
Characterization of Sargassum spp. from the Mexican Caribbean and Its Valorization through Fermentation Process


 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Sargassum Identification
3.2. Sargassum Characterization
3.3. Microbial Growth Rate
3.4. Total Polyphenol Concentrations
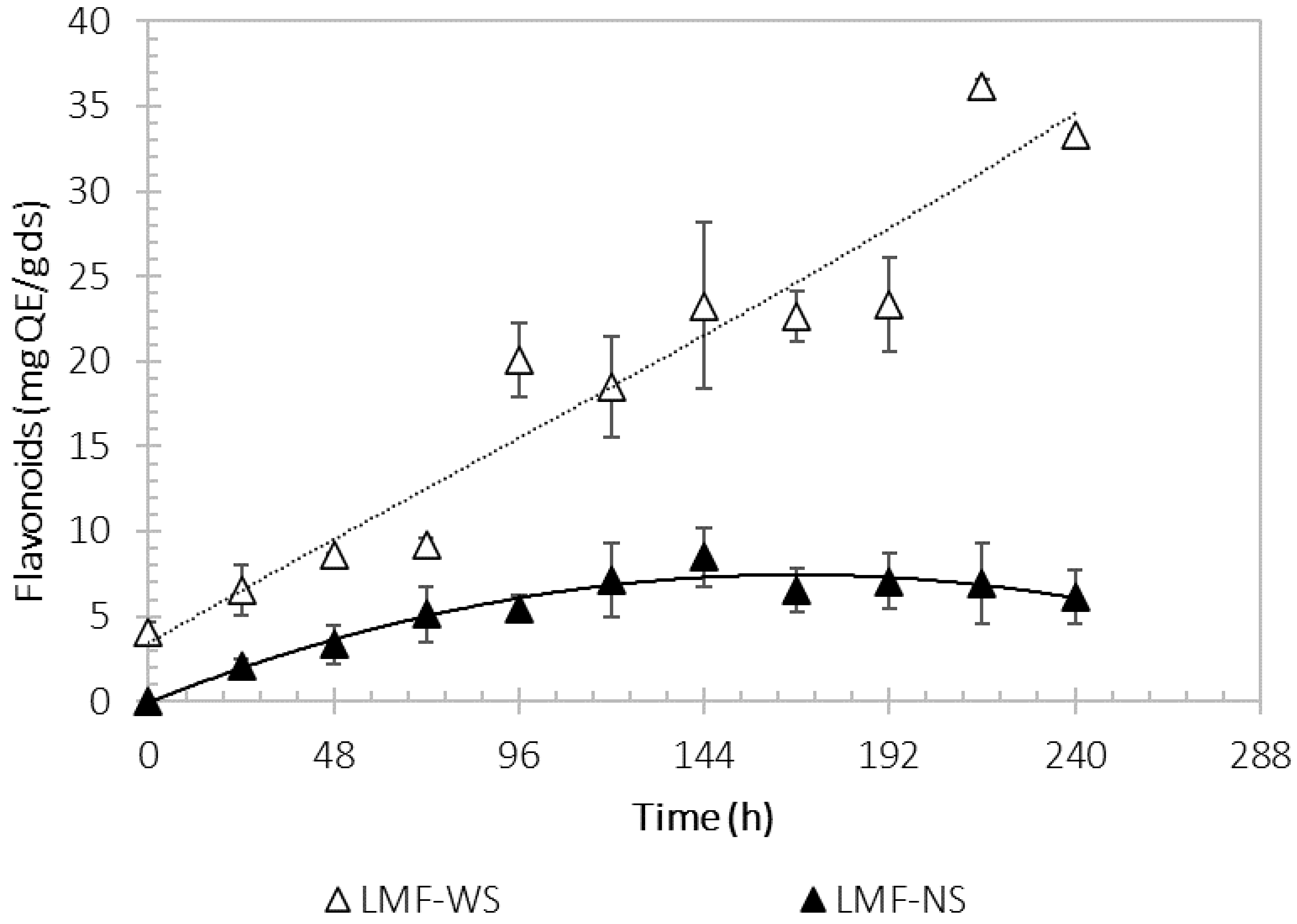
3.5. Flavonoid Concentration
3.6. Polysaccharides
3.7. ABTS Antioxidant Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chávez, V.; Uribe-Martínez, A.; Cuevas, E.; Rodríguez-Martínez, R.E.; van Tussenbroek, B.I.; Francisco, V.; Estévez, M.; Celis, L.B.; Monroy-Velázquez, L.V.; Leal-Bautista, R. Massive influx of pelagic Sargassum spp. on the coasts of the Mexican Caribbean 2014–2020: Challenges and opportunities. Water 2020, 12, 2908. [Google Scholar] [CrossRef]
- Leal-Bautista, R.M.; Tapia-Tussel, R.; Alzate-Gaviria, L. Usos potenciales del sargazo. Rev. Acad. Méxicana Cienc. 2020, 71, 52–57. [Google Scholar]
- Martínez-González, G. Sargassum: The atypical irruption of an ancient ecosystem. Salud Pública México 2019, 61, 698–700. [Google Scholar] [CrossRef] [PubMed]
- Sentíes, A.; Dreckmann, K.M. Lista actualizada de las macroalgas de Tabasco, México. Acta Bot. Venez. 2013, 36, 109–118. [Google Scholar]
- Marx, U.C.; Roles, J.; Hankamer, B. Sargassum blooms in the Atlantic Ocean–From a burden to an asset. Algal Res. 2021, 54, 102–188. [Google Scholar] [CrossRef]
- Devault, D.A.; Pierre, R.; Marfaing, H.; Dolique, F.; Lopez, P.-J. Sargassum contamination and consequences for downstream uses: A review. J. Appl. Phycol. 2021, 33, 567–602. [Google Scholar] [CrossRef]
- Hardouin, K.; Bedoux, G.; Burlot, A.-S.; Nyvall-Collén, P.; Bourgougnon, N. Enzymatic recovery of metabolites from seaweeds: Potential applications. Adv. Bot. Res. 2014, 7, 1279–1320. [Google Scholar]
- Shobharani, P.; Nanishankar, V.; Halami, P.; Sachindra, N. Antioxidant and anticoagulant activity of polyphenol and polysaccharides from fermented Sargassum sp. Int. J. Biol. Macromol. 2014, 65, 542–548. [Google Scholar] [CrossRef]
- Aparicio, E.; Rodríguez-Jasso, R.M.; Pinales-Márquez, C.D.; Loredo-Treviño, A.; Robledo-Olivo, A.; Aguilar, C.N.; Kostas, E.T.; Ruiz, H.A. High-pressure technology for Sargassum spp. biomass pretreatment and fractionation in the third generation of bioethanol production. Bioresour. Technol. 2021, 329, 124935. [Google Scholar] [CrossRef]
