

Rapeseed Meal Waste Biomass as a Single-Cell Protein Substrate for Nutritionally-Enhanced Feed Components

 , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Waste Material and Sample Preparation
2.2. Enzymatic Pre-Hydrolysis and Simultaneous Saccharification and Fermentation (SSF)
2.3. Determination of the Protein Content
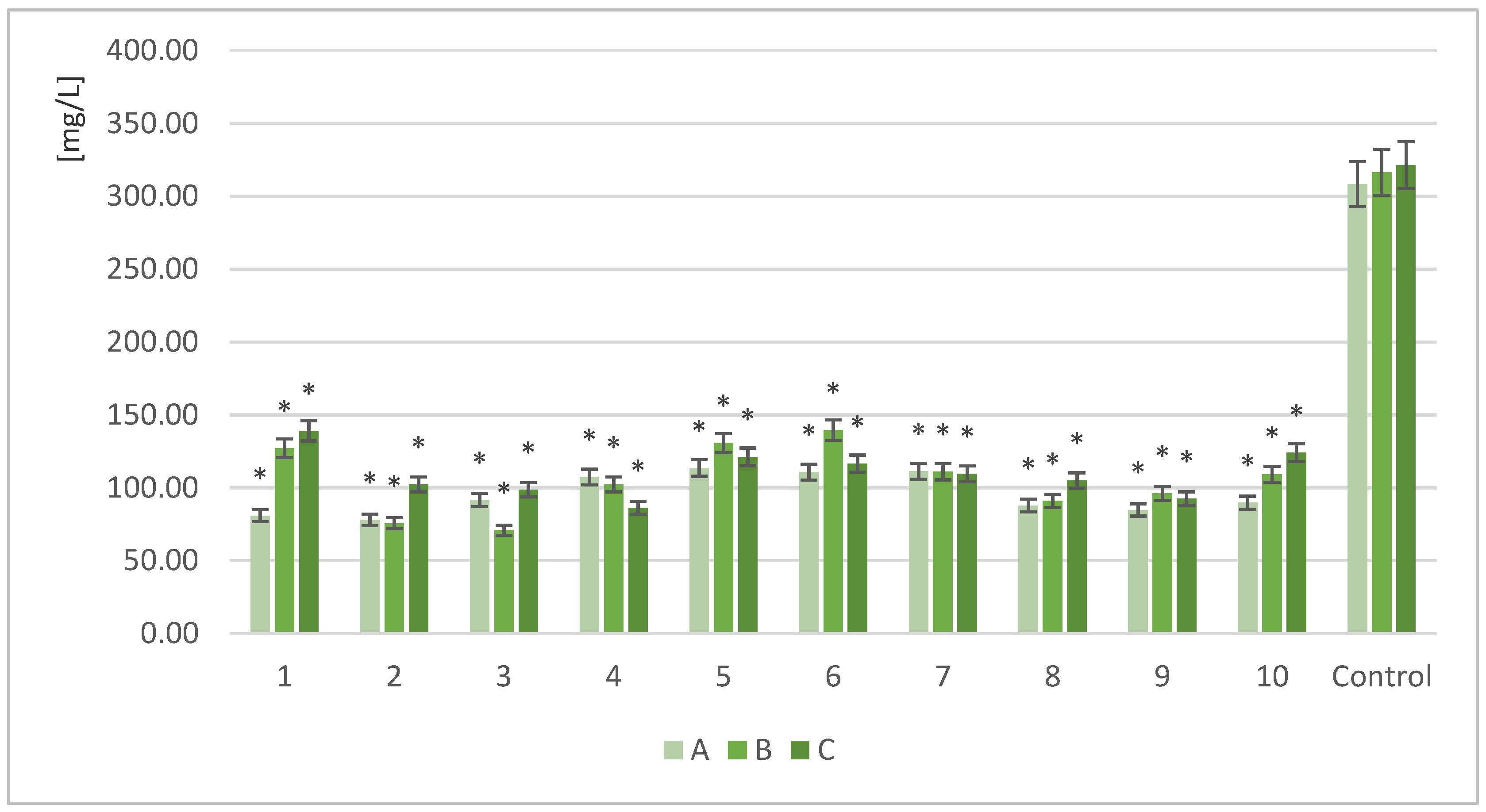
2.4. Determination of the Free Nitrogen (FAN)
2.5. Determination of the Crude Fiber (CF) Content
