
Cyclic Production of Galacto-Oligosaccharides through Ultrafiltration-Assisted Enzyme Recovery



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Enzyme Activity Assay
2.3. Enzymatic Conversion in Stirred Tank Reactor (STR)
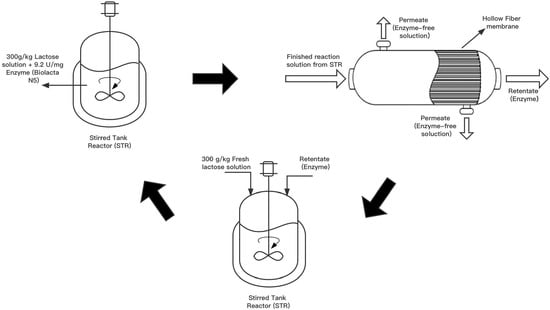
2.4. Cyclic Production of GOS in an Ultrafiltration-Assisted Reactor (Cyclic-EMR)
- In the first step, a traditional STR was employed to carry out a batchwise reaction. A 9.5 kg reaction solution with an initial lactose concentration of 300 g·kg−1 was introduced in the reactor TK-1. The reaction was performed at 50 °C and pH 6.0, using an initial enzyme activity of 8307 U·kg−1. Samples were regularly taken from the reactor and heat-treated at 90 °C for 30 min prior to the saccharides analysis by HPLC.
- In the second step, the membrane unit M-1 was attached to the reactor, and the reaction liquid was filtered through UF in an inside-out flow configuration at 0.5 bar transmembrane pressure until 8.4 kg of permeate was collected.
- In the third step, the membrane module M-1 was de-attached from the plant. A total of 8.4 kg of fresh substrate solution consisting of 300 g·kg−1 of lactose was added into the concentrated enzyme solution in the reactor. Then, step 1 of the next cycle was started. The de-attached membrane was subject to a cleaning procedure, as detailed in Section 2.5.
2.5. Membrane Regeneration Procedure
2.6. Analysis of Progress Curves
2.7. High Performance Liquid Chromatography (HPLC) Analysis
3. Results
3.1. STR Performance
3.2. Cyclic-EMR Performance
3.3. Quantification of Enzyme Losses
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Symbols
Abbreviations
References
- Galacto-oligosaccharide Market Segmentation By End User (Food & Beverage Industry, Pharmaceutical, Personal Care and Animal Feed)–Global Demand Analysis & Opportunity Outlook 2028. Available online: https://www.researchnester.com/reports/galacto-oligosaccharide-market/2814 (accessed on 1 February 2022).
- Illanes, A.; Vera, C. Enzymatic production of galacto-oligosaccharides. In Lactose-Derived Prebiotics: A Process Perspective; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 111–189. [Google Scholar]
- Pázmándi, M.; Maráz, A.; Ladányi, M.; Kovács, Z. The impact of membrane pretreatment on the enzymatic production of whey-derived galacto-oligosaccharides. J. Food Process. Eng. 2018, 41, e12649. [Google Scholar] [CrossRef]
- Pires, A.F.; Marnotes, N.G.; Rubio, O.D.; Garcia, A.C.; Pereira, C.D. Dairy by-products: A review on the valorization of whey and second cheese whey. Foods 2021, 10, 1067. [Google Scholar] [CrossRef]
