Application of Laser-Induced Fluorescence in Functional Studies of Photosynthetic Biofilms



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
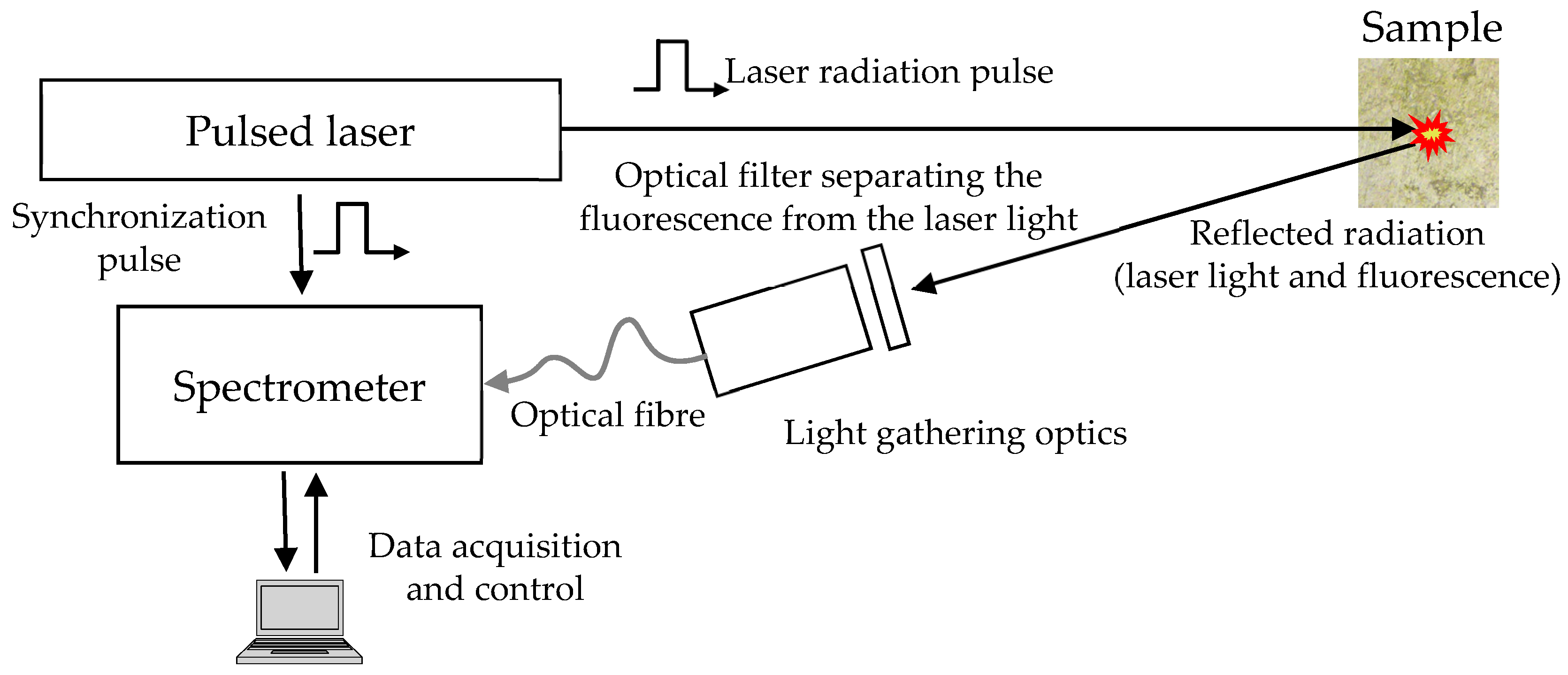
2. Laser-Induced Fluorescence (LIF)
- the early detection and evaluation of biofilms may require a high overall sensitivity of the sensor;
- the characteristic peaks due to the biofilm fluorescence are relatively wide, with the FWHM (full width at half maximum) of the order of tens of nanometers;
- the instrument for the field measurements must be robust and of low power consumption.