- Ardalan, Y.; Jazini, M.; Karimi, K. Sargassum angustifolium brown macroalga as a high potential substrate for alginate and ethanol production with minimal nutrient requirement. Algal Res. 2018, 36, 29–36. [Google Scholar] [CrossRef]
- Thompson, T.; Young, B.; Baroutian, S. Pelagic Sargassum for energy and fertiliser production in the Caribbean: A case study on Barbados. Renew. Sustain. Energy Rev. 2020, 118, 109564. [Google Scholar] [CrossRef]
- Davis, D.; Simister, R.; Campbell, S.; Marston, M.; Bose, S.; McQueen-Mason, S.J.; Gomez, L.D.; Gallimore, W.A.; Tonon, T. Biomass composition of the golden tide pelagic seaweeds Sargassum fluitans and S. natans (morphotypes I and VIII) to inform valorisation pathways. Sci. Total Environ. 2021, 762, 143134. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Lin, X.; Wang, J.; Liu, Y.; Tao, H.; Zhou, X. Asperpyrone-Type Bis-Naphtho-γ-Pyrones with COX-2–Inhibitory Activities from Marine-Derived Fungus Aspergillus niger. Molecules 2016, 21, 941. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.A.S.; Oliveira, M.d.C.F.d.; Pimenta, A.T.; Uchôa, P.K. Aspergillus niger: A hundred years of contribution to the natural products chemistry. J. Braz. Chem. Soc. 2019, 30, 2029–2059. [Google Scholar] [CrossRef]
- Charles-Rodríguez, V.; Guerrero-Mata, A.; Martínez-Vázquez, G.; Cruz-Hernández, M.A.; Belmares-Cerda, R.E.; Robledo, A. Bioreactor Analysis for the Corn-Cob Valorization in the Xylanase Production. Waste Biomass Valorization 2018, 9, 995–1001. [Google Scholar] [CrossRef]
- Orzua, M.C.; Mussatto, S.I.; Contreras-Esquivel, J.C.; Rodriguez, R.; de la Garza, H.; Teixeira, J.A.; Aguilar, C.N. Exploitation of agro industrial wastes as immobilization carrier for solid-state fermentation. Ind. Crops Prod. 2009, 30, 24–27. [Google Scholar] [CrossRef]
- Govindarajan, A.F.; Cooney, L.; Whittaker, K.; Bloch, D.; Burdorf, R.M.; Canning, S.; Carter, C.; Cellan, S.M.; Eriksson, F.A.A.; Freyer, H.; et al. The distribution and mitochondrial genotype of the hydroid Aglaophenia latecarinata is correlated with its pelagic Sargassum substrate type in the tropical and subtropical western Atlantic Ocean. PeerJ 2019, 7, e7814. [Google Scholar] [CrossRef]
- Godínez-Ortega, J.L.; Cuatlán-Cortés, J.V.; López-Bautista, J.M.; van Tussenbroek, B.I. A Natural History of Floating Sargassum Species (Sargasso) from Mexico; IntechOpen: London, UK, 2021. [Google Scholar]
