
A Review of In Vitro Methods for Measuring the Glycemic Index of Single Foods: Understanding the Interaction of Mass Transfer and Reaction Engineering by Dimensional Analysis



Abstract

1. Introduction
2. In Vivo Methods for GI Measurement
2.1. ISO 26642:2010 Method
2.2. The Opportunities for Using Alternative Methods
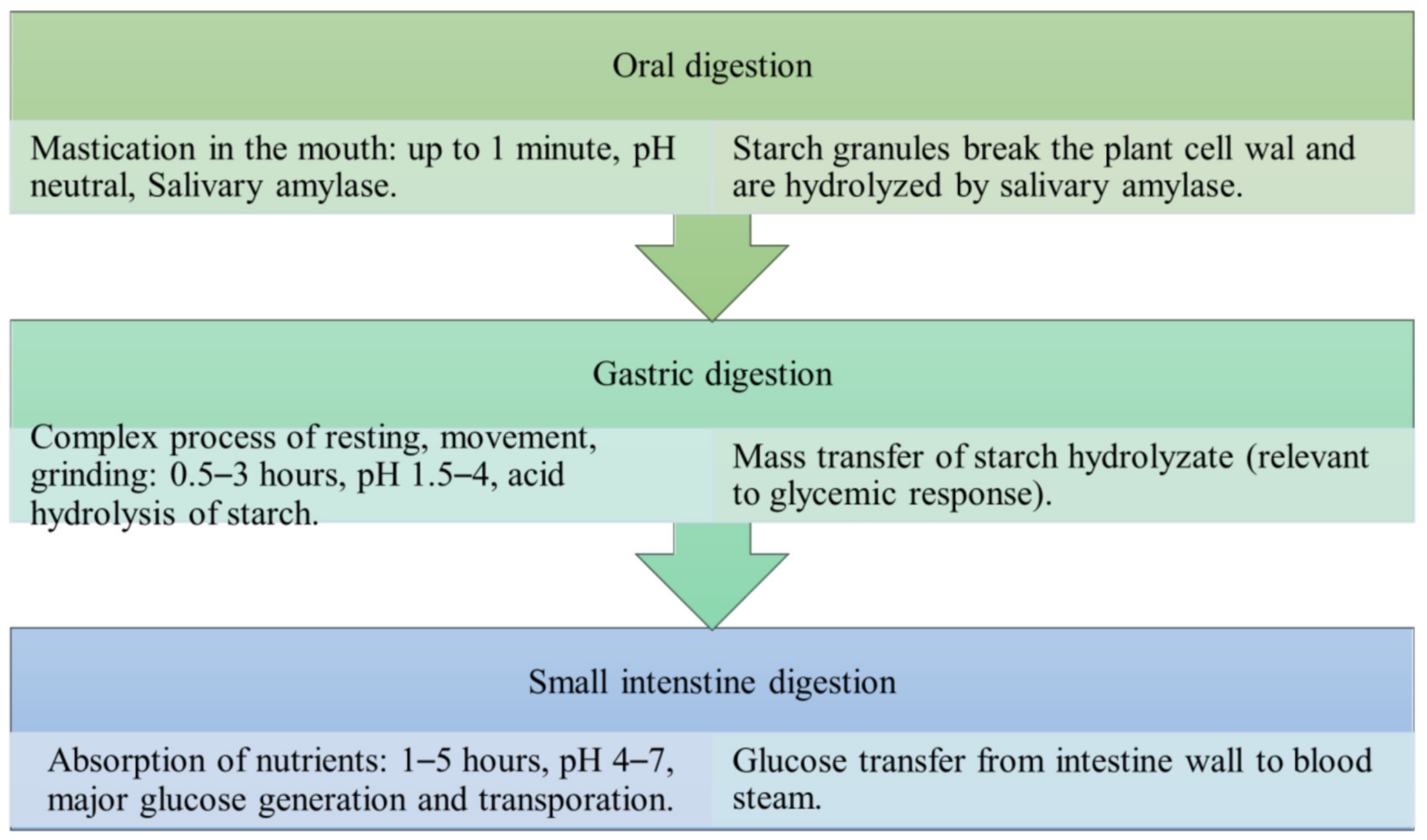
3. Food Digestion and Process
3.1. Carbohydrates
3.2. Digestion Process
3.3. Starch Digestion
4. In Vitro Digestion
4.1. Digestion Models
4.2. In Vitro Starch Digestion
4.3. Engineering Perspectives
4.4. Estimating Damköhler Numbers from Literature Reaction Data
= 2.77 z
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jenkins, D.J.; Wolever, T.M.; Taylor, R.H.; Barker, H.; Fielden, H.; Baldwin, J.M.; Bowling, A.C.; Newman, H.C.; Jenkins, A.L.; Goff, D.V. Glycemic index of foods: A physiological basis for carbohydrate exchange. Am. J. Clin. Nutr. 1981, 34, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, F.S.; Foster-Powell, K.; Brand-Miller, J.C. International tables of glycemic index and glycemic load values: 2008. Diabetes Care 2008, 31, 2281–2283. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, D.S. The glycemic index: Physiological mechanisms relating to obesity, diabetes, and cardiovascular disease. JAMA 2002, 287, 2414–2423. [Google Scholar] [CrossRef] [PubMed]
- Brand-Miller, J.; Foster-Powell, K.; Holt, S. The New Glucose Revolution Complete Guide to Glycemic Index Values; Marlowe & Company: Boston, MA, USA, 2003. [Google Scholar]
- Warren, J.M.; Henry, C.J.K.; Simonite, V. Low glycemic index breakfasts and reduced food intake in preadolescent children. Pediatrics 2003, 112, e414. [Google Scholar] [CrossRef]
- Thomas, D.E.; Brotherhood, J.R.; Brand, J.C. Carbohydrate feeding before exercise: Effect of glycemic index. Int. J. Sports Med. 1991, 12, 180–186. [Google Scholar] [CrossRef]
- Baghurst, K.I. Dietary guidelines: The development process in Australia and New Zealand. J. Am. Diet. Assoc. 2003, 103 (Suppl. S12), 17–21. [Google Scholar] [CrossRef]
- Brand-Miller, J.; McMillan-Price, J.; Steinbeck, K.; Caterson, I. Dietary glycemic index: Health implications. J. Am. Coll. Nutr. 2009, 28 (Suppl. S4), 446s–449s. [Google Scholar] [CrossRef]
- Garsetti, M.; Vinoy, S.; Lang, V.; Holt, S.; Loyer, S.; Brand-Miller, J.C. The glycemic and insulinemic index of plain sweet biscuits: Relationships to in vitro starch digestibility. J. Am. Coll. Nutr. 2005, 24, 441–447. [Google Scholar] [CrossRef]
- Barclay, A.W.; Flood, V.M.; Brand-Miller, J.C.; Mitchell, P. Validity of carbohydrate, glycaemic index and glycaemic load data obtained using a semi-quantitative food-frequency questionnaire. Public Health Nutr. 2008, 11, 573–580. [Google Scholar] [CrossRef]