3. Application of LIF in Plant Sciences
4. Photosynthetic Biofilms
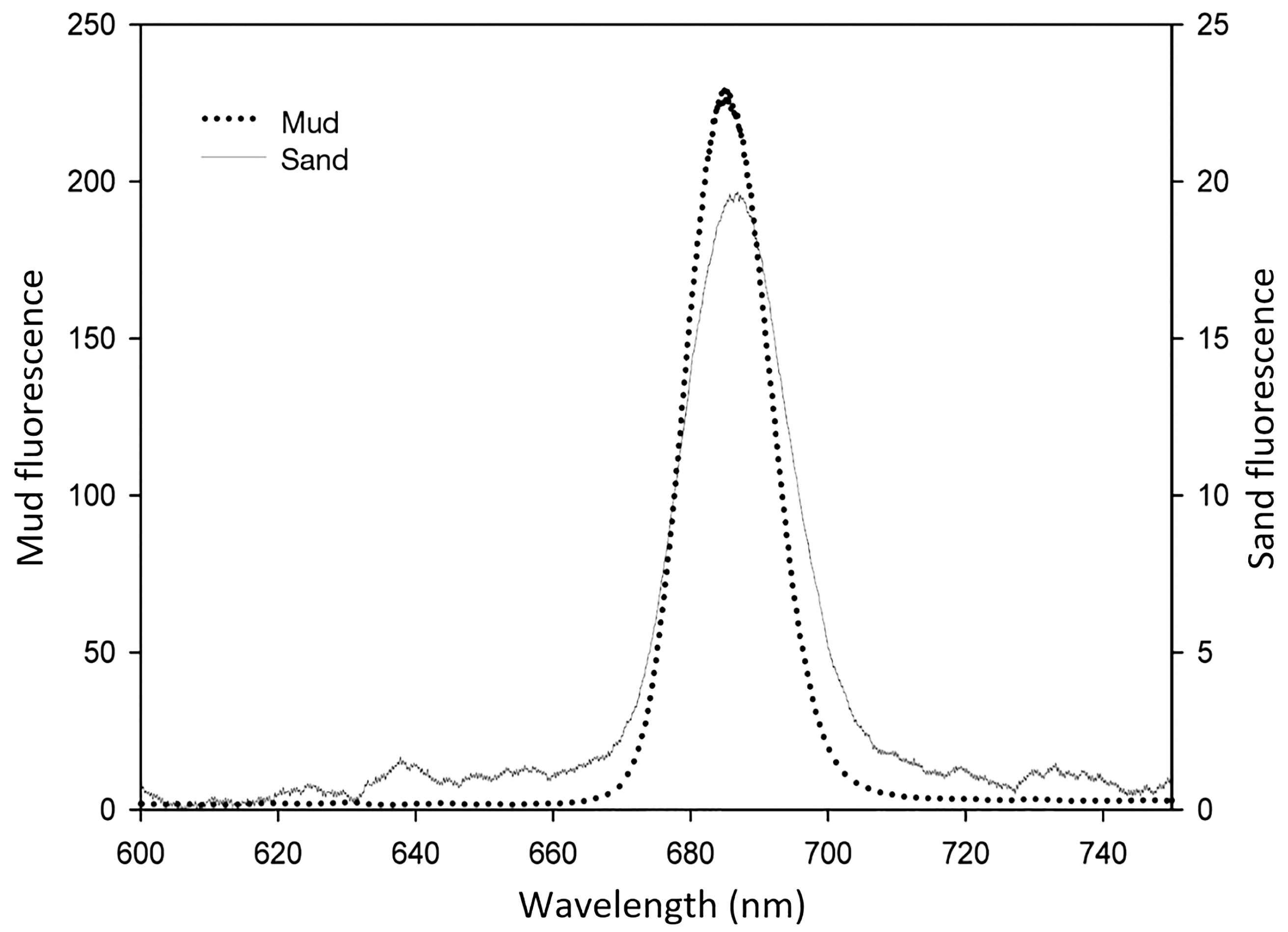
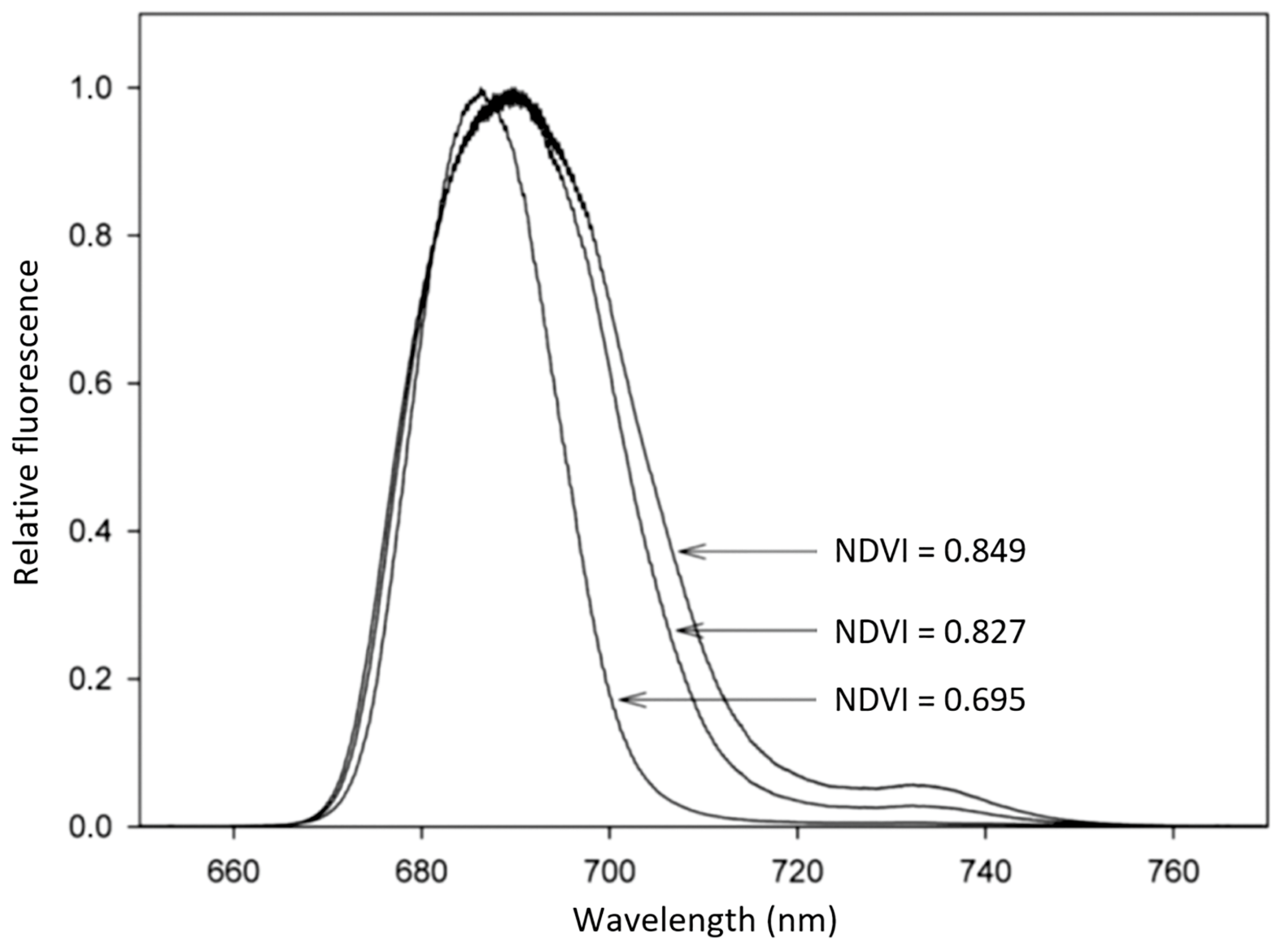
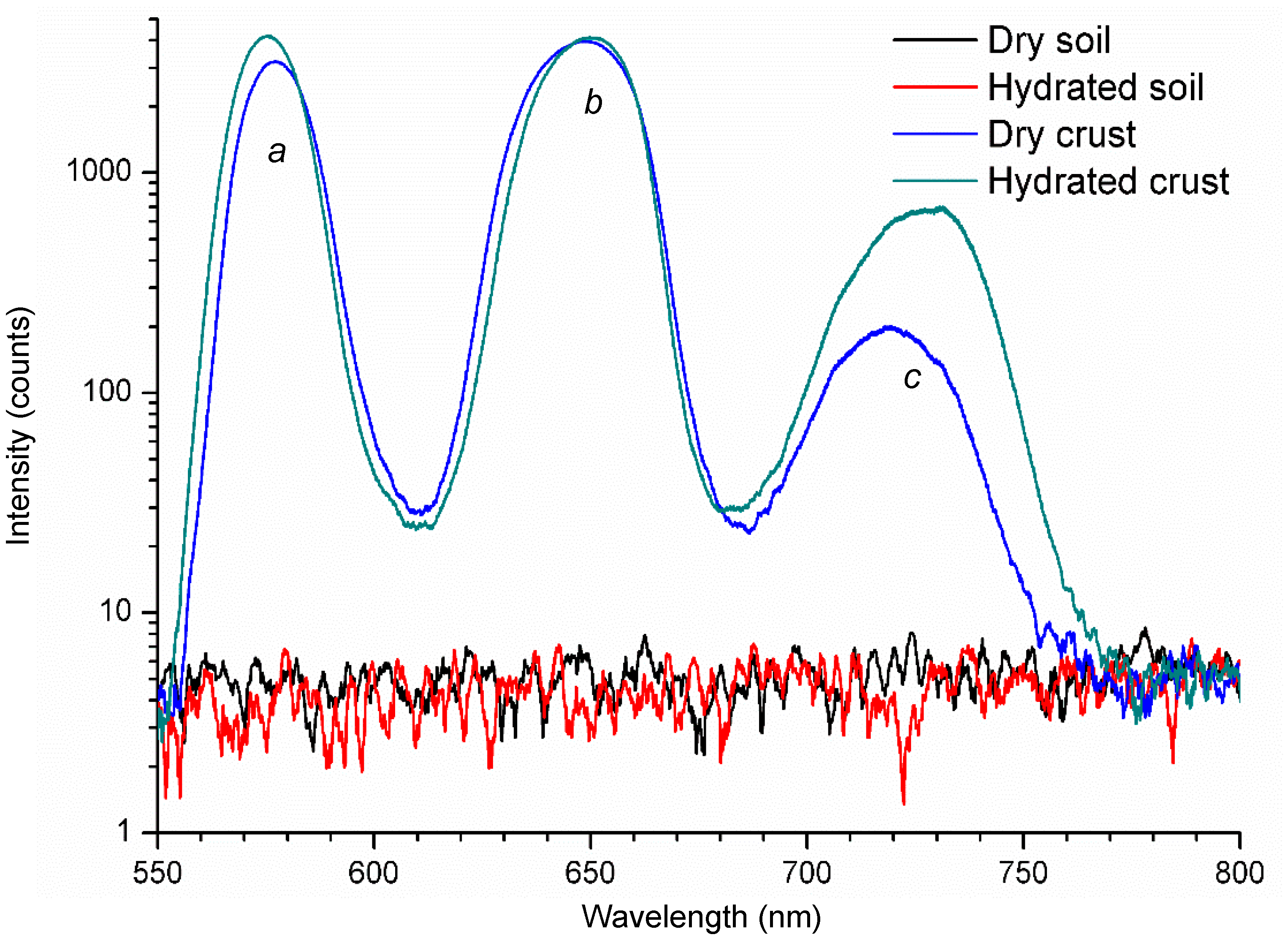
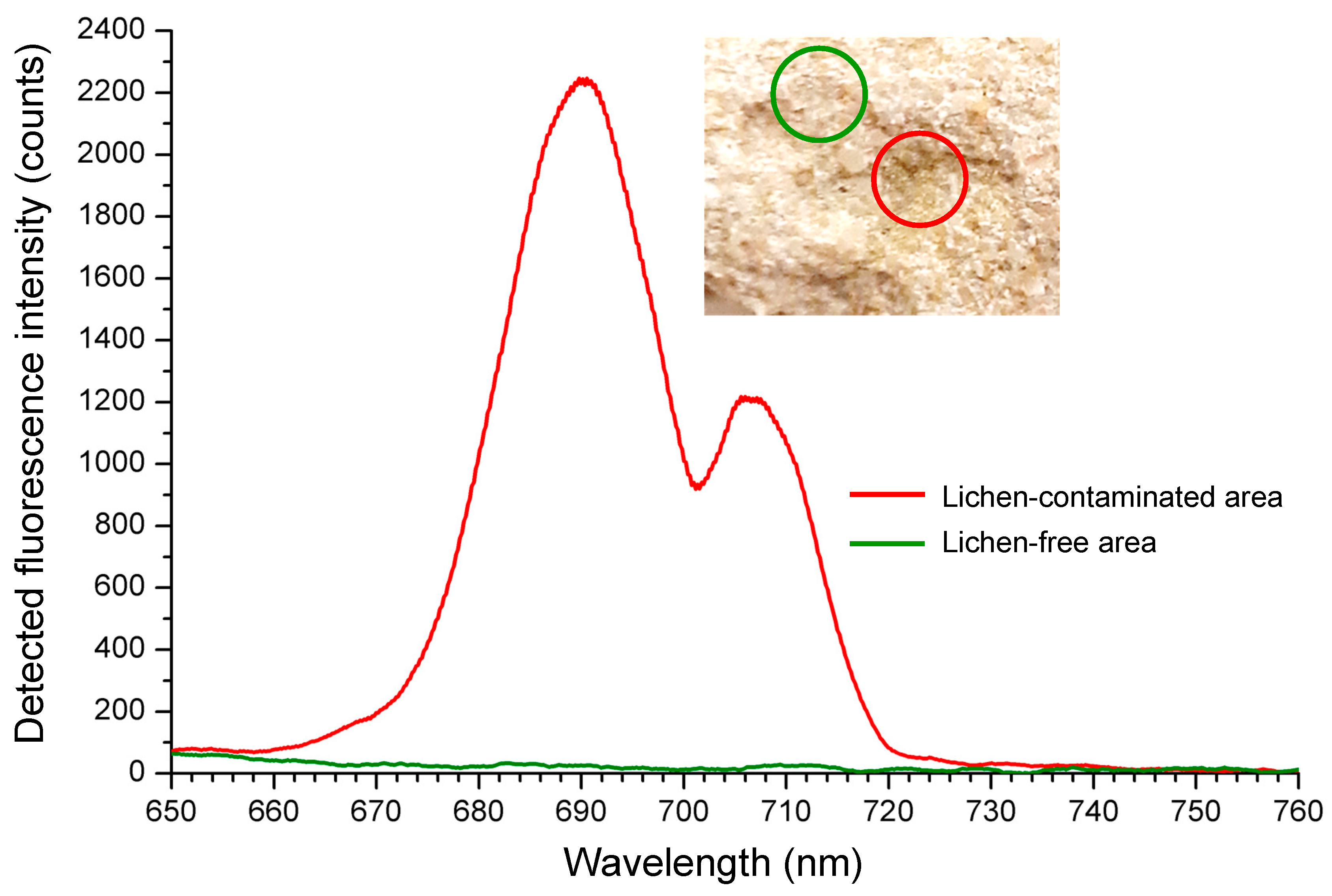
5. Application of LIF to Photosynthetic Biofilm
6. Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marques da Silva, J.; Bernardes da Silva, A.; Pádua, M. Modulated chlorophyll a fluorescence: A tool for teaching photosynthesis. J. Biol. Educ. 2007, 41, 178–183. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Marques da Silva, J.; Arrabaça, M.C. Characteristics of fluorescence emission by leaves of nitrogen starved Paspalum dilatatum POIR. Photosynthetica 1992, 26, 253–256. [Google Scholar]
