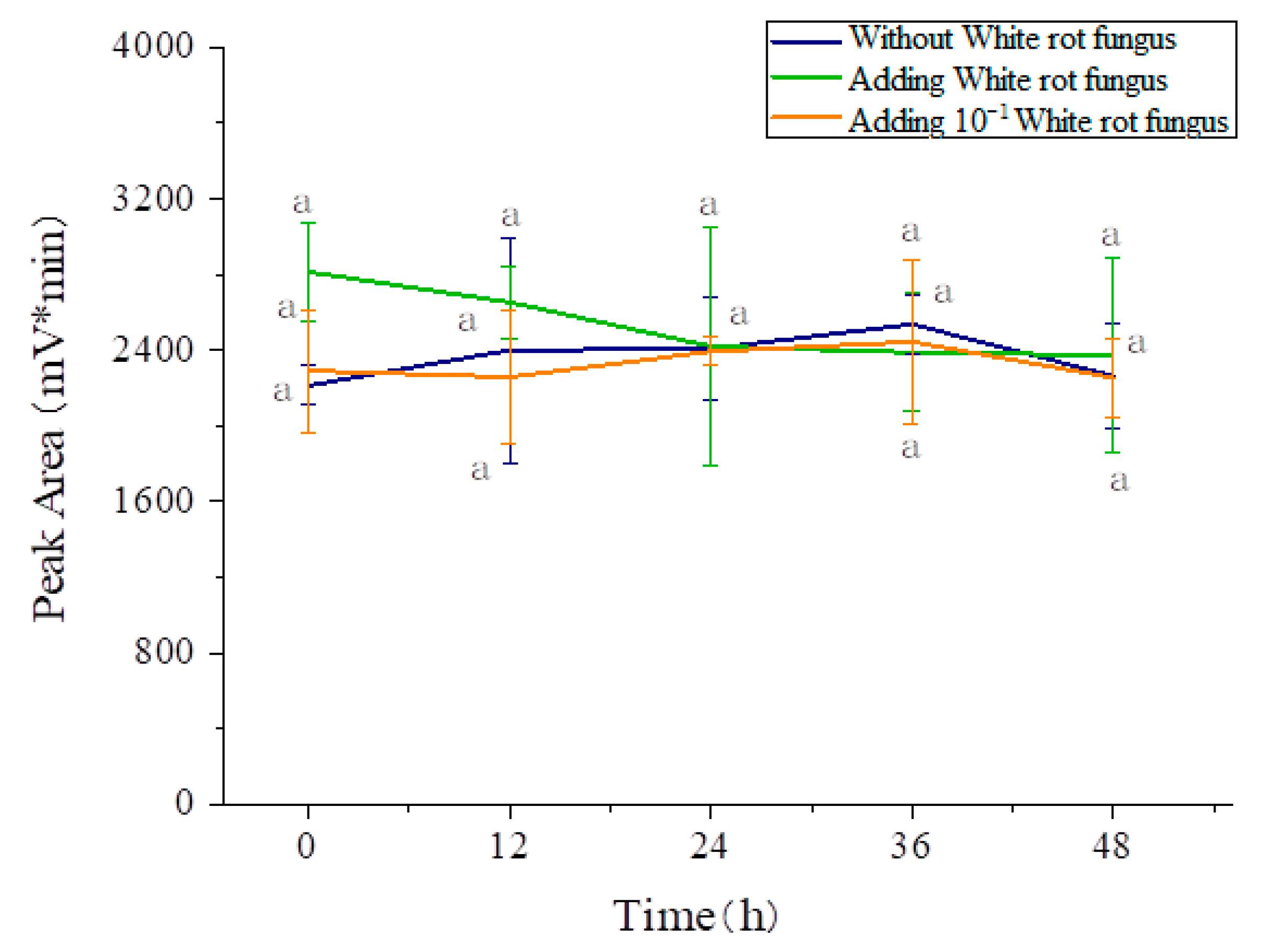
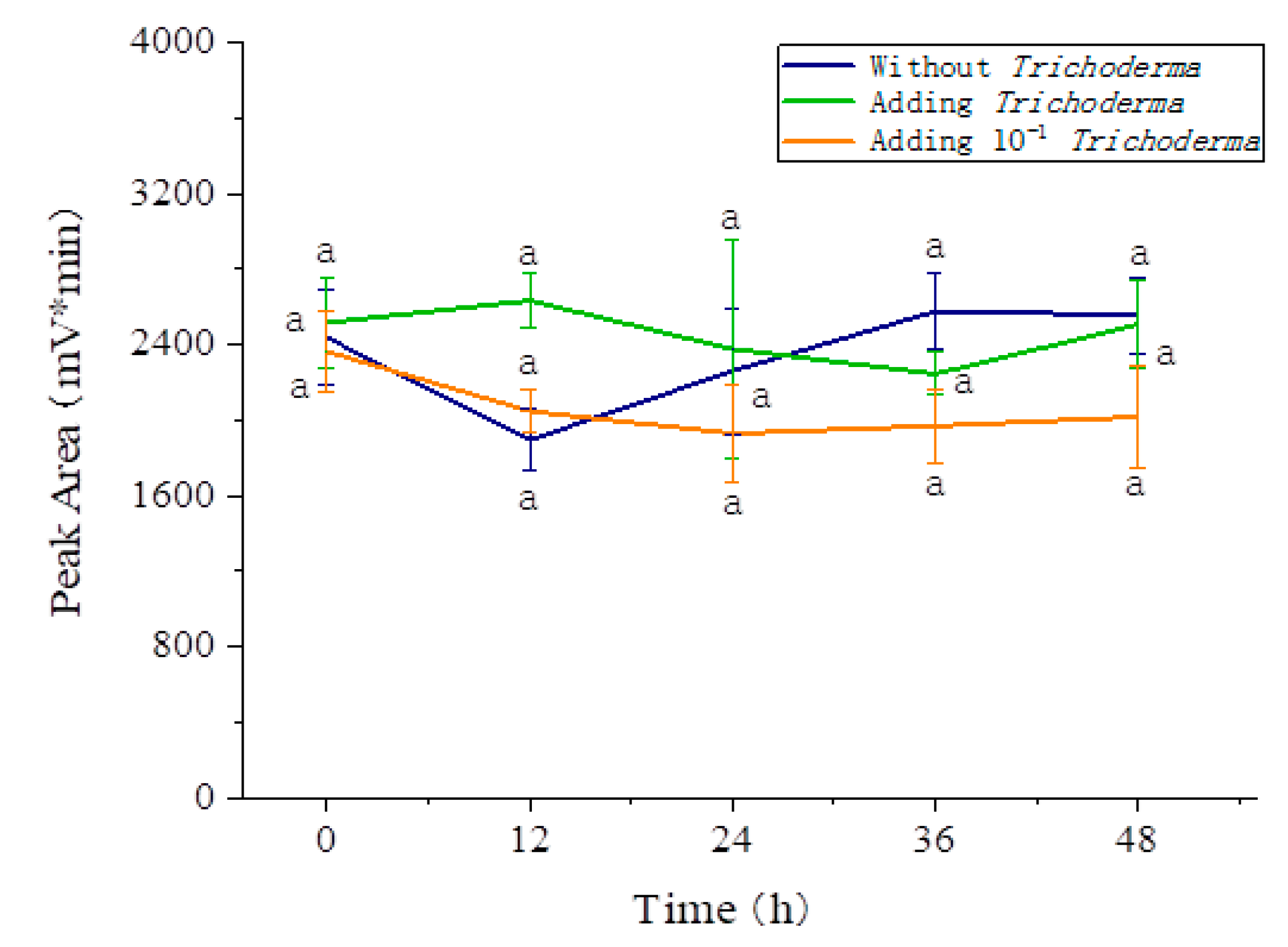
3.5. Effects of the Three Fungi on the Degradation of Amygdalin
3.6. Effects of the Three Fungi on the Activity of β-Glucosidase
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bolarinwa, I.F.; Orfila, C.; Morgan, M.R.A. Amygdalin content of seeds, kernels and food products commercially-available in the UK. Food Chem. 2014, 152, 133–139. [Google Scholar] [CrossRef]
- Dixit, A.K. Drying characteristics of wild apricot (Prunus armeniaca) fruit bar and economic evaluation of market potential of the enterprise. Indian J. Agric. Sci. 2018, 83. [Google Scholar] [CrossRef]
- Zacharias, E. Carbohydrates; Springer: New York, NY, USA, 2012; pp. 127–139. [Google Scholar] [CrossRef]
- Zhang, Q.-A.; Wei, C.-X.; Fan, X.-H.; Shi, F.-F. Chemical compositions and antioxidant capacity of by-products generated during the apricot kernels processing. CyTA J. Food 2018, 16, 422–428. [Google Scholar] [CrossRef]
- Abboud, M.M.; Awaida, W.A.; Alkhateeb, H.H.; Abu-Ayyad, A.N. Antitumor action of amygdalin on human breast cancer cells by selective sensitization to oxidative stress. Nutr. Cancer 2019, 71, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Pan, W.; Lv, J.; Wu, H. Protective effect of amygdalin on LPS-induced acute lung injury by inhibiting NF-κB and NLRP3 signaling pathways. Inflammation 2017, 40, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Bolarinwa, I.F.; Orfila, C.; Morgan, M.R.A. Determination of amygdalin in apple seeds, fresh apples and processed apple juices. Food Chem. 2015, 170, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, Q.-A.; Yao, J.-L.; Zhang, X.-Y. Changes of amygdalin and volatile components of apricot kernels during the ultrasonically-accelerated debitterizing. Ultrason. Sonochem. 2019, 58, 104614. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Jiao, Z.; Liu, J.; Liu, H.; Zhang, C.; Yang, W. Response surface optimization of ultrasonic-assisted extraction and HPCPC purification of amygdalin from bitter apricot kernel meal (Prunus armeniaca). Food Sci. 2016, 37, 81–87. [Google Scholar] [CrossRef]