- Englyst, K.; Vinoy, S.; Englyst, H.; Lang, V. Glycaemic index of cereal products explained by their rapidly and slowly available glucose. Br. J. Nutr. 2003, 89, 329–340. [Google Scholar] [CrossRef]
- Jenkins, D.J.A.; Wolever, T.M.S.; Collier, G.; Ocana, A.; Rao, V.; Buckley, G.; Lam, Y.; Mayer, A.; Thompson, L. Metaboliceffects of low-glycemic-index diet. Am. J. Clin. Nutr. 1988, 46, 968–975. [Google Scholar] [CrossRef] [PubMed]
- Urooj, A.; Puttaraj, S. Glycaemic responses to cereal-based Indian food preparations in patients with non-insulin-dependent diabetes mellitus and normal subjects. Br. J. Nutr. 2000, 83, 483–488. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vinoy, S.; Normand, S.; Meynier, A.; Sothier, M.; Louche-Pelissier, C.; Peyrat, J.; Maitrepierre, C.; Nazare, J.-A.; Brand-Miller, J.; Laville, M. Cereal processing influences postprandial glucose metabolism as well as the GI effect. J. Am. Coll. Nutr. 2013, 32, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Kendall, C.W.; Augustin, L.S.; Franceschi, S.; Hamidi, M.; Marchie, A.; Jenkins, A.L.; Axelsen, M. Glycemic index: Overview of implications in health and disease. Am. J. Clin. Nutr. 2002, 76, 266S–273S. [Google Scholar] [CrossRef]
- Dona, A.C.; Landrey, K.; Atkinson, F.S.; Miller, J.C.B.; Kuchel, P.W. 1H NMR spectroscopy for the in vitro understanding of the glycaemic index. Br. J. Nutr. 2013, 109, 1934–1939. [Google Scholar] [CrossRef]
- Ek, K.L.; Wang, S.; Copeland, L.; Brand-Miller, J.C. Discovery of a low-glycaemic index potato and relationship with starch digestion in vitro. Br. J. Nutr. 2014, 111, 699–705. [Google Scholar] [CrossRef]
- Brand-Miller, J.; Holt, S. Testing the glycaemic index of foods: In vivo, not in vitro. Eur. J. Clin. Nutr. 2004, 58, 700–701. [Google Scholar] [CrossRef]
- Louie, J.C.Y.; Moshtaghian, H.; Boylan, S.; Flood, V.M.; Rangan, A.M.; Barclay, A.W.; Brand-Miller, J.C.; Gill, T.P. A systematic methodology to estimate added sugar content of foods. Eur. J. Clin. Nutr. 2015, 69, 154–161. [Google Scholar] [CrossRef]
- Li, H.; Dhital, S.; Gidley, M.J.; Gilbert, R.G. A more general approach to fitting digestion kinetics of starch in food. Carbohydr. Polym. 2019, 225, 115244. [Google Scholar] [CrossRef]
- Nantel, G. Carbohydrates in human nutrition. Food Nutr. Agric. 1999, FNA/ANA 24, 6–10. [Google Scholar]
- International Organization for Standardization. 26642:2010; Food products—Determination of the Glycaemic Index (GI) and Recommendation for Food Classification. ISO: Geneva, Switzerland, 2010.