2.6. Chromatographic Analysis of the Flavonoids
2.7. Statistical Analysis
3. Results
3.1. Influence of Different Biomass Loads on Various Parameters
3.1.1. Protein Content Increase with Various Biomass Loads
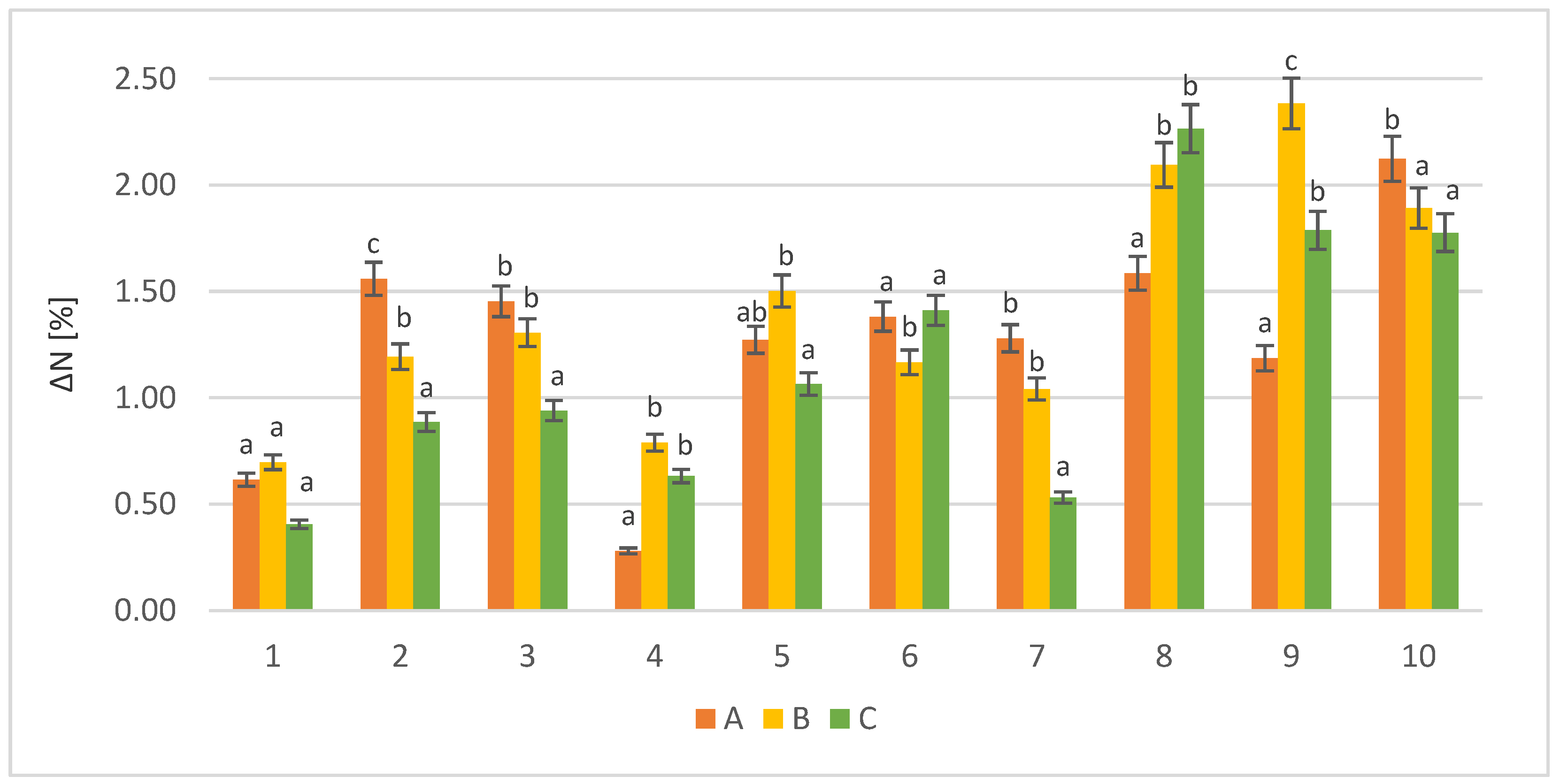
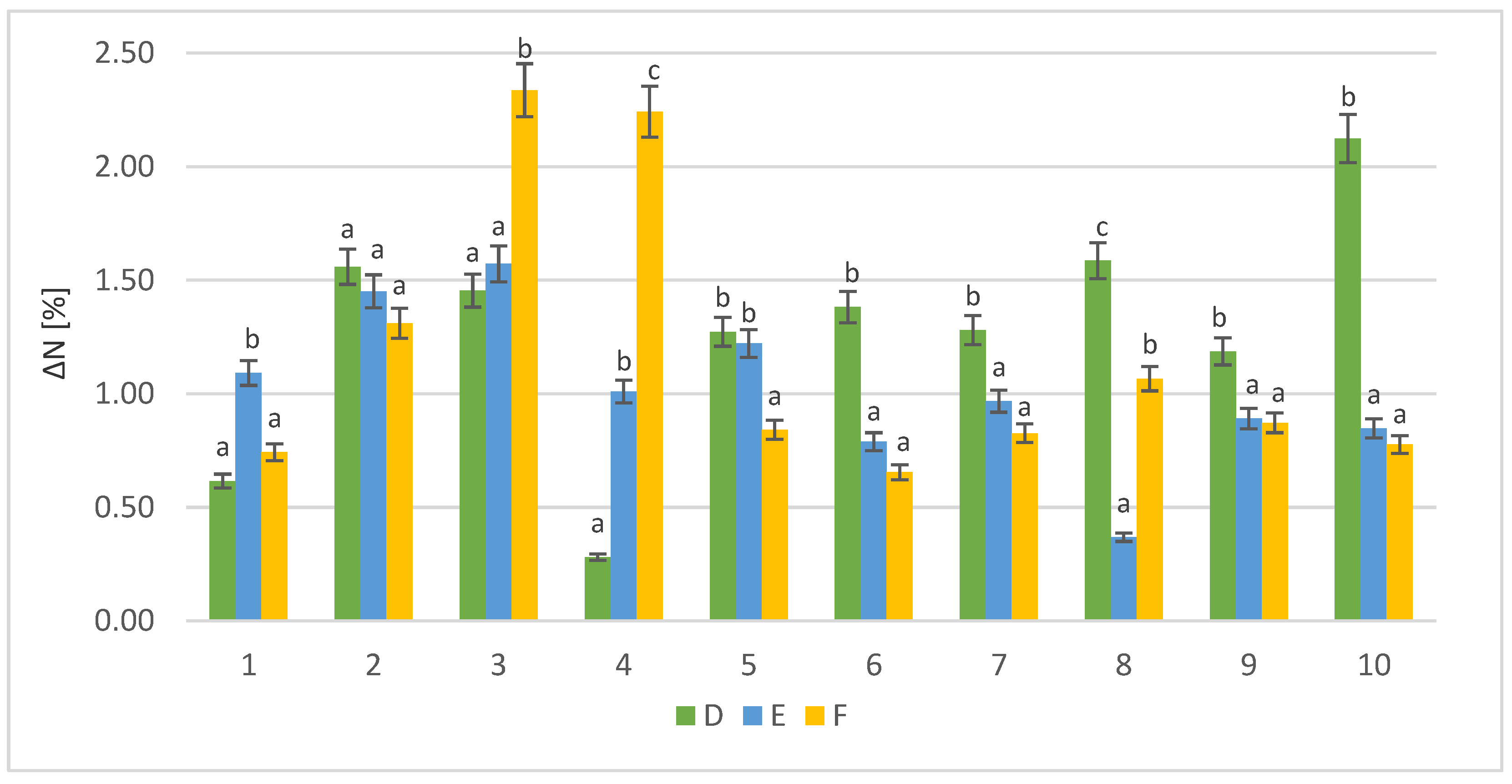
3.1.2. Free Amino Nitrogen Content in Sample of Post-Culture Liquid
3.2. Influence of Different Doses of Enzymatic Preparation on Production of Feeds