- Silem, A.; Günter, H.O.; Einfeldt, J.; Boualia, A. The occurrence of mass transport processes during the leaching of amygdalin from bitter apricot kernels: Detoxification and flavour improvement. Int. J. Food Sci. Technol. 2006, 41, 201–213. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, X.-Y.; Fan, X.-H.; Zhang, Q.-A. Optimization on fast debitterizing technologies of apricot seed by ultrasound with response surface methodology. Food Mach. 2018, 34, 189–194. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, Q.-A.; Yao, J.-L. Consecutive membrane filtration and re-utilization of the debitterizing wastewater of apricot kernels for a flavor beverage-making. J. Clean. Prod. 2020, 262, 121360. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, Q.-A.; Fan, X.-H.; Zhang, X.-Y. Effect of debitterizing treatment on the quality of the apricot kernels in the industrial processing. J. Food Proc. Preser. 2017, 42, 1–8. [Google Scholar] [CrossRef]
- Jin, Y.N.; Wu, M.F.; Zhao, G.H.; Li, M.F. Photocatalysis-enhanced electrosorption process for degradation of high-concentration dye wastewater on TiO2/carbon aerogel. Chem. Eng. J. 2011, 168, 1248–1255. [Google Scholar] [CrossRef]
- Nout, M.J.; Tunçel, G.; Brimer, L. Microbial degradation of amygdalin of bitter apricot seeds (Prunus armeniaca). Int. J. Food Microbiol. 1995, 24, 407–412. [Google Scholar] [CrossRef]
- Zhu, J.; Sanidad, K.Z.; Sukamtoh, E.; Zhang, G. Potential roles of chemical degradation in the biological activities of curcumin. Food Funct. 2017, 8, 907–914. [Google Scholar] [CrossRef]
- Chen, L.; Wu, H. Research advances in degradation of polysaccharides. Chin. Arch. Tradit. Chin. Med. 2008, 26, 133–135. (In Chinese) [Google Scholar] [CrossRef]
- Pan, Z. Research development of the degradation methods of lignin. Guangdong Chem. Indus. 2015, 42, 100–101. (In Chinese) [Google Scholar] [CrossRef]
- Hu, Y.; Catchmark, J.M. Integration of cellulases into bacterial cellulose: Toward bioabsorbable cellulose composites. J. Biomed. Mater. Res. Part B 2011, 97, 114–123. [Google Scholar] [CrossRef]
- Purschke, B.; Meinlschmidt, P.; Horn, C.; Rieder, O.; Jäger, H. Improvement of techno-functional properties of edible insect protein from migratory locust by enzymatic hydrolysis. Eur. Food Res. Technol. 2018, 244, 1–15. [Google Scholar] [CrossRef]