- Mendoza-Becerril, M.A.; Serviere-Zaragoza, E.; Mazariegos-Villarreal, A.; Rivera-Perez, C.; Calder, D.R.; Vázquez-Delfín, E.F.; Freile-Pelegrín, Y.; Agüero, J.; Robledo, D. Epibiont hydroids on beachcast Sargassum in the Mexican Caribbean. PeerJ 2020, 8, e9795. [Google Scholar] [CrossRef]
- Schell, J.; Goodwin, D.; Siuda, A. Recent Sargassum Inundation Events in the Caribbean: Shipboard Observations Reveal Dominance of a Previously Rare Form. Oceanography 2015, 28, 8–10. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2016. [Google Scholar]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Waterman, P.G.; Mole, S. Analysis of Phenolic Plant Metabolites; Blackwell Scientific: London, UK, 1994. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colometric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Rodríguez-Luna, D.; Ruiz, H.A.; González-Morales, S.; Sandoval-Rangel, A.; Cabrera de la Fuente, M.; Charles-Rodríguez, A.V.; Robledo-Olivo, A. Recovery of melon residues (Cucumis melo) to produce lignocellulolytic enzymes. Biomass Convers. Biorefinery 2020, 12, 5915–5922. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Camacho, O.; Mattio, L.; Draisma, S.; Fredericq, S.; Diaz-Pulido, G. Morphological and molecular assessment of Sargassum (Fucales, Phaeophyceae) from Caribbean Colombia, including the proposal of Sargassum giganteum sp. nov., Sargassum schnetteri comb. nov. and Sargassum section Cladophyllum sect. nov. Syst. Biodivers. 2015, 13, 105–130. [Google Scholar] [CrossRef]
- Vázquez-Delfín, E.; Freile-Pelegrín, Y.; Salazar-Garibay, A.; Serviere-Zaragoza, E.; Méndez-Rodríguez, L.C.; Robledo, D. Species composition and chemical characterization of Sargassum influx at six different locations along the Mexican Caribbean coast. Sci. Total Environ. 2021, 795, 148852. [Google Scholar] [CrossRef] [PubMed]
- García-Sánchez, M.; Graham, C.; Vera, E.; Escalante-Mancera, E.; Álvarez-Filip, L.; van Tussenbroek, B.I. Temporal changes in the composition and biomass of beached pelagic Sargassum species in the Mexican Caribbean. Aquat. Bot. 2020, 167, 103275. [Google Scholar] [CrossRef]
- Robledo, D.; Vázquez-Delfín, E.; Freile-Pelegrín, Y.; Vásquez-Elizondo, R.M.; Qui-Minet, Z.N.; Salazar-Garibay, A. Challenges and Opportunities in Relation to Sargassum Events Along the Caribbean Sea. Front. Mar. Sci. 2021, 8, 699664. [Google Scholar] [CrossRef]
- Saldarriaga-Hernandez, S.; Hernandez-Vargas, G.; Iqbal, H.M.N.; Barceló, D.; Parra-Saldívar, R. Bioremediation potential of Sargassum sp. biomass to tackle pollution in coastal ecosystems: Circular economy approach. Sci. Total Environ. 2020, 715, 136978. [Google Scholar] [CrossRef]