3.2.1. Measurement of Yeast Growth by the Plate Count Method
3.2.2. Protein Content under Different Doses of Enzymatic Preparation
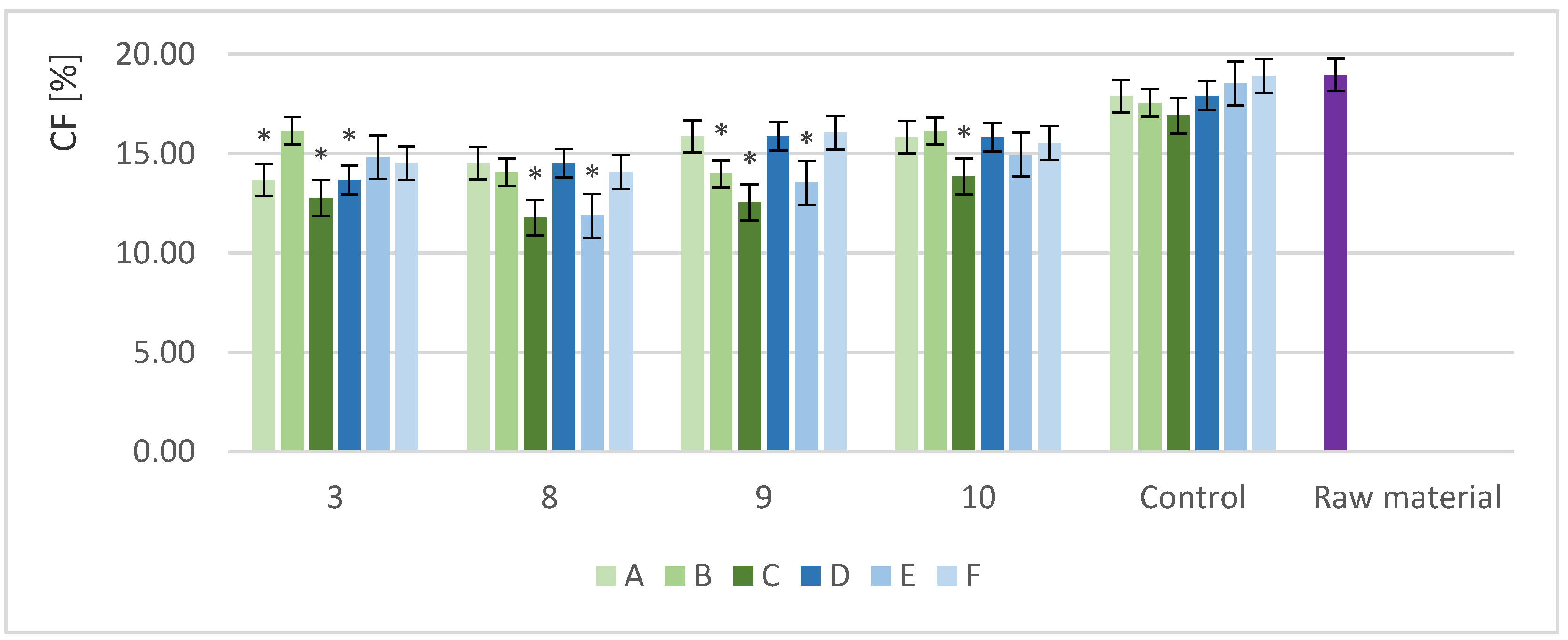
3.2.3. Crude Fiber Content in Samples with Different Biomass Loads and Enzyme Doses
3.2.4. Flavonoid Content after Fermentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahbandeh, M. Rapeseed Oil: Global Production Volume 2012/13–2022/23; Statista: Hamburg, Germany, 2023; Available online: https://www.statista.com/statistics/613487/rapeseed-oil-production-volume-worldwide/ (accessed on 3 April 2023).
- Lemaire, A.; Limbourg, S. How Can Food Loss and Waste Management Achieve Sustainable Development Goals? J. Clean. Prod. 2019, 234, 1221–1234. [Google Scholar] [CrossRef]
- Zentrale Markt-und Preisinformationen Rapeseed Meal Current Stock Exchange Prices & Charts Rapeseed Meal. Available online: https://www.zmp.de/en/oilseeds/rapeseed-meal_future (accessed on 3 April 2023).