- Hao, T.; Yan, J.M.; Wan, Y.L.; Jian, W.W. Efficient degradation of triclosan by an endophytic fungus Penicillium oxalicum B4. Environ. Sci. Pollut. Res. 2018, 25, 8963–8975. [Google Scholar] [CrossRef]
- Feng, Y.M.; Fan, X.H.; Zhan, H.; Teng, S.Y.; Yang, F.; Chen, S.H. Research progress on ecotoxicity and microbial degradation of strobilurin fungicides. Biotechnol. Bull. 2017, 33, 52–58. [Google Scholar] [CrossRef]
- Meng, Y.; Fu, Y.B.; Liang, S.K.; Chen, W.; Liu, Z.H. Effects of oil pollutants on the performance of marine benthonic microbial fuel cells and its acceleration of degradation. Environ. Sci. 2015, 36, 3080–3085. [Google Scholar] [CrossRef]
- Hechmi, N.; Bosso, L.; El-Bassi, L.; Scelza, R.; Testa, A.; Jedidi, N.; Rao, M.A. Depletion of pentachlorophenol in soil microcosms with byssochlamys nivea and scopulariopsis brumptii as detoxification agents. Chemosphere 2016, 165, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Yan, M.; Treu, L.; Angelidaki, I.; Fotidis, I.A. Hydrogenotrophic methanogens are the key for a successful bioaugmentation to alleviate ammonia inhibition in thermophilic anaerobic digesters. Bioresour. Technol. 2019, 293, 122070. [Google Scholar] [CrossRef]
- Zhang, X.; Cai, T.; Xu, X. Isolation and identification of a tetracycline-degrading bacterium and optimizing condition for tetracycline degradation. Biotechnol. Bull. 2015, 1, 173–180. (In Chinese) [Google Scholar] [CrossRef]
- Yang, T.; Yan, H.L. Comparative analysis of bacteria associated with different mosses by 16S rRNA and 16S rDNA sequencing. J. Basic Microbiol. 2016, 57, 57–67. [Google Scholar] [CrossRef]
- Cao, B.Y.; Wang, M.; Liu, L.; Zhou, Z.M.; Wen, S.P.; Rozalskif, A.; Wang, L. 16S-23S rDNA internal transcribed spacer regions in four Proteus species. J. Microbiol. Methods 2009, 77, 109–118. [Google Scholar] [CrossRef]
- Wang, P.; Cui, Z.L.; Liu, B.; Sun, B.; Cao, H. Effect of cultivation methods on bacterial diversity by PCR-rflp analysis. Acta Pedol. Sin. 2009, 46, 1096–1101. [Google Scholar] [CrossRef]
- Wang, L.; Li, B.; Zhu, J. Review of high-throughput sequencing techniques on constructed wetland microbial diversity. Chin. Agric. Sci. Bull. 2016, 32, 10–15. [Google Scholar]