- Brouns, F.; Bjorck, I.; Frayn, K.N.; Gibbs, A.L.; Lang, V.; Slama, G.; Wolever, T.M.S. Glycaemic index methodology. Nutr. Res. Rev. 2005, 18, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Wolever, T.M.S.; Meynier, A.; Jenkins, A.L.; Brand-Miller, J.C.; Atkinson, F.S.; Gendre, D.; Leuillet, S.; Cazaubiel, M.; Housez, B.; Vinoy, S. Glycemic index and insulinemic index of foods: An interlaboratory study using the ISO 2010 method. Nutrients 2019, 11, 2218. [Google Scholar] [CrossRef] [PubMed]
- Kunst, A. UV-methods with hexokinase and glucose-6-phosphate dehydrogenase. Methods Enzym. Anal. 1984, VI, 163–172. [Google Scholar]
- Sepulveda, J.L. Chapter 9—Challenges in routine clinical chemistry testing analysis of small molecules. In Accurate Results in the Clinical Laboratory, 2nd ed.; Dasgupta, A., Sepulveda, J.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 101–140. [Google Scholar]
- Woolnough, J.W.; Monro, J.A.; Brennan, C.S.; Bird, A.R. Simulating human carbohydrate digestion in vitro: A review of methods and the need for standardisation. Int. J. Food Sci. Technol. 2008, 43, 2245–2256. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food-an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Goff, H.D.; Repin, N.; Fabek, H.; el Khoury, D.; Gidley, M.J. Dietary fibre for glycaemia control: Towards a mechanistic understanding. Bioact. Carbohydr. Diet. Fibre 2018, 14, 39–53. [Google Scholar] [CrossRef]
- Guerra, A.; Etienne-Mesmin, L.; Livrelli, V.; Denis, S.; Blanquet-Diot, S.; Alric, M. Relevance and challenges in modeling human gastric and small intestinal digestion. Trends Biotechnol. 2012, 30, 591–600. [Google Scholar] [CrossRef]
- Bordoloi, A.; Singh, J.; Kaur, L. In vitro digestibility of starch in cooked potatoes as affected by guar gum: Microstructural and rheological characteristics. Food Chem. 2012, 133, 1206–1213. [Google Scholar] [CrossRef]
- Dartois, A.; Singh, J.; Kaur, L.; Singh, H. Influence of guar gum on the in vitro starch digestibility—rheological and microstructural characteristics. Food Biophys. 2010, 5, 149–160. [Google Scholar] [CrossRef]
- Gidley, M.J. Hydrocolloids in the digestive tract and related health implications. Curr. Opin. Colloid Interface Sci. 2013, 18, 371–378. [Google Scholar] [CrossRef]
- Liu, D.; Dhital, S.; Wu, P.; Chen, X.-D.; Gidley, M.J. In vitro digestion of apple tissue using a dynamic stomach model: Grinding and crushing effects on polyphenol bioaccessibility. J. Agric. Food Chem. 2020, 68, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Langrish, T. A comparison of different physical stomach models and an analysis of shear stresses and strains in these system. Food Res. Int. 2020, 135, 109296. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gidley, M.J.; Dhital, S. High-amylose starches to bridge the “fiber gap”: Development, structure, and nutritional functionality. Compr. Rev. Food Sci. Food Saf. 2019, 18, 362–379. [Google Scholar] [CrossRef]
- Ai, Y.; Jane, J.-L. Chapter 3—Understanding Starch Structure and Functionality. In Starch in Food: Structure, Function and Applications, 2nd ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2018; pp. 151–178. [Google Scholar]
- Pérez, S.; Bertoft, E. The molecular structures of starch components and their contribution to the architecture of starch granules: A comprehensive review. Starch Stärke 2010, 62, 389–420. [Google Scholar] [CrossRef]
- Zhou, Z.; Robards, K.; Helliwell, S.; Blanchard, C. Compositionand functional properties of rice. Int. J. Food Sci. Technol. 2002, 37, 849–868. [Google Scholar] [CrossRef]
- Orman, B.A.; Schumann, R.A. Comparison of near-infrared spectroscopy calibration methods for the prediction of protein, oil, and starch in maize grain. J. Agric. Food Chem. 1991, 39, 883–886. [Google Scholar] [CrossRef]
- Robertson, T.M.; Alzaabi, A.Z.; Robertson, M.D.; Fielding, B.A. Starchy carbohydrates in a healthy diet: The role of the humble potato. Nutrients 2018, 10, 1764. [Google Scholar] [CrossRef]
- Wang, N.; Daun, J.K. Quality of Western Canadian Pulse Crops—2006; Canadian Grain Commission: Winnipeg, MB, Canada, 2006.
- Preiss, J. Chapter 1—Plant starch synthesis. In Starch in Food, 2nd ed.; Sjöö, M., Nilsson, L., Eds.; Woodhead Publishing: Sawston, UK, 2018; pp. 3–95. [Google Scholar]
- Takeda, Y.; Shitaozono, T.; Hizukuri, S. Structures of sub-fractions of corn amylose. Carbohydr. Res. 1990, 199, 207–214. [Google Scholar] [CrossRef]
- Hizukuri, S. Polymodal distribution of the chain lengths of amylopectins, and its significance. Carbohydr. Res. 1986, 147, 342–347. [Google Scholar] [CrossRef]
- Hizukuri, S.; Takeda, Y.; Yasuda, M.; Suzuki, A. Multi-branched nature of amylose and the action of debranching enzymes. Carbohydr. Res. 1981, 94, 205–213. [Google Scholar] [CrossRef]
- Li, H.; Dhital, S.; Slade, A.J.; Yu, W.; Gilbert, R.G.; Gidley, M.J. Alteringstarch branching enzymes in wheat generates high-amylose starch with novel molecular structure and functional properties. Food Hydrocoll. 2019, 92, 51–59. [Google Scholar] [CrossRef]