- Mattila, P.; Pap, N.; Järvenpää, E.; Kahala, M.; Mäkinen, S. Underutilized Northern Plant Sources and Technological Aspects for Recovering Their Polyphenols. Adv. Food Nutr. Res. 2021, 98, 125–169. [Google Scholar] [PubMed]
- De Corato, U.; Viola, E. Biofuel Co-Products for Livestock Feed. In Agricultural Bioeconomy; Academic Press: Cambridge, MA, USA, 2023; pp. 245–286. [Google Scholar]
- Rafii, F. The Role of Colonic Bacteria in the Metabolism of the Natural Isoflavone Daidzin to Equol. Metabolites 2015, 5, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-Y.; Ye, Y.; Xiao, L.; Rahman, K.; Xia, W.; Zhang, H. Daidzein: A Review of Pharmacological Effects. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 117. [Google Scholar] [CrossRef]
- Ahmed, T.; Javed, S.; Tariq, A.; Budzyńska, B.; D’Onofrio, G.; Daglia, M.; Fazel Nabavi, S.; Mohammad Nabavi, S. Daidzein and Its Effects on Brain. Curr. Med. Chem. 2017, 24, 365–375. [Google Scholar] [CrossRef]
- Tropea, A.; Ferracane, A.; Albergamo, A.; Potortì, A.G.; Lo Turco, V.; Di Bella, G. Single Cell Protein Production through Multi Food-Waste Substrate Fermentation. Fermentation 2022, 8, 91. [Google Scholar] [CrossRef]
- Gong, Y.; Liu, J.; Jiang, M.; Liang, Z.; Jin, H.; Hu, X.; Wan, X.; Hu, C. Improvement of Omega-3 Docosahexaenoic Acid Production by Marine Dinoflagellate Crypthecodinium Cohnii Using Rapeseed Meal Hydrolysate and Waste Molasses as Feedstock. PLoS ONE 2015, 10, e0125368. [Google Scholar] [CrossRef]
- Lomascolo, A.; Uzan-Boukhris, E.; Sigoillot, J.C.; Fine, F. Rapeseed and Sunflower Meal: A Review on Biotechnology Status and Challenges. Appl. Microbiol. Biotechnol. 2012, 95, 1105–1114. [Google Scholar] [CrossRef]
- Yusuf, H.A.; Piao, M.; Ma, T.; Huo, R.; Tu, Y. Effect of Lactic Acid Bacteria and Yeast Supplementation on Anti-Nutritional Factors and Chemical Composition of Fermented Total Mixed Ration Containing Cottonseed Meal or Rapeseed Meal. Anim. Biosci. 2022, 35, 556–566. [Google Scholar] [CrossRef]
- Vlassa, M.; Filip, M.; Țăranu, I.; Marin, D.; Untea, A.E.; Ropotă, M.; Dragomir, C.; Sărăcilă, M. The Yeast Fermentation Effect on Content of Bioactive, Nutritional and Anti-Nutritional Factors in Rapeseed Meal. Foods 2022, 11, 2972. [Google Scholar] [CrossRef]
- Bau, H.-M.; Villaume, C.; Lin, C.-F.; Evrard, J.; Quemener, B.; Nicolas, J.-P.; Méjean, L. Effect of a Solid-state Fermentation Using Rhizopus Oligosporus Sp.T-3 on Elimination of Antinutritional Substances and Modification of Biochemical Constituents of Defatted Rapeseed Meal. J. Sci. Food Agric. 1994, 65, 315–322. [Google Scholar] [CrossRef]
- Mandiki, S.N.M.; Derycke, G.; Bister, J.L.; Mabon, N.; Wathelet, J.P.; Marlier, M.; Paquay, R. Chemical Changes and Influences of Rapeseed Antinutritional Factors on Gestating and Lactating Ewes 1. Animal Performances and Plasma Hormones and Glucose. Anim. Feed Sci. Technol. 2002, 98, 25–35. [Google Scholar] [CrossRef]
- Dygas, D.; Janicka, P.; Berłowska, J.; Kręgiel, D. Conventional and Unconventional Yeasts Able to Grow on Rapeseed Meal Hydrolysates. BioResources 2022, 17, 3082–3094. [Google Scholar] [CrossRef]
- Geisler, J.; Weiß, N. Free Amino Nitrogen (FAN) Measurement in Beer. Short Protoc. 2015, 9, 10–12. [Google Scholar]
- ISO 6865:2000; Animal Feeding Stuffs—Determination of Crude Fibre Content—Method with Intermediate Filtration. ISO: Geneva, Switzerland, 2000; p. 10.