- Minatel, I.O.; Borges, C.V.; Ferreira, M.I.; Gomez, H.A.G.; Chen, C.-Y.O.; Lima, G.P.P. Phenolic Compounds: Functional Properties, Impact of Processing and Bioavailability; IntechOpen: London, UK, 2017. [Google Scholar]
- Ouattara, A.; Kpan, W.B.; Komoe, K.; Ouattara, D.; Kone, M.W. The acute toxicity of Sargassum fuiltans (Børgesen) Børgesen and Sargassum natans (Børgesen) Børgesen on some rats of wistar stock. J. Anim. Plant Sci. 2021, 47, 9. [Google Scholar]
- Oyesiku, O.O.; Egunyomi, A. Identification and chemical studies of pelagic masses of Sargassum natans (Linnaeus) Gaillon and S. fluitans (Borgessen) Borgesen (brown algae), found offshore in Ondo State, Nigeria. Afr. J. Biotechnol. 2014, 13, 1188–1193. [Google Scholar]
- Reyes-Ocampo, I.; González-Brambila, M.; López-Isunza, F. Un análisis del metabolismo de Aspergillus niger creciendo sobre un sustrato sólido. Rev. Mex. Ing. Química 2013, 12, 41–56. [Google Scholar]
- Bizukojc, M.; Gonciarz, J. Influence of oxygen on lovastatin biosynthesis by Aspergillus terreus ATCC 20542 quantitatively studied on the level of individual pellets. Bioprocess Biosyst. Eng. 2015, 38, 1251–1266. [Google Scholar] [CrossRef]
- Zapata Bustamante, S.; Tamayo Tenorio, A.; Alberto Rojano, B. Efecto de la fermentación sobre la actividad antioxidante de diferentes clones de cacao Colombia. Rev. Cuba. Plantas Med. 2013, 18, 391–404. [Google Scholar]
- Rubio, M.C.; Alurralde, T.; Suárez, S.; Navarro, A. Producción de naringenina por Aspergillus niger IB-56. Boletín Micológico 2011, 26, 23–27. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Telles, C.B.S.; Dantas-Santos, N.; Camara, R.B.G.; Cordeiro, S.L.; Pereira Costa, M.S.S.; Almeida-Lima, J.; Melo-Silveira, R.F.; Oliveira, R.M. Antioxidant and antiproliferative activities of heterofucans from the seaweed Sargassum filipendula. Mar. Drugs 2011, 9, 952–966. [Google Scholar] [CrossRef] [PubMed]
- Hifney, A.F.; Fawzy, M.A.; Abdel-Gawad, K.M.; Gomaa, M. Upgrading the antioxidant properties of fucoidan and alginate from Cystoseira trinodis by fungal fermentation or enzymatic pretreatment of the seaweed biomass. Food Chem. 2018, 269, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Cuellar Alvarez, L.; Cuellar Alvarez, N.; Galeano Garcia, P.; Suárez Salazar, J.C. Effect of fermentation time on phenolic content and antioxidant potential in Cupuassu (Theobroma grandiflorum (Willd. ex Spreng.) K. Schum.) beans. Acta Agronómica 2017, 66, 473–479. [Google Scholar] [CrossRef]