- Manners, D.J.; Matheson, N.K. The fine structure of amylopectin. Carbohydr. Res. 1981, 90, 99–110. [Google Scholar] [CrossRef]
- Berglund, J.; Mikkelsen, D.; Flanagan, B.M.; Dhital, S.; Gaunitz, S.; Henriksson, G.; Lindström, M.E.; Yakubov, G.E.; Gidley, M.J.; Vilaplana, F. Wood hemicelluloses exert distinct biomechanical contributions to cellulose fibrillar networks. Nat. Commun. 2020, 11, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Lal, U.R.; Mukhtar, H.M.; Singh, P.S.; Shah, G.; Dhawan, R.K. Phytochemical profile of sugarcane and its potential health aspects. Pharmacogn. Rev. 2015, 9, 45–54. [Google Scholar] [CrossRef]
- Holland, C.; Ryden, P.; Edwards, C.H.; Grundy, M.M.L. Plant cell walls: Impact on nutrient bioaccessibility and digestibility. Foods 2020, 9, 201. [Google Scholar] [CrossRef]
- Dhital, S.; Bhattarai, R.R.; Gorham, J.; Gidley, M.J. Intactness of cell wall structure controls the in vitro digestion of starch in legumes. Food Funct. 2016, 7, 1367–1379. [Google Scholar] [CrossRef]
- Dhital, S.; Baier, S.K.; Gidley, M.J.; Stokes, J.R. Microstructural properties of potato chips. Food Struct. 2018, 16, 17–26. [Google Scholar] [CrossRef]
- Liu, D.; Lopez-Sanchez, P.; Gidley, M.J. Cellular barriers in apple tissue regulate polyphenol release under different food processing and in vitro digestion conditions. Food Funct. 2019, 10, 3008–3017. [Google Scholar] [CrossRef]
- Beaumont, B.; Plattsburgh, M.D. Experiments and observations on the gastric juice and the physiology of digestion. Dublin J. Med. Chem. Sci. 1835, 22, 49–69. [Google Scholar] [CrossRef]
- Keppler, S.; O’Meara, S.; Bakalis, S.; Fryer, P.J.; Bornhorst, G.M. Characterization of individual particle movement during in vitro gastric digestion in the Human Gastric Simulator (HGS). J. Food Eng. 2020, 264, 109674. [Google Scholar] [CrossRef]
- Bornhorst, G.; Singh, R. Gastric digestion in vivo and in vitro: How the structural aspects of food influence the digestion process. Annu. Rev. Food Sci. Technol. 2014, 111, 32. [Google Scholar] [CrossRef]
- Gavião, M.B.; Bilt, A. Salivary secretion and chewing: Stimulatory effects from artificial and natural foods. J. Appl. Oral Sci. Rev. FOB 2004, 12, 159–163. [Google Scholar] [CrossRef]
- Bilt, A. Oral physiology, mastication and food perception. In Designing Functional Foods: Measuring and Controlling Food Structure Breakdown and Nutrient Absorption; Woodhead Publishing: Sawston, UK, 2009; pp. 1–35. [Google Scholar]
- Agrawal, K.R.; Lucas, P.W.; Prinz, J.F.; Bruce, I.C. Mechanical properties of foods responsible for resisting food breakdown in the human mouth. Arch. Oral Biol. 1997, 42, 1–9. [Google Scholar] [CrossRef]
- Yurkstas, A.A. The masticatory act: A review. J. Prosthet. Dent. 1965, 15, 248–260. [Google Scholar] [CrossRef]
- Griffiths, M. Crash Course: Gastrointestinal System E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Meyer, J.H. Gastric emptying of ordinary food: Effect of antrum on particle size. Am. J. Physiol. Gastrointest. Liver Physiol. 1980, 239, G133–G135. [Google Scholar] [CrossRef] [PubMed]
- Louie, J.C.Y.; Buyken, A.E.; Brand-Miller, J.C.; Flood, V.M. The link between dietary glycemic index and nutrient adequacy. Am. J. Clin. Nutr. 2012, 95, 694–702. [Google Scholar] [CrossRef]
- Dona, A.C.; Pages, G.; Gilbert, R.G.; Kuchel, P.W. Digestion of starch: In vivo and in vitro kinetic models used to characterise oligosaccharide or glucose release. Carbohydr. Polym. 2010, 80, 599–617. [Google Scholar] [CrossRef]
- Taniguchi, H.; Honnda, Y. Amylases. In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: Oxford, UK, 2009; pp. 159–173. [Google Scholar]
- Hoebler, C.; Devaux, M.-F.; Karinthi, A.; Belleville, C.; Barry, J.-L. Particle size of solid food after human mastication and in vitro simulation of oral breakdown. Int. J. Food Sci. Nutr. 2000, 51, 353–366. [Google Scholar] [CrossRef]
- Boron, W.; Boulpaep, E.L. Medical Physiology; Elsevier: Saint Louis, MO, USA, 2016. [Google Scholar]
- Foster, T.; Adams, G.; di Bari, V.; Connerton, I.; Gould, J.; Gouseti, O.; Gray, D.; Yakubov, G. Food biotechnology. Curr. Opin. Chem. Eng. 2020, 30, 53–59. [Google Scholar] [CrossRef]
- Livesey, G.; Taylor, R.; Livesey, H.F.; Buyken, A.E.; Jenkins, D.J.; Augustin, L.S.; Sievenpiper, J.L.; Barclay, A.W.; Liu, S.; Wolever, T. Dietary glycemic index and load and the risk of type 2 diabetes: A systematic review and updated meta-analyses of prospective cohort studies. Nutrients 2019, 11, 1280. [Google Scholar] [CrossRef]
- Priebe, M.G.; Eelderink, C.; Wachters-Hagedoorn, R.E.; Vonk, R.J. Starch digestion and applications of slowly available starch. In Starch in Food; Elsevier: Amsterdam, The Netherlands, 2018; pp. 805–826. [Google Scholar]
