
Airway Epithelial Cultures of Children with Esophageal Atresia as a Model to Study Respiratory Tract Disorders

 , , , , ,
, , , , ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Materials and Informed Consent
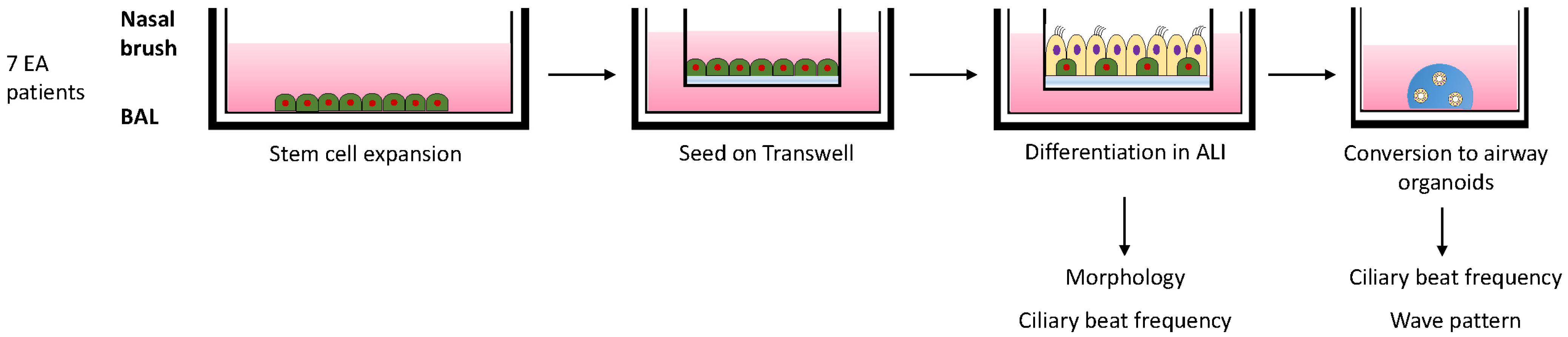
2.2. Isolation and Differentiation of Airway Cells
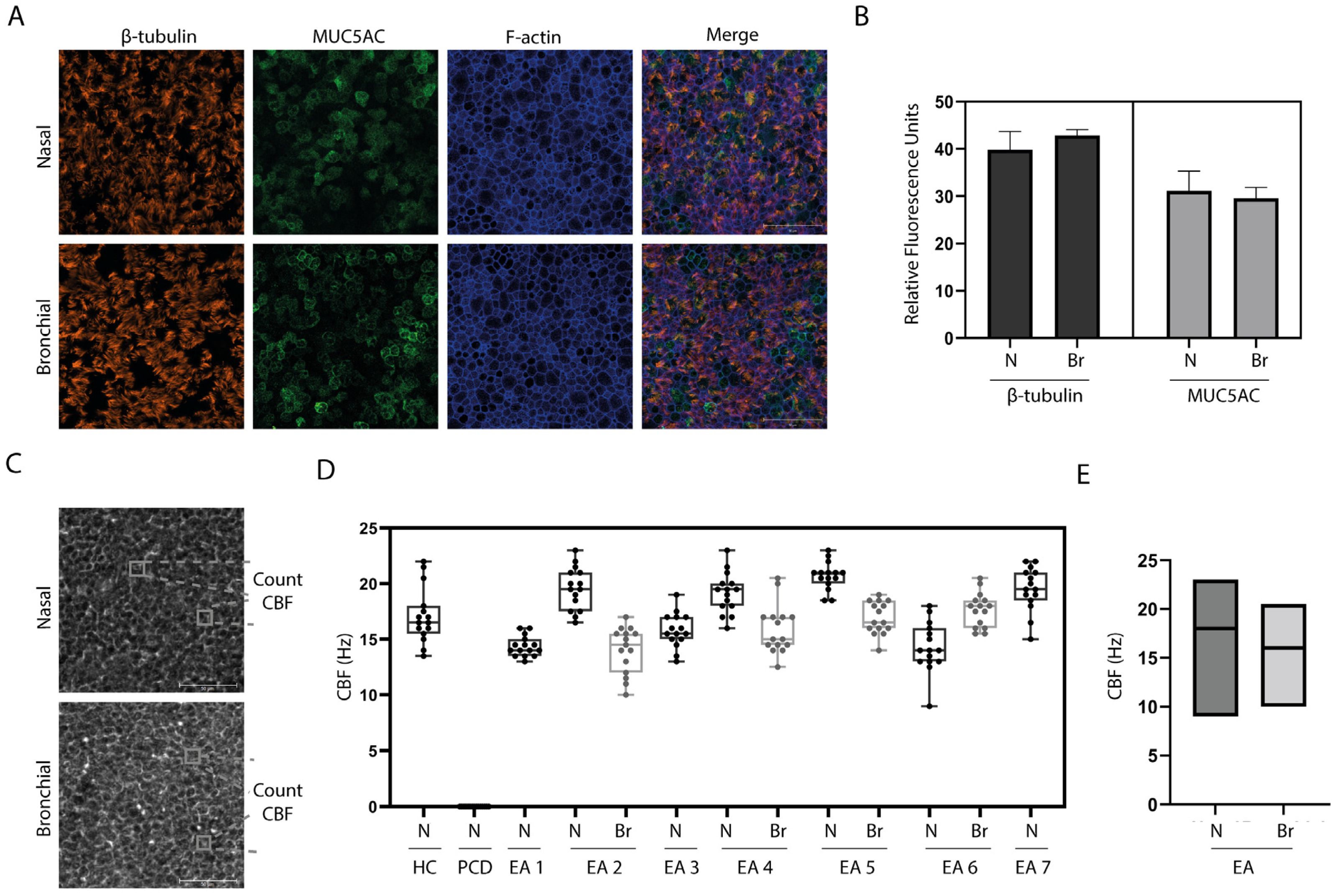
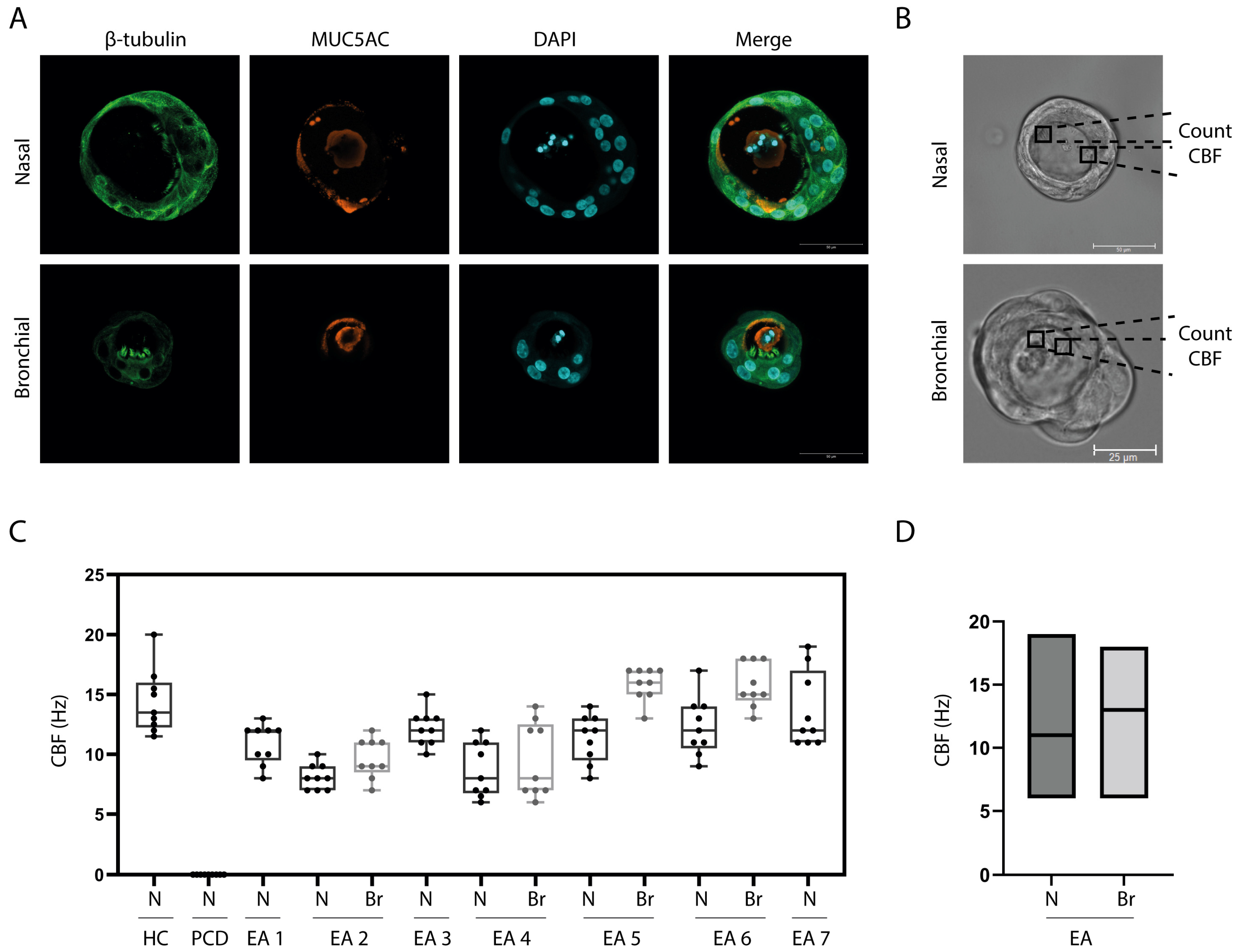
2.3. Fixation and Immunofluorescent Microscopy
2.4. Analysis of Ciliary Beat Frequency (CBF)
3. Results
3.1. Characteristics of Patients with Esophageal Atresia
3.2. In Vitro Differentiation of EA Patient Derived Airway Cultures
3.3. Ciliary Activity
4. Discussion

{kind=link}
{kind=link}
{kind=link}
| Isolation Technique | Risk for Donor | Respiratory Model | Literature |
|---|---|---|---|
| Leftover tissue from operation | Not invasive | Cigarette smoke, lung cancer, Rhinovirus, RSV, SARS- CoV-2 | [16,17,20,23,28] |