- Osman, A.; Tzortzis, G.; Rastall, R.A.; Charalampopoulos, D. A comprehensive investigation of the synthesis of prebiotic galactooligosaccharides by whole cells of Bifidobacterium bifidum NCIMB 41171. J. Biotechnol. 2010, 150, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; O’sullivan, D. Production of galactooligosaccharides using a hyperthermophilic β-galactosidase in permeabilized whole cells of Lactococcus lactis. J. Dairy Sci. 2014, 97, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Pázmándi, M.; Galambos, I.; Kovács, Z. Continuous production of galacto-oligosaccharides by an enzyme membrane reactor utilizing free enzymes. Membranes 2020, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Sen, D.; Sarkar, A.; Bhattacharyya, S.; Bhattacharjee, C. A comparative study on the production of galacto-oligosaccharide from whey permeate in recycle membrane reactor and in enzymatic batch reactor. Ind. Eng. Chem. Res. 2011, 50, 806–816. [Google Scholar] [CrossRef]
- Hackenhaar, C.R.; Spolidoro, L.S.; Flores, E.E.E.; Klein, M.P.; Hertz, P.F. Batch synthesis of galactooligosaccharides from co-products of milk processing using immobilized β-galactosidase from Bacillus circulans. Biocatal. Agric. Biotechnol. 2021, 36, 102136. [Google Scholar] [CrossRef]
- Huerta, L.M.; Vera, C.; Guerrero, C.; Wilson, L.; Illanes, A. Synthesis of galacto-oligosaccharides at very high lactose concentrations with immobilized β-galactosidases from Aspergillus oryzae. Process. Biochem. 2011, 46, 245–252. [Google Scholar] [CrossRef]
- Pázmándi, M.; Kovács, Z.; Balga, E.; Kovács, M.; Maráz, A. Production of high-purity galacto-oligosaccharides by depleting glucose and lactose from galacto-oligosaccharide syrup with yeasts. Yeast 2020, 37, 515–530. [Google Scholar] [CrossRef]
- Gänzle, M.G. Enzymatic synthesis of galacto-oligosaccharides and other lactose derivatives (hetero-oligosaccharides) from lactose. Int. Dairy J. 2012, 22, 116–122. [Google Scholar] [CrossRef]
- Fischer, C.; Kleinschmidt, T. Synthesis of galactooligosaccharides in milk and whey: A review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 678–697. [Google Scholar] [CrossRef] [PubMed]
- Füreder, V.; Rodriguez-Colinas, B.; Cervantes, F.V.; Fernandez-Arrojo, L.; Poveda, A.; Jimenez-Barbero, J.; Ballesteros, A.O.; Plou, F.J. Selective synthesis of galactooligosaccharides containing β (1 → 3) linkages with β-galactosidase from Bifidobacterium bifidum (Saphera). J. Agric. Food Chem. 2020, 68, 4930–4938. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhu, J.; Liu, L.; Yaqoob, M.U.; Pei, X.; Tao, W.; Xiao, Z.; Sun, W.; Wang, M. Optimization for galactooligosaccharides synthesis: A potential alternative for gut health and immunity. Life Sci. 2020, 245, 117353. [Google Scholar] [CrossRef] [PubMed]
- Scott, F.; Vera, C.; Conejeros, R. Technical and economic analysis of industrial production of lactose-derived prebiotics with focus on galacto-oligosaccharides. In Lactose-Derived Prebiotics: A Process Perspective; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 261–284. [Google Scholar]