- Lea, A.S.I. A comparative study of natural and artificial digestions. (Preliminary account.). Proc. R. Soc. Lond. 1890, 47, 192–197. [Google Scholar]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Langrish, T.A.G.; Zhong, C.; Sun, L. Probing differences in mass-transfer coefficients in beaker and stirrer digestion systems and the USP dissolution apparatus 2 using benzoic acid tablets. Processes 2021, 9, 2168. [Google Scholar] [CrossRef]
- Jones, D.; Caballero, S.; Davidov-Pardo, G. Chapter Six-Bioavailability of nanotechnology-based bioactives and nutraceuticals. In Advances in Food and Nutrition Research; Lim, L.-T., Rogers, M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 235–273. [Google Scholar]
- Le Révérend, B.J.D.; Gouseti, O.; Bakalis, S. Design Structures for Controlled Manipulation of Flavour and Texture. In Formulation Engineering of Foods; Wiley: Hoboken, NJ, USA, 2013; pp. 136–162. [Google Scholar]
- Swackhamer, C.; Bornhorst, G.M. Fracture properties of foods: Experimental considerations and applications to mastication. J. Food Eng. 2019, 263, 213–226. [Google Scholar] [CrossRef]
- Van Vliet, T. Rheology and Fracture Mechanics of Foods; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Bornhorst, G.M.; Singh, R.P. Kinetics of in vitro bread bolus digestion with varying oral and gastric digestion parameters. Food Biophys. 2013, 8, 50–59. [Google Scholar] [CrossRef]
- Chen, J. Food oral processing—A review. Food Hydrocoll. 2009, 23, 1–25. [Google Scholar] [CrossRef]
- Foster, K.D.; Woda, A.; Peyron, M.-A. Effect of texture of plastic and elastic model foods on the parameters of mastication. J. Neurophysiol. 2006, 95, 3469–3479. [Google Scholar] [CrossRef] [PubMed]
- Kohyama, K.; Sawada, H.; Nonaka, M.; Kobori, C.; Hayakawa, F.; Sasaki, T. Textural evaluation of rice cake by chewing and swallowing measurements on human subjects. Biosci. Biotechnol. Biochem. 2007, 71, 358. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, S.K.; Novotny, J.A.; Bornhorst, G.M.; Baer, D.J. Food processing and structure impact the metabolizable energy of almonds. Food Funct. 2016, 7, 4231–4238. [Google Scholar] [CrossRef]
- Read, N.W.; Welch, I.M.; Austen, C.J.; Barnish, C.; Bartlett, C.E.; Baxter, A.J.; Brown, G.; Compton, M.E.; Hume, K.E.; Storie, I.; et al. Swallowing food without chewing; a simple way to reduce postprandial glycaemia. Br. J. Nutr. 1986, 55, 43–47. [Google Scholar] [CrossRef]
- Madhu, V.; Shirali, A.; Pawaskar, P.N.; Madi, D.; Chowta, N.; Ramapuram, J.T. Mastication frequency and postprandial blood sugar levels in normoglycaemic and dysglycaemic individuals: A cross-sectional comparative study. J. Clin. Diagn. Res. 2016, 10, Oc06–Oc08. [Google Scholar] [CrossRef]
- Hoebler, C.; Karinthi, A.; Devaux, M.F.; Guillon, F.; Gallant, D.J.G.; Bouchet, B.; Melegari, C.; Barry, J.L. Physical and chemical transformations of cereal food during oral digestion in human subjects. Br. J. Nutr. 1998, 80, 429–436. [Google Scholar] [CrossRef]
- Choy, J.Y.M.; Goh, A.T.; Chatonidi, G.; Ponnalagu, S.; Wee, S.M.M.; Stieger, M.; Forde, C.G. Impact of food texture modifications on oral processing behaviour, bolus properties and postprandial glucose responses. Curr. Res. Food Sci. 2021, 4, 891–899. [Google Scholar] [CrossRef]
- Berry, E.S.; Valdes, A.M.; Drew, D.A.; Asnicar, F.; Mazidi, M.; Wolf, J.; Capdevila, J.; Hadjigeorgiou, G.; Davies, R.; Al Khatib, H.; et al. Human postprandial responses to food and potential for precision nutrition. Nat. Med. 2020, 26, 964–973. [Google Scholar] [CrossRef]
- Blanquet-Diot, S.; François, O.; Denis, S.; Hennequin, M.; Peyron, M.A. Importance of oral phase in in vitro starch digestibility related to wholegrain versus refined pastas and mastication impairment. Food Hydrocoll. 2021, 112, 106277. [Google Scholar] [CrossRef]
- Dooley, C.P.; di Lorenzo, C.; Valenzuela, J.E. Variability of migrating motor complex in humans. Dig. Dis. Sci. 1992, 37, 723–728. [Google Scholar] [CrossRef]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Bellmann, S.; Lelieveld, J.; Gorissen, T.; Minekus, M.; Havenaar, R. Development of an advanced in vitro model of the stomach and its evaluation versus human gastric physiology. Food Res. Int. 2016, 88, 191–198. [Google Scholar] [CrossRef]
- Li, C.; Yu, W.; Wu, P.; Chen, X.D. Current in vitro digestion systems for understanding food digestion in human upper gastrointestinal tract. Trends Food Sci. Technol. 2020, 96, 114–126. [Google Scholar] [CrossRef]
- Hayes, A.M.R.; Swackhamer, C.; Mennah-Govela, Y.A.; Martinez, M.M.; Diatta, A.; Bornhorst, G.M.; Hamaker, B.R. Pearl millet (Pennisetum glaucum) couscous breaks down faster than wheat couscous in the Human Gastric Simulator, though has slower starch hydrolysis. Food Funct. 2020, 11, 111–122. [Google Scholar] [CrossRef]
- Van de Wiele, T.; van den Abbeele, P.; Ossieur, W.; Possemiers, S.; Marzorati, M. The Simulator of the Human Intestinal Microbial Ecosystem (SHIME(®)). In The Impact of Food Bioactives on Health; Springer: Berlin/Heidelberg, Germany, 2015; pp. 305–317. [Google Scholar]
- Molly, K.; Woestyne, M.V.; Verstraete, W. Development of a 5-step multi-chamber reactor as a simulation of the human intestinal microbial ecosystem. Appl. Microbiol. Biotechnol. 1993, 39, 254–258. [Google Scholar] [CrossRef]