- Xiong, W.; Li, H.T.; Zhan, A.B. Early detection of invasive species in marine ecosystems using high-throughput sequencing: Technical challenges and possible solutions. Mar. Biol. 2016, 163, 139. [Google Scholar] [CrossRef]
- Dalmassoa, A.; Civeraa, T.; Pattonoa, D.; Cardazzob, B.; Botteroa, M.T. Characterization of microbiota in Plaisentif cheese by high-throughput sequencing. LWT Food Sci. Technol. 2016, 69, 490–496. [Google Scholar] [CrossRef]
- He, G.Q.; Liu, T.J.; Sadiq, F.A.; Gu, J.S.; Zhang, G.H. Insights into the microbial diversity and community dynamics of Chinese traditional fermented foods from using high throughput sequencing approaches. J. Zhejiang Univer. Sci. B 2017, 18, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.Z.; Ding, Z.W.; Ding, J.; Fu, L.; Zeng, R.J. Design and evaluation of universal 16S rRNA gene primers for high-throughput sequencing to simultaneously detect DAMO microbes and anammox bacteria. Water Res. 2015, 87, 385–394. [Google Scholar] [CrossRef]
- Bary, A.D.; Balfour, I.B. Comparative Morphology and Biology of the Fungi, Mycetozoa and Bacteria; Clarendon Press: Oxford, UK, 2015; p. 554. [Google Scholar]
- Pazouki, M.; Panda, T. Understanding the morphology of fungi. Bioprocess Eng. 2000, 22, 127–143. [Google Scholar] [CrossRef]
- Bosso, L.; Lacatena, F.; Varlese, R.; Nocerino, S.; Cristinzio, G.; Russo, D. Plant pathogens but not antagonists change in soil fungal communities across a land abandonment gradient in a Mediterranean landscape. Acta Oecol. 2017, 78, 1–6. [Google Scholar] [CrossRef]
- Mohanty, S.; Ghosh, S.; Nayak, S.; Das, A.P. Isolation, identification and screening of manganese solubilizing fungi from low-grade manganese ore deposits. Geomicrobiol. J. 2017, 34, 309–316. [Google Scholar] [CrossRef]
- Anandan, R.; Deenathayalan, T.; Prakash, M.; Sunilkumar, B.; Narayanan, G.S. Sassessment of genetic diversity among sesame (Sesamum indicum L.) germplasm as revealed by RAPD and SSR markers. Indian J. Biochem. Biophys. 2018, 55, 143–150. [Google Scholar]
- Sun, K.; Wei, S.; Liu, Y.L.; Xu, R.S.; Liu, Z.Y.; Yin, J.P.; Yuan, J.J.; Wu, X.Y. Real-time PCR for the detection of Lactobacillus plantarum based on dual priming oligonucleotide system. Food Fermn. Ind. 2016, 42, 178–183. (In Chinese) [Google Scholar] [CrossRef]