- Albayrak, N.; Yang, S.T. Production of galacto-oligosaccharides from lactose by Aspergillus oryzae β-galactosidase immobilized on cotton cloth. Biotechnol. Bioeng. 2002, 77, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Torres, P.; Batista-Viera, F. Improved biocatalysts based on Bacillus circulans β-galactosidase immobilized onto epoxy-activated acrylic supports: Applications in whey processing. J. Mol. Catal. B: Enzym. 2012, 83, 57–64. [Google Scholar] [CrossRef]
- Warmerdam, A.; Zisopoulos, F.K.; Boom, R.M.; Janssen, A.E. Kinetic characterization of galacto-oligosaccharide (GOS) synthesis by three commercially important β-galactosidases. Biotechnol. Prog. 2014, 30, 38–47. [Google Scholar] [CrossRef]
- Fujimoto, H.; Miyasato, M.; Ito, Y.; Sasaki, T.; Ajisaka, K. Purification and properties of recombinant β-galactosidase from Bacillus circulans. Glycoconj. J. 1998, 15, 155–160. [Google Scholar] [CrossRef]
- Huang, J.; Zhu, S.; Zhao, L.; Chen, L.; Du, M.; Zhang, C.; Yang, S.-T. A novel β-galactosidase from Klebsiella oxytoca ZJUH1705 for efficient production of galacto-oligosaccharides from lactose. Appl. Microbiol. Biotechnol. 2020, 104, 6161–6172. [Google Scholar] [CrossRef]
- Urrutia, P.; Mateo, C.; Guisán, J.M.; Wilson, L.; Illanes, A. Immobilization of Bacillus circulans β-galactosidase and its application in the synthesis of galacto-oligosaccharides under repeated-batch operation. Biochem. Eng. J. 2013, 77, 41–48. [Google Scholar] [CrossRef]
- An, S.M.; Wu, J.H.; Qian, L.F.; Gao, Y.L.; Wu, Y.; Yu, G.P. Applications of Ultrafiltration-nanofiltration Membrane Continuous Combination Technology for Refining of Milk-Derived Oligosaccharides. In Advanced Materials Research; Trans Tech Publications Ltd.: Bäch, Switzerland, 2013; pp. 1429–1434. [Google Scholar]
- Matella, N.; Dolan, K.; Lee, Y. Comparison of galactooligosaccharide production in free-enzyme ultrafiltration and in immobilized-enzyme systems. J. Food Sci. 2006, 71, C363–C368. [Google Scholar] [CrossRef]
- Warmerdam, A.; Boom, R.M.; Janssen, A.E. β-galactosidase stability at high substrate concentrations. Springerplus 2013, 2, 1–8. [Google Scholar] [CrossRef]
- Su, Z.; Luo, J.; Li, X.; Pinelo, M. Enzyme membrane reactors for production of oligosaccharides: A review on the interdependence between enzyme reaction and membrane separation. Sep. Purif. Technol. 2020, 243, 116840. [Google Scholar] [CrossRef]
- Córdova, A.; Astudillo, C.; Vera, C.; Guerrero, C.; Illanes, A. Performance of an ultrafiltration membrane bioreactor (UF-MBR) as a processing strategy for the synthesis of galacto-oligosaccharides at high substrate concentrations. J. Biotechnol. 2016, 223, 26–35. [Google Scholar] [CrossRef]
- Splechtna, B.; Nguyen, T.-H.; Haltrich, D. Comparison between discontinuous and continuous lactose conversion processes for the production of prebiotic galacto-oligosaccharides using β-galactosidase from Lactobacillus reuteri. J. Agric. Food Chem. 2007, 55, 6772–6777. [Google Scholar] [CrossRef] [PubMed]
- IBM SPSS Statistics for Windows; Version 27.0.; IBM Corp.: Armonk, NY, USA, 2020.