- Minekus, M.; Smeets-Peeters, M.; Bernalier, A.; Marol-Bonnin, S.; Havenaar, R.; Marteau, P.; Alric, M.; Fonty, G.; Veld, J.H.H.i. A computer-controlled system to simulate conditions of the large intestine with peristaltic mixing, water absorption and absorption of fermentation products. Appl. Microbiol. Biotechnol. 1999, 53, 108–114. [Google Scholar] [CrossRef]
- Chen, X.D. GIT physicochemical modeling-a critical review. Int. J. Food Eng. 2006, 2, 367–371. [Google Scholar] [CrossRef]
- Agatonovic-Kustrin, S.; Beresford, R. Basic concepts of artificial neural network (ANN) modeling and its application in pharmaceutical research. J. Pharm. Biomed. Anal. 2000, 22, 717–727. [Google Scholar] [CrossRef]
- Goddard, M.S.; Young, G.; Marcus, R. The effect of amylose content on insulin and glucose responses to ingested rice. Am. J. Clin. Nutr. 1984, 39, 388–392. [Google Scholar] [CrossRef]
- Granfeldt, Y.; Liljeberg, H.; Drews, A.; Newman, R.; Björck, I. Glucose and insulin responses to barley products: Influence of food structure and amylose-amylopectin ratio. Am. J. Clin. Nutr. 1994, 59, 1075–1082. [Google Scholar] [CrossRef]
- Hallfrisch, J.; Behall, K.M. Mechanisms of the effects of grains on insulin and glucose responses. J. Am. Coll. Nutr. 2000, 19 (Suppl. S3), 320S–325S. [Google Scholar] [CrossRef]
- Kristensen, M.; Jensen, M.G.; Riboldi, G.; Petronio, M.; Bügel, S.; Toubro, S.; Tetens, I.; Astrup, A. Wholegrain vs. refined wheat bread and pasta. effect on postprandial glycemia, appetite, and subsequent ad libitum energy intake in young healthy adults. Appetite 2010, 54, 163–169. [Google Scholar] [CrossRef]
- Peyrot des Gachons, C.; Breslin, P.A.S. Salivary amylase: Digestion and metabolic syndrome. Curr. Diabetes Rep. 2016, 16, 102. [Google Scholar] [CrossRef]
- Csorba, T.R.; Matsuda, I.; Kalant, N. Effects of insulin and diabetes on flux rates of plasma glucose and free fatty acids. Metabolism 1966, 15, 262–270. [Google Scholar] [CrossRef]
- Woda, A.; Mishellany-Dutour, A.; Batier, L.; François, O.; Meunier, J.P.; Reynaud, B.; Alric, M.; Peyron, M.A. Development and validation of a mastication simulator. J. Biomech. 2010, 43, 1667–1673. [Google Scholar] [CrossRef]
- Salles, C.; Tarrega, A.; Mielle, P.; Maratray, J.; Gorria, P.; Liaboeuf, J.; Liodenot, J.J. Developmentof a chewing simulator for food breakdown and the analysis of in vitro flavor compound release in a mouth environment. J. Food Eng. 2007, 82, 189–198. [Google Scholar] [CrossRef]
- Harrison, S.M.; Cleary, P.W.; Eyres, G.; Sinnott, M.D.; Lundin, L. Challenges in computational modelling of food breakdown and flavour release. Food Funct. 2014, 5, 2792–2805. [Google Scholar] [CrossRef]
- Bellmann, S.; Minekus, M.; Zeijdener, E.; Verwei, M.; Sanders, P.; Basten, W.; Havenaar, R. TIM-carbo: A rapid, cost-efficient and reliable in vitro method for glycaemic response after carbohydrate ingestion. In Dietary Fibre: New Frontiers for Food and Health; Wageningen Academic: Wageningen, The Netherlands, 2010; pp. 467–473. [Google Scholar]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Thuenemann, E.C.; Mandalari, G.; Rich, G.T.; Faulks, R.M. Dynamic Gastric Model (DGM). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 47–59. [Google Scholar]
- Ballance, S.; Sahlstrøm, S.; Lea, P.; Nagy, N.E.; Andersen, P.V.; Dessev, T.; Hull, S.; Vardakou, M.; Faulks, R. Evaluation of gastric processing and duodenal digestion of starch in six cereal meals on the associated glycaemic response using an adult fasted dynamic gastric model. Eur. J. Nutr. 2013, 52, 799–812. [Google Scholar] [CrossRef]
- Moxon, T.E.; Gouseti, O.; Bakalis, S. In silico modelling of mass transfer & absorption in the human gut. J. Food Eng. 2016, 176, 110–120. [Google Scholar]
- Ye, Q.; Ge, F.; Wang, Y.; Wu, P.; Chen, X.D.; Selomulya, C. Digestion of curcumin-fortified yogurt in short/long gastric residence times using a near-real dynamic in vitro human stomach. Food Chem. 2022, 372, 131327. [Google Scholar] [CrossRef]
- Iqbal, S.; Zhang, P.; Wu, P.; Ge, A.; Ge, F.; Deng, R.; Chen, X.D. Evolutions of rheology, microstructure and digestibility of parboiled rice during simulated semi-dynamic gastrointestinal digestion. LWT 2021, 148, 111700. [Google Scholar] [CrossRef]
- Dhital, S.; Warren, F.J.; Butterworth, P.J.; Ellis, P.R.; Gidley, M.J. Mechanisms of starch digestion by α-amylase—Structural basis for kinetic properties. Crit. Rev. Food Sci. Nutr. 2017, 57, 875–892. [Google Scholar] [CrossRef]
- Gouseti, O.; Jaime-Fonseca, M.R.; Fryer, P.J.; Mills, C.; Wickham, M.S.J.; Bakalis, S. Hydrocolloids in human digestion: Dynamic in-vitro assessment of the effect of food formulation on mass transfer. Food Hydrocoll. 2014, 42, 378–385. [Google Scholar] [CrossRef]