- Sulyok, M.; Stadler, D.; Steiner, D.; Krska, R. Validation of an LC-MS/MS-Based Dilute-and-Shoot Approach for the Quantification of > 500 Mycotoxins and Other Secondary Metabolites in Food Crops: Challenges and Solutions. Anal. Bioanal. Chem. 2020, 412, 2607–2620. [Google Scholar] [CrossRef]
- Steglińska, A.; Sulyok, M.; Janas, R.; Grzesik, M.; Liszkowska, W.; Kręgiel, D.; Gutarowska, B. Metabolite Formation by Fungal Pathogens of Potatoes (Solanum tuberosum L.) in the Presence of Bioprotective Agents. Int. J. Environ. Res. Public Health 2023, 20, 5221. [Google Scholar] [CrossRef]
- Bell, J.M. Nutrients and Toxicants in Rapeseed Meal: A Review. J. Anim. Sci. 1984, 58, 996–1010. [Google Scholar] [CrossRef]
- Yang, Z.; Huang, Z.; Cao, L. Biotransformation technology and high-value application of rapeseed meal: A review. Bioresour. Bioprocess. 2022, 9, 103. [Google Scholar] [CrossRef]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.-S.; Hatziloukas, E.; Parapouli, M.; Vasileiadis, A.; Afendra, A.-S.; Hatziloukas, E. Saccharomyces Cerevisiae and Its Industrial Applications. AIMS Microbiol. 2020, 6, 1–31. [Google Scholar] [CrossRef]
- Liu, H.H.; Ji, X.J.; Huang, H. Biotechnological Applications of Yarrowia Lipolytica: Past, Present and Future. Biotechnol. Adv. 2015, 33, 1522–1546. [Google Scholar] [CrossRef]
- Postma, E.; Kuiper, A.; Tomasouw, W.F.; Scheffers, W.A.; Van Dijken, J.P. Competition for Glucose between the Yeasts Saccharomyces Cerevisiae and Candida Utilis. Appl. Environ. Microbiol. 1989, 55, 3214–3220. [Google Scholar] [CrossRef]
- Carranza-Méndez, R.C.; Chávez-González, M.L.; Sepúlveda-Torre, L.; Aguilar, C.N.; Govea-Salas, M.; Ramos-González, R. Production of Single Cell Protein from Orange Peel Residues by Candida Utilis. Biocatal. Agric. Biotechnol. 2022, 40, 102298. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol Yeasts: Mechanisms and Applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, H.; Han, B.; Li, H.Y.; Liu, X.L. Improvement of Nutritional Value, Molecular Weight Patterns (Soluble Peptides), Free Amino Acid Patterns, Total Phenolics and Antioxidant Activity of Fermented Extrusion Pretreatment Rapeseed Meal with Bacillus subtilis YY-1 and Saccharomyces cerevisiae Y. LWT 2022, 160, 113280. [Google Scholar] [CrossRef]
- Bertolo, A.P.; Biz, A.P.; Kempka, A.P.; Rigo, E.; Cavalheiro, D. Yeast (Saccharomyces cerevisiae): Evaluation of Cellular Disruption Processes, Chemical Composition, Functional Properties and Digestibility. J. Food Sci. Technol. 2019, 56, 3697–3706. [Google Scholar] [CrossRef]
- Uçkun Kiran, E.; Salakkam, A.; Trzcinski, A.P.; Bakir, U.; Webb, C. Enhancing the Value of Nitrogen from Rapeseed Meal for Microbial Oil Production. Enzyme Microb. Technol. 2012, 50, 337–342. [Google Scholar] [CrossRef]
- Poulsen, H.D.; Blaabjerg, K. Fermentation of rapeseed meal, sunflower meal and faba beans in combination with wheat bran increases solubility of protein and phosphorus. J. Sci. Food Agric. 2017, 97, 244–251. [Google Scholar] [CrossRef]
- Jeong, H.S.; Kim, H.Y.; Ahn, S.H.; Oh, S.C.; Yang, I.; Choi, I.G. Optimization of Enzymatic Hydrolysis Conditions for Extraction of Pectin from Rapeseed Cake (Brassica napus L.) Using Commercial Enzymes. Food Chem. 2014, 157, 332–338. [Google Scholar] [CrossRef]