- Rodriguez-Colinas, B.; Poveda, A.; Jimenez-Barbero, J.; Ballesteros, A.O.; Plou, F.J. Galacto-oligosaccharide synthesis from lactose solution or skim milk using the β-galactosidase from Bacillus circulans. J. Agric. Food Chem. 2012, 60, 6391–6398. [Google Scholar] [CrossRef]
- Song, J.; Abe, K.; Imanaka, H.; Imamura, K.; Minoda, M.; Yamaguchi, S.; Nakanishi, K. Causes of the production of multiple forms of β-galactosidase by Bacillus circulans. Biosci. Biotechnol. Biochem. 2011, 75, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Vetere, A.; Paoletti, S. Separation and characterization of three β-galactosidases from Bacillus circulans. Biochim. Et Biophys. Acta Gen. Subj. 1998, 1380, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Boon, M.; Janssen, A.; Van der Padt, A. Modelling and parameter estimation of the enzymatic synthesis of oligosaccharides by β-galactosidase from Bacillus circulans. Biotechnol. Bioeng. 1999, 64, 558–567. [Google Scholar] [CrossRef]
- Palai, T.; Mitra, S.; Bhattacharya, P.K. Kinetics and design relation for enzymatic conversion of lactose into galacto-oligosaccharides using commercial grade β-galactosidase. J. Biosci. Bioeng. 2012, 114, 418–423. [Google Scholar] [CrossRef]
- Botelho, V.A.; Mateus, M.; Petrus, J.C.; de Pinho, M.N. Membrane Bioreactor for Simultaneous Synthesis and Fractionation of Oligosaccharides. Membranes 2022, 12, 171. [Google Scholar] [CrossRef]
- Córdova, A.; Astudillo, C.; Giorno, L.; Guerrero, C.; Conidi, C.; Illanes, A.; Cassano, A. Nanofiltration potential for the purification of highly concentrated enzymatically produced oligosaccharides. Food Bioprod. Process. 2016, 98, 50–61. [Google Scholar] [CrossRef]
- Córdova, A.; Astudillo, C.; Guerrero, C.; Vera, C.; Illanes, A. Assessment of the fouling mechanisms of an ultrafiltration membrane bioreactor during synthesis of galacto-oligosaccharides: Effect of the operational variables. Desalination 2016, 393, 79–89. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Type/ Microorganism | Reactor Configuration | Reaction Conditions | Half-Life (h) | Deactivation Model | Activity Measurement Method | Ref. | ||
|---|---|---|---|---|---|---|---|---|
| Lactose Conc. | pH | Temp. (°C) | ||||||
| β-galactosidase/Aspergillus oryzae | Batch (FE) | 0 g·L−1 | 4.5 | 40 | 399 | Equation (6) | DM * | [17] |
| 50 | 49 | |||||||
| 60 | 2 | |||||||
| Continuous (IE) | 200 g·L−1 | 4.5 | 40 | 10,040 | ||||
| 50 | 1155 | |||||||
| 60 | 49 | |||||||
| β-galactosidase/Aspergillus oryzae | Batch (FE) | 0.1 g·L−1 | 4.5 | 50 | 42 | Equation (2) | oNPG | [10] |
| 55 | 7.4 | |||||||
| 60 | 0.8 | |||||||
| Batch (IE) | 2% w·v−1 | 4.5 | 50 | 163–166 | ||||
| 55 | 9.3–20.5 | |||||||
| 60 | 0.9–1.9 | |||||||
| Biolacta N5/Bacillus circulans | Batch (FE) | 30% w·w−1 | 6 | 25 | 29 | Equation (6) | oNPG | [25] |
| 40 | 29 | |||||||
| 60 | 16 | |||||||
| Biolacta N5/Bacillus circulans | Batch (FE) | 4.6% w·w−1 | 6 | 50 | 12 | Equation (5) | oNPG | [18] |
| Batch (IE) | 4.6% w·w−1 | 6 | 50 | 21–387 | ||||