- Mackie, A.; Mulet-Cabero, A.-I.; Torcello-Gómez, A. Simulating human digestion: Developing our knowledge to create healthier and more sustainable foods. Food Funct. 2020, 11, 9397–9431. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, M.J.; Rosenmund, H. The action of human pancreatic and salivary isoamylases on starch and glycogen. Clin. Chim. Acta 1977, 79, 69–73. [Google Scholar] [CrossRef]
- Prinz, J.; Janssen, A.; de Wijk, R. In vitro simulation of the oral processing of semi-solid foods. Food Hydrocoll. 2007, 21, 397–401. [Google Scholar] [CrossRef]
- Prinz, J.; de Wijk, R.; Huntjens, L. Load dependency of the coefficient of friction of oral mucosa. Food Hydrocoll. 2007, 21, 402–408. [Google Scholar] [CrossRef]
- Humphrey, S.P.; Williamson, R.T. A review of saliva: Normal composition, flow, and function. J. Prosthet. Dent. 2001, 85, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Fernández, C.I.; Wiley, A.S. Rethinking the starch digestion hypothesis for AMY1 copy number variation in humans. Am. J. Phys. Anthropol. 2017, 163, 645–657. [Google Scholar] [CrossRef]
- Feher, J.J. Quantitative Human Physiology: An Introduction; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Cussler, E.L. Diffusion: Mass Transfer in Fluid Systems, 3rd ed; Cambridge University Press: New York, NY, USA, 2009. [Google Scholar]
- Tharakan, A.; Norton, I.T.; Fryer, P.J.; Bakalis, S. Mass transfer and nutrient absorption in a simulated model of small intestine. J. Food Sci. 2010, 75, E339–E346. [Google Scholar] [CrossRef]
- Lamond, A.R.; Janssen, A.E.M.; Mackie, A.; Bornhorst, G.M.; Bakalis, S.; Gouseti, O. An engineering perspective on human digestion. In Interdisciplinary Approaches to Food Digestion; Gouseti, O., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 255–273. [Google Scholar]
- Sherwood, T.K.; Pigford, R.L.; Wilke, C.R. Mass Transfer; McGraw-Hill Book Company: New York, NY, USA, 1975; 677p. [Google Scholar]
- Reynolds, O. XXIX. An experimental investigation of the circumstances which determine whether the motion of water shall be direct or sinuous, and of the law of resistance in parallel channels. Philos. Trans. R. Soc. Lond. 1883, 174, 935–982. [Google Scholar]
- Geankoplis, C. Transport Processes and Unit Operations; Allyn and Bacon: Newton, MA, USA, 1995. [Google Scholar]
- Catchpole, J.P.; Fulford, G. Dimensionless groups. Ind. Eng. Chem. 1966, 58, 46–60. [Google Scholar] [CrossRef]
- Baks, T.; Bruins, M.E.; Matser, A.M.; Janssen, A.E.M.; Boom, R.M. Effect of gelatinization and hydrolysis conditions on the selectivity of starch hydrolysis with α-amylase from bacillus licheniformis. J. Agric. Food Chem. 2008, 56, 488–495. [Google Scholar] [CrossRef]
- Yu, F.; Hunt, A.G. Damköhler number input to transport-limited chemical weathering calculations. ACS Earth Space Chem. 2017, 1, 30–38. [Google Scholar] [CrossRef]
- Çalık, P.; Çalık, G.; Özdamar, T.H. Oxygen-transfer strategy and its regulation effects in serine alkaline protease production by Bacillus licheniformis. Biotechnol. Bioeng. 2000, 69, 301–311. [Google Scholar] [CrossRef]
- Van Daele, T.; del Pozo, D.F.; van Hauwermeiren, D.; Gernaey, K.V.; Wohlgemuth, R.; Nopens, I. A generic model-based methodology for quantification of mass transfer limitations in microreactors. Chem. Eng. J. 2016, 300, 193–208. [Google Scholar] [CrossRef]
- Del Pozo, D.F.; Hauwermeiren, D.V. Effect of mass transfer limitations on enzymatic reactions in microreactors: A model-based analysis. Chem. Eng. J. 2016, 300, 193–208. [Google Scholar]
- Lewandowski, M.T.; Parente, A.; Pozorski, J. Generalised eddy dissipation concept for mild combustion regime at low local reynolds and damköhler numbers. Part 1: Model framework development. Fuel 2020, 278, 117743. [Google Scholar] [CrossRef]
- Manickam, M.; Schwarz, M.P.; Perry, J. CFD modelling of waste heat recovery boiler. Appl. Math. Model. 1998, 22, 823–840. [Google Scholar] [CrossRef]
- Wild, P.N.; FaltsiSaravelou, O. Mathematical modelling of a 2.25 MW(t) swirling natural gas flame 2. Conserved scalar approach for turbulent combustion. Combust. Sci. Technol. 1995, 111, 103–121. [Google Scholar] [CrossRef]
- Klein, S.A.; Kok, J.B.W. Sound generation by turbulent non-premixed flames. Combust. Sci. Technol. 1999, 149, 267–295. [Google Scholar] [CrossRef]
- Tyagi, M.; Jamadar, N.; Chakravarthy, S.R. Oscillatory response of an idealized two-dimensional diffusion flame: Analytical and numerical study. Combust. Flame 2007, 149, 271–285. [Google Scholar] [CrossRef]
- Goñi, I.; Garcia-Alonso, A.; Saura-Calixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]
- Butterworth, P.J.; Warren, F.J.; Ellis, P.R. Human α-amylase and starch digestion: An interesting marriage. Starch Stärke 2011, 63, 395–405. [Google Scholar] [CrossRef]
- Liu, X.; Li, L.; Yu, J.; Copeland, L.; Wang, S.; Wang, S. In vitro digestibility of starches with different crystalline polymorphs at low α-amylase activity to substrate ratio. Food Chem. 2021, 349, 129170. [Google Scholar] [CrossRef] [PubMed]