- Dygas, D.; Nowak, S.; Olszewska, J.; Szymańska, M.; Mroczyńska-Florczak, M.; Berłowska, J.; Dziugan, P.; Kręgiel, D. Ability of Yeast Metabolic Activity to Reduce Sugars and Stabilize Betalains in Red Beet Juice. Fermentation 2021, 7, 105. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, L.; Zhang, Z.; Ding, L.; Hang, S. Combination of Fiber-Degrading Enzymatic Hydrolysis and Lactobacilli Fermentation Enhances Utilization of Fiber and Protein in Rapeseed Meal as Revealed in Simulated Pig Digestion and Fermentation in Vitro. Anim. Feed. Sci. Technol. 2021, 278, 115001. [Google Scholar] [CrossRef]
- Ramonet, Y.; Meunier-Salaün, M.C.; Dourmad, J.Y. High-Fiber Diets in Pregnant Sows: Digestive Utilization and Effects on the Behavior of the Animals. J. Anim. Sci. 1999, 77, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Guillemet, R.; Hamard, A.; Quesnel, H.; Père, M.C.; Etienne, M.; Dourmad, J.Y.; Meunier-Salaün, M.C. Dietary Fibre for Gestating Sows: Effects on Parturition Progress, Behaviour, Litter and Sow Performance. Animal 2007, 1, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Grigorev, M.; Grigoreva, A.; Sharvadze, R.; Chernogradskaya, N.; Stepanova, S. The Effectiveness of Unconventional Feed Additives at Feeding Cattle in Conditions Yakutia. Lect. Notes Netw. Syst. 2023, 574, 156–166. [Google Scholar] [CrossRef]
- Grgic, D.; Varga, E.; Novak, B.; Müller, A.; Marko, D. Isoflavones in Animals: Metabolism and Effects in Livestock and Occurrence in Feed. Toxins 2021, 13, 836. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, X.; Cai, L.; Zhang, Y.; Ding, H.; Yin, J.; Li, X. Effects of Daidzein on Antioxidant Capacity in Weaned Pigs and IPEC-J2 Cells. Anim. Nutr. 2022, 11, 48–59. [Google Scholar] [CrossRef]
- Li, Y.P.; Jiang, X.R.; Wei, Z.X.; Cai, L.; Yin, J.D.; Li, X.L. Effects of Soybean Isoflavones on the Growth Performance, Intestinal Morphology and Antioxidative Properties in Pigs. Animal 2020, 14, 2262–2270. [Google Scholar] [CrossRef]
- Pan, W.; Ikeda, K.; Takebe, M.; Yamori, Y. Genistein, Daidzein and Glycitein Inhibit Growth and DNA Synthesis of Aortic Smooth Muscle Cells from Stroke-Prone Spontaneously Hypertensive Rats. J. Nutr. 2001, 131, 1154–1158. [Google Scholar] [CrossRef]
- Hu, Y.J.; Gao, K.G.; Zheng, C.T.; Wu, Z.J.; Yang, X.F.; Wang, L.; Ma, X.Y.; Zhou, A.G.; Jiang, Z.J. Effect of Dietary Supplementation with Glycitein during Late Pregnancy and Lactation on Antioxidative Indices and Performance of Primiparous Sows. J. Anim. Sci. 2015, 93, 2246–2254. [Google Scholar] [CrossRef]
- Smith, B.N.; Dilger, R.N. Immunomodulatory Potential of Dietary Soybean-Derived Isoflavones and Saponins in Pigs. J. Anim. Sci. 2018, 96, 1288–1304. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Yeast | Collection Name |
|---|---|---|
| 1 | Yarrowia lipolytica | ŁOCK 0264 |
| 2 | Metschnikowia pulcherrima | NCYC 747 |
| 3 | Scheffersomyces stipitis | NCYC 1541 |
| 4 | Kluyveromyces marxianus | ŁOCK 0024 |
| 5 | Candida humicola | ŁOCK 0013 |
| 6 | Saccharomyces bayanus | BC S103 |
| 7 | Candida utilis | ŁOCK 0021 |
| 8 | Saccharomyces cerevisiae TT | ŁOCK 0105 |