| Biolactasa-NTL CONC X2/Bacillus circulans | Batch (FE) | 0 g·L−1 | 6 | 60 | ~0.05 | Equation (3) | oNPG | [22] |
| Batch (IE) | 0 g·L−1 | 6 | 60 | 0.25–3 | ||||
| β-galactosidase/Klebsiella oxytoca ZJUH1705 | Batch (FE) β-gal 1 | 40% w·w−1 | 7 | 30 | 141.67 | Equation (6) | oNPG | [21] |
| 40 | 18.33 | |||||||
| 50 | 0.04 | |||||||
| 60 | 0.01 | |||||||
| Batch (FE) β-gal 2 | 40% w·w−1 | 7 | 30 | 88.33 | ||||
| 40 | 1.17 | |||||||
| 50 | 0.17 | |||||||
| 60 | 0.01 | |||||||
| Enzyme Activity (U·kg−1) | Parameters | Estimate | Std. Error | 95% Confidence Interval | F (2; df2) | p1 × p2 | ||
|---|---|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | |||||||
| 923 | −162.07 * | 3.11 | 155.30 | 168.84 | 28,504.6 * df2 = 12 | >0.99 * | −22.69 | |
| 0.14 * | 0.01 | 0.13 | 0.15 | |||||
| 2307 | −172.91 * | 2.92 | 166.56 | 179.27 | 5352.4 * df2 = 12 | 0.99 * | −70.03 | |
| 0.41 * | 0.03 | 0.35 | 0.46 | |||||
| 4615 | −180.19 * | 3.35 | 172.60 | 187.76 | 4539.8 * df2 = 9 | 0.99 * | −134.24 | |
| 0.75 * | 0.06 | 0.62 | 0.87 | |||||
| 6923 | −177.63 * | 1.64 | 174.07 | 181.20 | 6763.6 * df2 = 12 | 0.99 * | −220.44 | |
| 1.24 * | 0.08 | 1.08 | 1.41 | |||||
| 8307 | −179.05 * | 1.72 | 175.53 | 182.56 | 8630.4 * df2 = 29 | 0.99 * | −237.95 | |
| 1.33 * | 0.07 | 1.20 | 1.46 | |||||
| Saccharides Compounds | No-Intercept Linear Regression Slopes | R2 |
|---|---|---|
| DP2 | 0.03 * | 0.997 * |
| DP3 | 0.025 * | 0.991 * |
| DP4 | 0.004 * | 0.995 * |
| DP3-6 | 0.024 * | 0.997 * |
| Glucose | 0.007 * | 0.999 * |
| Cycles | |||||
|---|---|---|---|---|---|
| Fractions | No.1 | No.2 | No.3 | No.4 | No.5 |
| DP2 | 7077 | 2124 | 846 | 388 | 329 |
| DP3 | 7925 | 2480 | 890 | 379 | 306 |
| DP3-6 | 6999 | 2081 | 785 | 355 | 295 |
| Glucose | 7903 | 2588 | 992 | 448 | 400 |
| Mean | 7476 | 2318 | 878 | 392 | 333 |
| STDEV | 507 | 254 | 87 | 40 | 47 |
| Model | k1 | k2 | α1 | α2 | R2 | SSR |
|---|---|---|---|---|---|---|
| Equation (2) | 9.692 × 10−1 | 4.839 × 10−2 | 8.125 × 10−1 | 2.977 × 10−2 | 0.9974 | 7.338 × 10−3 |
| Equation (3) | 5.947 × 10−2 | 9.980 × 10−3 | 7.493 × 10−2 | 0 | 0.9987 | 3.535 × 10−3 |
| Equation (5) | 5.537 × 10−2 | 0 | 3.651 × 10−2 | 0 | 0.9986 | 3.674 × 10−3 |
| Equation (6) | 4.891 × 10−2 | 0 | 0 | 0 | 0.9957 | 1.194 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, T.; Kovács, Z.; Ladányi, M. Cyclic Production of Galacto-Oligosaccharides through Ultrafiltration-Assisted Enzyme Recovery. Processes 2023, 11, 225. https://doi.org/10.3390/pr11010225
Cao T, Kovács Z, Ladányi M. Cyclic Production of Galacto-Oligosaccharides through Ultrafiltration-Assisted Enzyme Recovery. Processes. 2023; 11(1):225. https://doi.org/10.3390/pr11010225
Chicago/Turabian StyleCao, Teng, Zoltán Kovács, and Márta Ladányi. 2023. "Cyclic Production of Galacto-Oligosaccharides through Ultrafiltration-Assisted Enzyme Recovery" Processes 11, no. 1: 225. https://doi.org/10.3390/pr11010225
APA StyleCao, T., Kovács, Z., & Ladányi, M. (2023). Cyclic Production of Galacto-Oligosaccharides through Ultrafiltration-Assisted Enzyme Recovery. Processes, 11(1), 225. https://doi.org/10.3390/pr11010225