- Serôdio, J.; Marques da Silva, J.; Catarino, F. Relationship between chlorophyll fluorescence quenching and O2 evolution in microalgae. In Photosynthesis: Mechanisms and Effects; Garab, G., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; Volume IV, pp. 2585–2588. [Google Scholar]
- Campbell, D.; Hurry, V.; Clarke, A.K.; Gustafsson, P.; Öquist, G. Chlorophyll fluorescence analysis of cyanobacterial photosynthesis and acclimation. Microbiol. Mol. Biol. Rev. 1998, 62, 667–683. [Google Scholar] [PubMed]
- Besemer, K.; Hödl, I.; Sikger, G.; Battin, T.J. Architectural differentiation reflects bacterial community structure in stream biofilms. ISME J. 2009, 3, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Serôdio, J.; Marques da Silva, J.; Catarino, F. Nondestructive tracing of migratory rhythms of intertidal benthic microalgae using in vivo chlorophyll a fluorescence. J. Phycol. 1997, 33, 542–553. [Google Scholar] [CrossRef]
- Vieira, S.; Ribeiro, L.; Jesus, B.; Cartaxana, P.; Marques da Silva, J. Photosynthesis assessment in microphytobenthos with conventional and imaging pulse amplitude modulation fluorometry. Photochem. Photobiol. 2013, 89, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y. Diurnal variations of chlorophyll fluorescence and CO2 exchange of biological soil crusts in different successional stages in the Gurbantunggut Desert of northwestern China. Ecol. Res. 2014, 29, 289–298. [Google Scholar] [CrossRef]
- Thorlabs: Air-Spaced Doublet Collimators. Available online: https://www.thorlabs.com/newgrouppage9.cfm?objectgroup_id=1337 (accessed on 21 March 2018).