- De, A.; Jorgensen, A.N.; Beatty, W.L.; Lemos, J.; Wen, Z.T. Deficiency of MecA in streptococcus mutans causes major defects in cell envelope biogenesis, cell division, and biofilm formation. Front. Microbiol. 2018, 9, 2130. [Google Scholar] [CrossRef]
- Li, M.; Du, L.N.; Zhang, Y.; Zhang, N.; Teng, L.R. Improvement of medium for the fermentation of Irpex lacteus Fr.by respond surface methodology coupled with desirability function. Chin. J. Pharm. 2011, 42, 17–20. (In Chinese) [Google Scholar] [CrossRef]
- Wang, M.L. Study on optimization of wastewater with DDNP of high concentration by medium of white rot fungus. Shanxi Chem. Ind. 2015, 35, 78–81. (In Chinese) [Google Scholar] [CrossRef]
- Lv, S.X.; Yu, X.D.; Zhang, C.X.; Chen, J. Influence of fermentation conditions on hyphae growth and chitinase activity of Trichoderma strain of T23. J. Shenyang Agric. Univ. 2005, 36, 332–335. (In Chinese) [Google Scholar] [CrossRef]
- Huson, D.H.; Weber, N. Microbial community analysis using MEGAN. Methods Enzymol. 2013, 531, 465–485. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.J.; Li, Z.J.; Tian, Z.Q. Analysis of bacterial diversity in Daqu and fermented grains for Maotai-flavor liquor. Food Sci. 2019, 40, 152–159. [Google Scholar] [CrossRef]
- Brimer, L.; Tuncel, G.; Nout, M.J.R. Simple screening procedure for microorganisms to degrade amygdalin. Biotechnol. Tech. 1993, 7, 683–687. [Google Scholar] [CrossRef]
- Xu, C.Y. Advance in decolorization and biodegradation of dye by white rot fungus. Prog. Biotechnol. 2002, 22, 57–60. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagents | Control | Measurement | Standard Tube |
|---|---|---|---|
| pNPG (μL) | 400 | ||
| Disodium hydrogen phosphate-citric acid buffer (μL) | 500 | 500 | |
| β-glucosidase crude enzyme solution (μL) | 100 | 100 | |
| Operation step 1 | |||
| pNPG (μL) | 400 | ||
| Operation step 2 | |||
| Supernatant (μL) | 500 | 500 | |
| p-Nitrophenol standard solution (μL) | 500 | ||
| Na2CO3 (μL) | 1000 | 1000 | 1000 |
| Samples | Simpson | Chao1 | ACE | Shannon |
|---|---|---|---|---|
| 1 | 0.0386 | 26.00 | 26.00 | 0.19 |
| 2 | 0.0029 | 35 | 31.44 | 0.02 |
| 3 | 0.485 | 161.54 | 168.31 | 2.19 |
| Level | OUT ID | 1 | 2 | 3 |
|---|---|---|---|---|
| Phylum | Ascomycota | 1.9% | 99.9% | 99.8% |