| 9 | Saccharomyces cerevisiae Ethanol Red | Leaf/Lesaffre Advanced Fermentation |
| 10 | Saccharomyces cerevisiae Tokay | ŁOCK 0204 |
| Strain | D | E | F | Control Sample |
|---|---|---|---|---|
| 1 | AV: 4.92 × 108 | AV: 1.38 × 108 | AV: 1.64 × 108 | AV: 6.02 × 105 |
| SD: 2.03 × 108 | SD: 7.69 × 107 | SD: 4.51 × 107 | SD: 8.61 × 104 | |
| 2 | AV: 1.64 × 108 | AV: 1.26 × 108 | AV: 9.32 × 107 | AV: 6.18 × 105 |
| SD: 8.14 × 107 | SD: 2.07 × 107 | SD: 4.62 × 107 | SD: 1.09 × 105 | |
| 3 | AV: 2.50 × 108 | AV: 2.30 × 108 | AV: 2.32 × 108 | AV: 8.50 × 105 |
| SD: 1.50 × 108 | SD: 1.53 × 108 | SD: 1.46 × 108 | SD: 5.83 × 104 | |
| 4 | AV: 2.20 × 107 | AV: 1.05 × 107 | AV: 9.06 × 106 | AV: 1.56 × 105 |
| SD: 6.44 × 106 | SD: 1.45 × 106 | SD: 7.33 × 105 | SD: 5.35 × 104 | |
| 5 | AV: 3.82 × 108 | AV: 2.16 × 108 | AV: 5.00 × 107 | AV: 5.18 × 106 |
| SD: 1.79 × 107 | SD: 7.54 × 107 | SD: 1.22 × 107 | SD: 5.45 × 105 | |
| 6 | AV: 2.62 × 108 | AV: 2.92 × 108 | AV: 1.96 × 108 | AV: 1.90 × 106 |
| SD: 6.18 × 107 | SD: 7.29 × 107 | SD: 1.01 × 108 | SD: 1.13 × 106 | |
| 7 | AV: 9.20 × 107 | AV: 1.34 × 108 | AV: 8.80 × 107 | AV: 1.46 × 106 |
| SD: 9.88 × 106 | SD: 3.91 × 107 | SD: 4.55 × 107 | SD: 7.47 × 105 | |
| 8 | AV: 2.84 × 108 | AV: 1.50 × 108 | AV: 8.20 × 107 | AV: 3.78 × 106 |
| SD: 6.84 × 107 | SD: 8.60 × 107 | SD: 2.49 × 107 | SD: 1.03 × 106 | |
| 9 | AV: 2.40 × 108 | AV: 1.56 × 108 | AV: 1.03 × 108 | AV: 6.78 × 106 |
| SD: 3.14 × 107 | SD: 1.32 × 107 | SD: 6.02 × 107 | SD: 9.83 × 105 | |
| 10 | AV: 1.56 × 108 | AV: 2.20 × 107 | AV: 2.72 × 107 | AV: 1.94 × 106 |
| SD: 9.32 × 107 | SD: 1.10 × 107 | SD: 1.49 × 107 | SD: 8.35 × 105 |
| Flavonoid Compound | Control | 9 B | 3 F |
|---|---|---|---|
| Daidzein | 2019.5 ± 5.1 | 3258.9 ± 7.8 | 3740.7 ± 3.1 |
| Daidzin | 12,868.3 ± 23.5 | 52.3 ± 0.1 | 28.2 ± 0.1 |
| Genistein | 2596.1 ± 2.2 | 4020.9 ± 3.2 | 4699.6 ± 2.4 |
| Genistin | 15,200.0 ± 56.8 | 534.5 ± 0.7 | 86.9 ± 0.1 |
| Glycitein | 1108.3 ± 1.1 | 901.4 ± 1.2 | 1391.5 ± 0.8 |
| Glycitin | 5377.561 ± 3.1 | <LOD | <LOD |
| Sum | 39,169.8 | 8768.0 | 9946.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dygas, D.; Liszkowska, W.; Steglińska, A.; Sulyok, M.; Kręgiel, D.; Berłowska, J. Rapeseed Meal Waste Biomass as a Single-Cell Protein Substrate for Nutritionally-Enhanced Feed Components. Processes 2023, 11, 1556. https://doi.org/10.3390/pr11051556
Dygas D, Liszkowska W, Steglińska A, Sulyok M, Kręgiel D, Berłowska J. Rapeseed Meal Waste Biomass as a Single-Cell Protein Substrate for Nutritionally-Enhanced Feed Components. Processes. 2023; 11(5):1556. https://doi.org/10.3390/pr11051556
Chicago/Turabian StyleDygas, Dawid, Wiktoria Liszkowska, Aleksandra Steglińska, Michael Sulyok, Dorota Kręgiel, and Joanna Berłowska. 2023. "Rapeseed Meal Waste Biomass as a Single-Cell Protein Substrate for Nutritionally-Enhanced Feed Components" Processes 11, no. 5: 1556. https://doi.org/10.3390/pr11051556
APA StyleDygas, D., Liszkowska, W., Steglińska, A., Sulyok, M., Kręgiel, D., & Berłowska, J. (2023). Rapeseed Meal Waste Biomass as a Single-Cell Protein Substrate for Nutritionally-Enhanced Feed Components. Processes, 11(5), 1556. https://doi.org/10.3390/pr11051556