- GWU-Lasertechnik. Spectrelle® 5000 Compact High Resolution Echelle Spectrograph; GWU Group: Erftstadt, Germany, 2006. [Google Scholar]
- Edmund Optics. Optical and Optical Instruments Catalog, Spring 2012; Edmund Optics: Barrington, IL, USA, 2012; p. 448. [Google Scholar]
- Innovative Solutions for Your Application Needs; B&W Tek: Newark, NJ, USA, 2012; pp. 11–27.
- Thorlabs. CCS Series Spectrometer: Operational Manual. Available online: https://www.thorlabs.com/Thorcat/18100/18143-D02.pdf (accessed on 21 March 2018).
- Ibsen Photonics. How to Choose a Spectrometer. Available online: https://ibsen.com/technology/spectrometer-tutorial/how-to-choose-a-spectrometer/ (accessed on 21 March 2018).
- Ocean Optics. What’s the Right Spectrometer for You? Available online: https://oceanoptics.com/wp-content/uploads/What-is-the-Right-Spectrometer-for-You.pdf (accessed on 21 March 2018).
- Czerny, M.; Turner, A.F. Über den astigmatismus bei spiegelspektrometern. Zeitschrift für Physik 1930, 61, 792–797. [Google Scholar] [CrossRef]
- Utkin, A.B.; Vieira, S.; Marques da Silva, J.; Lavrov, A.; Leite, E.; Cartaxana, P. Compact low-cost detector for in vivo assessment of microphytobenthos using laser induced fluorescence. Opt. Spectrosc. 2013, 114, 471–477. [Google Scholar] [CrossRef]
- Ocean Optics. Flame Optical Bench Options. Available online: https://oceanoptics.com/product-details/flame-optical-bench-options/ (accessed on 3 May 2018).
- Ocean Optics. Spectrometers. Available online: https://oceanoptics.com/wp-content/uploads/OceanOptics_Spectrometers.pdf (accessed on 3 May 2018).
- Toshiba. Toshiba CCD Linear Image Sensor TCD1304AP. Available online: https://oceanoptics.com/wp-content/uploads/Toshiba-TCD1304AP-CCD-array.pdf (accessed on 3 May 2018).
- Sony. 2048-Pixel CCD Linear Sensor (B/W) for Single 5V Power Supply Bar-code Reader ILX511B. Available online: https://oceanoptics.com/wp-content/uploads/SONY-ILX511B.pdf (accessed on 3 May 2018).
- Brach, E.J.; Molnar, J.M.; Jasmin, J.J. Detection of lettuce maturity and variety by remote sensing techniques. J. Agric. Eng. Res. 1977, 22, 45–54. [Google Scholar] [CrossRef]
- Buschmann, C. Variability and application of the chlorophyll fluorescence emission ratio red/far red of leaves. Photosynth. Res. 2007, 92, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Rinderle, U. The role of chlorophyll fluorescence in the detection of stress conditions in plants. CRC Crit. Rev. Anal. Chem. 1988, 19, S29–S85. [Google Scholar] [CrossRef]
- McMurtrey, J.E., III; Chappelle, E.W.; Kim, M.S.; Meisinger, J.J.; Corp, L.A. Distinguishing nitrogen fertilization levels in field corn (Zea mays L.) with actively induced fluorescence and passive reflectance measurements. Remote Sens. Environ. 1994, 47, 36–44. [Google Scholar] [CrossRef]
- Subhash, N.; Mohanan, C.N. Curve fit analysis of chlorophyll fluorescence spectra: Application to nutrient stress detection in sunflower. Remote Sens. Environ. 1997, 60, 347–356. [Google Scholar] [CrossRef]
- Schuerger, A.C.; Capelle, G.A.; Di Benedetto, J.A.; Mao, C.; Thai, C.M.; Evans, M.D.; Richards, J.T.; Blank, T.A.; Stryjewski, E.C. Comparison of two hyperspectral imaging and two laser induced fluorescence instruments for the detection of zinc stress and chlorophyll concentration in Bahia grass (Paspalum notatum Flugge). Remote Sens. Environ. 2003, 84, 572–588. [Google Scholar] [CrossRef]
- Chappelle, E.W.; Wood, F.M.; McMurtrey, J.E., Jr.; Newcomb, W.W. Laser induced fluorescence of green plants. 1: A technique for the remote detection of plant stress and species differentiation. Appl. Opt. 1984, 23, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Hák, R.; Lichtenthaler, H.K.; Rinderle, U. Decrease of the chlorophyll fluorescence ratio F690/F730 during greening and development of leaves. Radiat. Environ. Biophys. 1990, 29, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Hák, R.; Rinderle, U. The chlorophyll fluorescence ratioF690/F730in leaves of different chlorophyll contents. Photosynth. Res. 1990, 25, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Lavrov, A.; Utkin, A.B.; Marques da Silva, J.; Vilar, R.; Santos, N.M.; Alves, B. Water stress assessment of cork oak leaves and maritime pine needles based on LIF spectra. Opt. Spectrosc. 2012, 112, 271–279. [Google Scholar] [CrossRef]
- Cerovic, Z.G.; Goulas, Y.; Gorbunov, M.; Briantais, J.-M.; Camenen, L.; Moya, I. Fluorosensing of water stress in plants: Diurnal changes of the mean lifetime and yield of chlorophyll fluorescence, measured simultaneously and at distance with a LIDAR and a modified PAM-fluorimeter, in maize, sugar beet, and kalanchoë. Remote Sens. Environ. 1996, 58, 311–321. [Google Scholar] [CrossRef]