| Basidiomycota | 98.1% | 0.1% | 0.1% | |
| Other | 0.1% | |||
| Class | Sordariomycetes | 1.9% | 99.9% | 99.7% |
| Agaricomycetes | 98.1% | 0.1% | 0.1% | |
| Other | 0.1% | |||
| Tremellomycetes | 0.1% | |||
| Order | Hypocreales | 1.9% | 99.9% | 99.7% |
| Polyporales | 98.0% | 0.1% | 0.1% | |
| Other | 0.1% | |||
| Tremellales | 0.1% | |||
| Sordariales | 0.0% | |||
| Family | Hypocreaceae | 1.8% | 99.6% | |
| Nectriaceae | 99.9% | |||
| Meruliaceae | 98.0% | |||
| Other | 0.1% | |||
| Genus | Trichoderma | 1.8% | 99.6% | |
| Unidentified | 99.9% | |||
| Irpex | 98.0% | 0.1% | 0.1% | |
| Other | 0.1% |
| Samples | Average of Colony (pcs) | Colony Count (CFU/mL) |
|---|---|---|
| Iprex lacteus | 64 ± 7 | 6.4 × 107 |
| White rot fungi | 54 ± 5 | 5.4 × 108 |
| Trichoderma | 68 ± 11 | 6.8 × 108 |
| Time | β-Glucosidase Activity (U/104 cell) | ||
|---|---|---|---|
| Irpex lacteus | Trichoderma | White Rot Fungus | |
| 0 h | −0.10 ± 0.16 | −0.41 ± 0.26 | −0.26 ± 0.15 |
| 12 h | −0.22 ± 0.19 | −0.33 ± 0.16 | −0.10 ± 0.33 |
| 24 h | −0.45 ± 0.17 | −0.88 ± 0.19 | −0.26 ± 0.30 |
| 36 h | −0.28 ± 0.09 | −0.52 ± 0.19 | −0.11 ± 0.23 |
| 48 h | −0.30 ± 0.03 | −0.32 ± 0.24 | −0.05 ± 0.09 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Zhang, Q.-A.; Yao, J.-L.; García-Martín, J.F. Identification of Fungi in the Debitterizing Water of Apricot Kernels and Their Preliminary Evaluation on Degrading Amygdalin. Processes 2020, 8, 1061. https://doi.org/10.3390/pr8091061
Zhang N, Zhang Q-A, Yao J-L, García-Martín JF. Identification of Fungi in the Debitterizing Water of Apricot Kernels and Their Preliminary Evaluation on Degrading Amygdalin. Processes. 2020; 8(9):1061. https://doi.org/10.3390/pr8091061
Chicago/Turabian StyleZhang, Ning, Qing-An Zhang, Jian-Li Yao, and Juan Francisco García-Martín. 2020. "Identification of Fungi in the Debitterizing Water of Apricot Kernels and Their Preliminary Evaluation on Degrading Amygdalin" Processes 8, no. 9: 1061. https://doi.org/10.3390/pr8091061
APA StyleZhang, N., Zhang, Q.-A., Yao, J.-L., & García-Martín, J. F. (2020). Identification of Fungi in the Debitterizing Water of Apricot Kernels and Their Preliminary Evaluation on Degrading Amygdalin. Processes, 8(9), 1061. https://doi.org/10.3390/pr8091061