- Dhital, S.; Dolan, G.; Stokes, J.R.; Gidley, M.J. Enzymatic hydrolysis of starch in the presence of cereal soluble fibre polysaccharides. Food Funct. 2014, 5, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Phillips, L.K.; Deane, A.M.; Jones, K.L.; Rayner, C.K.; Horowitz, M. Gastric emptying and glycaemia in health and diabetes mellitus. Nat. Rev. Endocrinol. 2015, 11, 112–128. [Google Scholar] [CrossRef]
- Fonte, P.; Araújo, F.; Reis, S.; Sarmento, B. Oral insulin delivery: How far are we? J. Diabetes Sci. Technol. 2013, 7, 520–531. [Google Scholar] [CrossRef]
- Qin, Y.; Xiao, J.; Wang, Y.; Dong, Z.; Woo, M.W.; Chen, X.D. Mechanistic exploration of glycemic lowering by soluble dietary fiber ingestion: Predictive modeling and simulation. Chem. Eng. Sci. 2020, 228, 115965. [Google Scholar] [CrossRef]
- Feher, J. 8.5-Digestion and Absorption of the Macronutrients. In Quantitative Human Physiology; Feher, J., Ed.; Academic Press: Boston, MA, USA, 2012; pp. 731–743. [Google Scholar]


{kind=link}
{kind=link}
| Key Steps | Test Design | Reference |
|---|---|---|
| Participants | More than 10 people; No known food allergy; 18–35 years old; Non-smokers; Healthy (8 data/range of criteria). | [24] |
| Test samples | Reference food: 50 g glucose; Test food: 50 g carbohydrate containing; 250 mL water served. | [22] |
| Blood samples | Take blood samples at −10, −5, 15, 30, 45, 60, 90, 120 min. | [22] |
| Data analysis | Spectrophotometry or electrochemical detection-coupled enzyme systems. | [25] |
| In Vitro Digestion Model for Starch Digestion | How It Works | References | ||
|---|---|---|---|---|
| Oral models: | AM2 | Physical model | Simulate human masticatory behavior and generate a food bolus with similar granulometric characteristics. | [106,107] |
| B-SPH | Mathematical model | Simulate several complicated aspects of mastication. | [108] | |
| Gastrointestinal models: | TIM-Carbo | Physical model | Test the digestibility of carbohydrates through the stomach and small intestine. Use the availability of monosaccharides to predict glycemic response. HOMA for Data process. | [109,110] |
| DGM | Physical model | Monitor the changes in the ratio of glucose:starch and total starch effect of particle size on starch hydrolysis from Durum wheat. | [111,112] | |
| CSTR PFR | Physical model | Investigate the digestion and absorption of starch and glucose in the small intestine. Understand the relative effect of gastric emptying time and luminal viscosity on the rate of glucose absorption. | [113] | |
| Dimensionless Group | Equation | Physical Meaning | Description | Ref. |
|---|---|---|---|---|
| Sherwood | k = mass transfer coefficient (m/s); l = length (m); D = mass diffusivity (m2/s). | Mass transfer | [128] | |
| Reynolds | ρ = density (kg/m3); u = velocity (m/s); l = length (m); µ = viscosity (Pa.s). | Forced convection, fluid mechanics, heat transfer and mass transfer | [129] | |
| Schmidt | Symbols defined above. | Fluid mechanics and mass transfer | [130] | |
| Damköhler | kr = reaction rate constant (mol1−n m3(1−n)/s); Co = initial concentration (mol/m3); n = reaction order; τ = mean residence time (s). | Reaction engineering and mass transfer | [131] |
| 5% (w/w) Wheat Starch Solution | Reaction Rate Constant (min−1) | Da |
|---|---|---|
| Amylase/Starch ratio (% w/w): 0.1 | 0.003 | 0.19 |
| Amylase/Starch ratio (% w/w): 1.0 | 0.011 | 0.65 |
| Amylase/Starch ratio (% w/w): 10 | 0.007 | 0.45 |
| Potato Starch Solution (about 20% w/w) | Reaction Rate Constant (min−1) | Da |
|---|---|---|
| Degree of gelatinization (%): 100 | 0.0496 | 2.9 |
| Degree of gelatinization (%): 34 | 0.0198 | 1.2 |
| Degree of gelatinization (%): 0 | 0.0118 | 0.7 |
| Fraction of Glucose Absorbed in the Intestine Model Simulation | Reaction Rate (min−1) | Da |
|---|---|---|
| 0.2 | 2 | 0.04 |
| 0.6 | 7 | 0.14 |
| 0.8 | 14 | 0.28 |
| 1.0 | 25 | 0.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Zhong, C.; Zhou, Z.; Lei, Z.; Langrish, T.A.G. A Review of In Vitro Methods for Measuring the Glycemic Index of Single Foods: Understanding the Interaction of Mass Transfer and Reaction Engineering by Dimensional Analysis. Processes 2022, 10, 759. https://doi.org/10.3390/pr10040759
Sun Y, Zhong C, Zhou Z, Lei Z, Langrish TAG. A Review of In Vitro Methods for Measuring the Glycemic Index of Single Foods: Understanding the Interaction of Mass Transfer and Reaction Engineering by Dimensional Analysis. Processes. 2022; 10(4):759. https://doi.org/10.3390/pr10040759
Chicago/Turabian StyleSun, Yongmei, Chao Zhong, Zelin Zhou, Zexin Lei, and Timothy A. G. Langrish. 2022. "A Review of In Vitro Methods for Measuring the Glycemic Index of Single Foods: Understanding the Interaction of Mass Transfer and Reaction Engineering by Dimensional Analysis" Processes 10, no. 4: 759. https://doi.org/10.3390/pr10040759
APA StyleSun, Y., Zhong, C., Zhou, Z., Lei, Z., & Langrish, T. A. G. (2022). A Review of In Vitro Methods for Measuring the Glycemic Index of Single Foods: Understanding the Interaction of Mass Transfer and Reaction Engineering by Dimensional Analysis. Processes, 10(4), 759. https://doi.org/10.3390/pr10040759