- Kolber, Z.; Klimov, D.; Ananyev, G.; Rascher, U.; Berry, J.; Osmond, B. Measuring photosynthetic parameters at a distance: Laser induced fluorescence transient (LIFT) method for remote measurements of photosynthesis in terrestrial vegetation. Photosynth. Res. 2005, 84, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, C.; Utkin, A.B.; Cartaxana, P.; Marques da Silva, J.; Matos, A.R. The use of laser induced chlorophyll fluorescence (LIF) as a fast and non-destructive method to investigate water deficit in Arabidopsis. Agric. Water Manag. 2016, 164, 127–136. [Google Scholar] [CrossRef]
- Macintyre, H.L.; Geider, R.J.; Miller, D.C. Microphytobenthos: The ecological role of the “secret garden” of unvegetated, shallow-water marine habitats. I. Distribution, abundance and primary production. Estuaries 1996, 19, 186–201. [Google Scholar] [CrossRef]
- Paterson, D.M.; Black, K.M. Water flow, sediment dynamics and benthic biology. Adv. Ecol. Res. 1999, 29, 155–193. [Google Scholar]
- Eldridge, D.J.; Leys, J.F. Exploring some relationships between biological soil crusts, soil aggregation and wind erosion. J. Arid Environ. 2003, 53, 457–466. [Google Scholar] [CrossRef]
- Adessi, A.; Cruz de Carvalho, R.; De Philippis, R.; Branquinho, C.; Marques da Silva, J. Microbial extracellular polymeric substances improve water retention in dryland biological soil crusts. Soil Biol. Biochem. 2018, 116, 67–69. [Google Scholar] [CrossRef]
- Nagarkar, S.; Williams, G.A. Comparative techniques to quantify cyanobacteria dominated epilithic biofilms on tropical rocky shores. Mar. Ecol. Prog. Ser. 1997, 154, 281–291. [Google Scholar] [CrossRef]
- Round, F.E.; Palmer, J.D. Persistent, vertical-migration rhythms in benthic microflora. II. Field and laboratory studies on diatoms from the banks of the river Avon. J. Mar. Biol. Assoc. UK 1966, 46, 191–214. [Google Scholar] [CrossRef]
- Paterson, D.M. The migratory behaviour of diatom assemblages in a laboratory tidal micro-ecosystem examined by low temperature scanning electron microscopy. Diatom Res. 1986, 1, 227–239. [Google Scholar] [CrossRef]
- Marques da Silva, J.; Cruz, S.; Cartaxana, P. Inorganic carbon availability in benthic diatom communities: Photosynthesis and migration. Philos. Trans. R. Soc. B 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Middelburg, J.J.; Barranguet, C.; Boschker, H.T.S.; Herman, P.M.J.; Moens, T.; Heip, C.H.R. The fate of intertidal microphytobenthos carbon: An in situ 13C-labeling study. Limnol. Oceanogr. 2000, 45, 1224–1234. [Google Scholar] [CrossRef]
- Marques da Silva, J. Reconciling science and nature by means of the aesthetical contemplation of natural diversity. Rivista di Estetica 2015, 59, 93–113. [Google Scholar] [CrossRef]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farberk, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Belnap, J.; Kaltenecker, J.H.; Rosentreter, R.; Williams, J.; Leonard, S.; Eldridge, D. Biological Soil Crusts: Ecology and Management; Technical Reference 1730-2; Paterson, P., Ed.; United States Department of Interior: Denver, CO, USA, 2001.
- Bartrons, M.; Catalan, J.; Casamayor, E.O. High bacterial diversity in epilithic biofilms of oligotrophic mountain lakes. Microb. Ecol. 2012, 64, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; McBain, A.J.; Stead, A.T.; Gilbert, P. Shear rate moderates community diversity in freshwater biofilms. Appl. Environ. Microbiol. 2004, 70, 7426–7435. [Google Scholar] [CrossRef] [PubMed]
- Besemer, K. Biodiversity, community structure and function of biofilms in stream ecosystems. Res. Microbiol. 2015, 166, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Cutler, N.A.; Chaput, D.L.; Oliver, A.E.; Viles, H.A. The spatial organization and microbial community structure of an epilithic biofilm. FEMS Microbiol. Ecol. 2015, 93, fiu027. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, A.; Li, X.; Osuga, Y.; Kawashima, A.; Gu, J.-D.; Nasu, M.; Katayama, Y. Bacterial communities in pigmented biofilms formed on the swandstone bas-relief walls of the Bayon temple, Angkor Thom, Cambodia. Microbes Environ. 2013, 28, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Zimermann, J.; Gonzalez, J.M.; Saiz-Jimenez, C. Epilithic biofilms in Saint Callixtus Catacombs (Rome) harbour a broad spectrum of Acidobacteria. Antonie van Leeuwenhoek 2006, 89, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Ereshefsky, M.; Pedroso, M. Rethinking evolutionary individuality. Proc. Natl. Acad. Sci. USA 2015, 112, 10126–10132. [Google Scholar] [CrossRef] [PubMed]
- Marques da Silva, J.; Casetta, E. Why a species-based approach to biodiversity is not enough. Lessons from multispecies biofilms. In From Assessing to Conserving Biodiversity; Casetta, E., Vecchi, D., Marques da Silva, J., Eds.; Springer: Dordrecht, The Netherlands, in press.