- Zhao, Y.-S.; Eweys, A.S.; Zhang, J.-Y.; Zhu, Y.; Bai, J.; Darwesh, O.M.; Zhang, H.-B.; Xiao, X. Fermentation Affects the Antioxidant Activity of Plant-Based Food Material through the Release and Production of Bioactive Components. Antioxidants 2021, 10, 2004. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COMPONENTS | Washed Sargassum | Natural Sargassum |
|---|---|---|
| Humidity (%) | 12.00 ± 0.56 | 12.00 ± 0.70 |
| Ash (%) | 18.40 ± 1.87 | 19.30 ± 1.11 |
| Crude fiber (%) | 20.00 ± 2.12 | 22.00 ± 3.87 |
| Fat (%) | 5.36 ± 0.35 | 6.02 ± 0.31 |
| Proteins (%) | 5.47 ± 0.98 | 8.30 ± 0.14 |
| Carbohydrates (%) (by difference) | 38.77 ± 3.58 | 32.38 ± 1.76 |
| Total phenolic compounds (mg GAE/g ds) | 4.35 ± 0.83 | 4.25 ± 0.94 |
| Flavonoids (mg QE/g ds) | 5.77 ± 0.49 | 3.92 ± 0.91 |
| Fucoidan (%) | 7.18 ± 0.29 | 7.09 ± 0.95 |
| Glucan (%) | 10.74 ± 0.28 | 10.41 ± 0.79 |
| Galactan (%) | 5.16 ± 0.29 | 5.59 ± 0.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paredes-Camacho, R.M.; González-Morales, S.; González-Fuentes, J.A.; Rodríguez-Jasso, R.M.; Benavides-Mendoza, A.; Charles-Rodríguez, A.V.; Robledo-Olivo, A. Characterization of Sargassum spp. from the Mexican Caribbean and Its Valorization through Fermentation Process. Processes 2023, 11, 685. https://doi.org/10.3390/pr11030685
Paredes-Camacho RM, González-Morales S, González-Fuentes JA, Rodríguez-Jasso RM, Benavides-Mendoza A, Charles-Rodríguez AV, Robledo-Olivo A. Characterization of Sargassum spp. from the Mexican Caribbean and Its Valorization through Fermentation Process. Processes. 2023; 11(3):685. https://doi.org/10.3390/pr11030685
Chicago/Turabian StyleParedes-Camacho, Rosa María, Susana González-Morales, José Antonio González-Fuentes, Rosa María Rodríguez-Jasso, Adalberto Benavides-Mendoza, Ana Verónica Charles-Rodríguez, and Armando Robledo-Olivo. 2023. "Characterization of Sargassum spp. from the Mexican Caribbean and Its Valorization through Fermentation Process" Processes 11, no. 3: 685. https://doi.org/10.3390/pr11030685
APA StyleParedes-Camacho, R. M., González-Morales, S., González-Fuentes, J. A., Rodríguez-Jasso, R. M., Benavides-Mendoza, A., Charles-Rodríguez, A. V., & Robledo-Olivo, A. (2023). Characterization of Sargassum spp. from the Mexican Caribbean and Its Valorization through Fermentation Process. Processes, 11(3), 685. https://doi.org/10.3390/pr11030685