- Kromkamp, J.; Barranguet, C.; Peene, J. Determination of microphytobenthos PSII quantum efficiency and photosynthetic activity by means of variable chlorophyll fluorescence. Mar. Ecol. Prog. Ser. 1998, 162, 45–55. [Google Scholar] [CrossRef]
- Richards, J.T.; Schuerger, A.C.; Capelle, G.; Guikema, J.A. Laser-induced fluorescence spectroscopy of dark and light-adapted bean (Phaseolus vulgaris L.) and wheat (Triticum aestivum L.) plants grown under three irradiance levels and subjected to fluctuating lighting conditions. Remote Sens. Environ. 2003, 84, 323–341. [Google Scholar] [CrossRef]
- Anderson, B.; Buah-Bassuah, P.; Tetteh, J. Using violet laser-induced chlorophyll fluorescence emission spectra for crop yield assessment of cowpea (Vigna unguiculate (L) Walp) varieties. Meas. Sci. Technol. 2004, 15, 1255–1265. [Google Scholar] [CrossRef]
- Ananyev, G.; Kolber, Z.S.; Klimov, D.; Falkowski, P.G.; Berry, J.A.; Raschers, U.; Martin, R.; Osmond, B. Remote sensing of heterogeneity in photosynthetic efficiency, electron transport and dissipation of excess light in Populus deltoides stands under ambient and elevated CO2 concentrations, and in a tropical forest canopy, using a new laser-induced fluorescence transient device. Glob. Chang. Biol. 2005, 11, 1195–1206. [Google Scholar] [CrossRef]
- Pieruschka, R.; Klimov, D.; Kolber, Z.; Berry, J.A. Monitoring of cold and light stress impact on photosynthesis by using laser induced fluorescence transient (LIFT) approach. Funct. Plant Biol. 2010, 37, 395–402. [Google Scholar] [CrossRef]
- Vieira, S. Photosynthetic Metabolism in Microphytobenthos: Modulation by Temperature and Dissolved Inorganic Carbon. Ph.D. Thesis, University of Lisbon, Lisbon, Portugal, 2016. [Google Scholar]
- Vieira, S.; Lavrov, A.; Utkin, A.; Santos, A.; Vilar, R.; Marques da Silva, J.; Cartaxana, P. Effects of intertidal microphytobenthos migration on biomass determination via laser-induced fluorescence. Mar. Ecol. Prog. Ser. 2011, 432, 45–52. [Google Scholar] [CrossRef]
- Launeau, P.; Méléder, V.; Verpoorter, C.; Barillé, L.; Kazemipour-Ricci, F.; Giraud, M.; Jesus, B.; Le Menn, E. Microphytobenthos biomass and diversity mapping at different spatial scales with a hyperspectral optical model. Remote Sens. 2018, 10, 716. [Google Scholar] [CrossRef]
- Myneni, R.B.; Hall, F.G.; Sellers, P.G.; Marshak, A.L. The interpretation of spectral vegetation indexes. IEEE Trans. Geosci. Remote Sens. 1995, 33, 481–486. [Google Scholar] [CrossRef]
- Brito, A.C.; Benyoucef, I.; Jesus, B.; Brotas, V.; Gernez, P.; Mendes, C.R.; Launeau, P.; Peixe Dias, M.; Barillé, L. Seasonality of microphytobenthos revealed by remote-sensing in a South European estuary. Cont. Shelf Res. 2013, 66, 83–91. [Google Scholar] [CrossRef]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic pigments in diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef] [PubMed]
- Pinevich, A.V.; Averina, S.G.; Velichko, N.V. Another view on the role of photosynthetic pigments in taxonomy of oxygenic-phototrophic bacteria: Proposed rejection of the order Prochlorales Florenzano, Balloni, and Materassi 1986 (Emend. Burger-Wiersma, Stal, and Mur 1989), the family Prochloraceae Florenzano, Balloni, and Materassi 1986, and the family Prochlorotrichaceae Burger-Wiersma, Stal, and Mur 1989. Int. J. Syst. Bacteriol. 1997, 47, 1264–1267. [Google Scholar]
- Beutler, M. Spectral Fluorescence of Chlorophyll and Phycobilins as an In-Situ Tool of Phytoplankton Analysis—Models, Algorithms and Instruments. Ph.D. Thesis, Christian-Albrechts-Universität zu Kiel, Kiel, Germany, 2003. [Google Scholar]
- Quantel. Ultra (50–100 mJ) Lamp Pumped Solid State Laser. Available online: http://www.quantel-laser.com/en/products/item/ultra-50-100-mj--134.html (accessed on 12 September 2018).
- Van der Weij-De Wit, C.D.; Doust, A.B.; van Stokkum, I.H.M.; Dekker, J.P.; Wilk, K.E.; Curmi, P.M.G.; Scholes, G.D.; van Grondelle, R. How energy frunnels from the phycoerythrin antenna complex to photosystem I and photosystem II in cryptophyte Rhodomonas CS24 cells. J. Phys. Chem. B 2006, 110, 25066–25073. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, S.A.; Sauer, K. Spectroscopic study of the light-harvesting protein C-phycocyanin associated with colorless linker peptides. Photochem. Photobiol. 2001, 73, 556–563. [Google Scholar] [CrossRef]
- Raanan, H.; Oren, N.; Treves, H.; Keren, N.; Ohad, I.; Berkowicz, S.M.; Hagemann, M.; Koch, M.; Shotland, Y.; Kaplan, A. Towards clarifying what distinguishes cyanobacteria able to resurrect after desiccation from those that cannot: The photosynthetic aspect. Biochim. Biophys. Acta 2016, 1857, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Limão, F.M.; Maciel, M.J. Tróia. Wiley Online Library. Available online: https://doi.org/10.1002/9781444338386.wbeah30227 (accessed on 21 March 2018).
- Solhaug, K.A.; Gauslaa, Y.; Nybakken, L.; Bilger, W. UV-induction of sun-screening pigments in lichens. New Phytol. 2003, 158, 91–100. [Google Scholar] [CrossRef]
- Thorne, K.M.; Elliott-Fisk, D.L.; Freeman, C.M.; Bui, T.-V.D.; Powelson, K.W.; Janousek, C.N.; Buffington, K.J.; Takekawa, J.Y. Are coastal managers ready for climate change? A case study from estuaries along the Pacific coast of the United States. Ocean Coast. Manag. 2017, 143, 38–50. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2016, 6, 166–171. [Google Scholar] [CrossRef]
- Chamizo, S.; Mugnai, G.; Rossi, F.; Certini, G.; De Philippis, R. Cyanobacteria inoculation improves soil stability and fertility on different textured soils: Gaining insights for applicability in soil restoration. Front. Environ. Sci. 2018, 6, 49. [Google Scholar] [CrossRef]
- Joiner, J.; Yoshida, Y.; Vasilkov, A.P.; Yoshida, Y.; Corp, L.A.; Middleton, E.M. First observations of global and seasonal terrestrial chlorophyll fluorescence from space. Biogeosciences 2011, 8, 637–651. [Google Scholar] [CrossRef]
- Cartaxana, P.; Vieira, S.; Ribeiro, L.; Rocha, R.; Cruz, S.; Calado, R.; Marques da Silva, J. Effects of elevated temperature and CO2 on intertidal microphytobenthos. BMC Ecol. 2015, 15, 10. [Google Scholar] [CrossRef] [PubMed]
- Escolar, C.; Martínez, I.; Bowker, M.A.; Maestre, F.T. Warming reduces the growth and diversity of biological soil crusts in a semi-arid environment: Implications for ecosystem structure and functioning. Philos. Trans. R. Soc. B 2012, 367, 3087–3099. [Google Scholar] [CrossRef] [PubMed]
- LDI Innovation. Spectral Fluorescence Signatures. Available online: http://ldi-innovation.eu/documents/SFStechnique.pdf (accessed on 12 September 2018).
- STORM Consortium. Early Detection of Biological Infestation: Experiments Started at Tróia. Available online: http://www.storm-project.eu/early-detection-of-biological-infestation-experiments-started-at-troia/ (accessed on 3 May 2018).
- Utkin, A.B.; Chaves, P.; Fernandes, L.; Pinto, I.V.; Revez, M.J. LIF and SFS techniques for early detection of biofilms harmful for cultural heritage. In Proceedings of the 2018 International Conference Laser Optics (ICLO), St. Petersburg, Russia, 4–8 June 2018; p. 275. [Google Scholar] [CrossRef]
- Strieth, D.; Ulber, R.; Muffler, K. Application of phototrophic biofilms: From fundamentals to processes. Bioprocess Biosyst. Eng. 2018, 41, 295–312. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.J.; Bombelli, P.; Scott, A.M.; Philips, A.J.; Smith, A.G.; Fisher, A.C.; Howe, C.J. Photosynthetic biofilms in pure culture harness solar energy in a mediatorless bio-photovoltaic cell (BPV) system. Energy Environ. Sci. 2011. [Google Scholar] [CrossRef]
- Vieira, S.; Cartaxana, P.; Máguas, C.; Marques da Silva, J. Photosynthesis in estuarine intertidal microphytobenthos is limited by inorganic carbon availability. Photosynth. Res. 2016, 128, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Podola, B.; de Beer, D.; Melkonian, M. A method to determine photosynthetic activity from oxygen microsensor data in biofilms subjected to evaporation. J. Microbiol. Methods 2015, 117, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Roeselers, G.; van Loosdrecht, M.C.M.; Muyzer, G. Phototrophic biofilms and their potential applications. J. Appl. Phycol. 2008, 20, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Kohl, A.; Fillardet, T.; Moisan, H.; Brousse, E. Progress of laser diode arrays operating under harsh conditions. Proc. SPIE 2010, 7835, 78350Q. [Google Scholar] [CrossRef]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques da Silva, J.; Borissovitch Utkin, A. Application of Laser-Induced Fluorescence in Functional Studies of Photosynthetic Biofilms. Processes 2018, 6, 227. https://doi.org/10.3390/pr6110227
Marques da Silva J, Borissovitch Utkin A. Application of Laser-Induced Fluorescence in Functional Studies of Photosynthetic Biofilms. Processes. 2018; 6(11):227. https://doi.org/10.3390/pr6110227
Chicago/Turabian StyleMarques da Silva, Jorge, and Andrei Borissovitch Utkin. 2018. "Application of Laser-Induced Fluorescence in Functional Studies of Photosynthetic Biofilms" Processes 6, no. 11: 227. https://doi.org/10.3390/pr6110227
APA StyleMarques da Silva, J., & Borissovitch Utkin, A. (2018). Application of Laser-Induced Fluorescence in Functional Studies of Photosynthetic Biofilms. Processes, 6(11), 227. https://doi.org/10.3390/pr6110227