| Tracheal aspirates | Not invasive | Cigarette smoke, homeostasis | [18,23] |
| Nasal brush/wash | Minimal invasive | Asthma, CF, COPD, PCD, RSV, SARS-CoV-2 | [15,21,24,27,29,30,31] |
| Bronchial biopsy/brushing | Medium invasive | Asthma, COPD, PCD, Rhinovirus, SARS-CoV-2 | [27,32,33,34,35] |
| BAL | Medium invasive | Asthma, lung cancer, RSV | [16,22,27] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chetcuti, P.; Phelan, P.D. Respiratory Morbidity after Repair of Oesophageal Atresia and Tracheo-Oesophageal Fistula. Arch. Dis. Child 1993, 68, 167–170. [Google Scholar] [CrossRef]
- Cartabuke, R.H.; Lopez, R.; Thota, P.N. Long-Term Esophageal and Respiratory Outcomes in Children with Esophageal Atresia and Tracheoesophageal Fistula. Gastroenterol. Rep. 2016, 4, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Sistonen, S.J.; Pakarinen, M.P.; Rintala, R.J. Long-Term Results of Esophageal Atresia: Helsinki Experience and Review of Literature. Pediatr. Surg. Int. 2011, 27, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, I.; Beardsmore, C.S.; MacFadysen, U.M. Respiratory Function in Childhood Following Repair of Oesophageal Atresia and Tracheoesophageal Fistula. Arch. Dis. Child. 1999, 81, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Spoel, M.; Meeussen, C.J.H.M.; Gischler, S.J.; Hop, W.C.J.; Bax, N.M.A.; Wijnen, R.M.H.; Tibboel, D.; de Jongste, J.C.; Ijsselstijn, H. Respiratory Morbidity and Growth after Open Thoracotomy or Thoracoscopic Repair of Esophageal Atresia. J. Pediatr. Surg. 2012, 47, 1975–1983. [Google Scholar] [CrossRef] [PubMed]
- Sistonen, S.; Malmberg, P.; Malmström, K.; Haahtela, T.; Sarna, S.; Rintala, R.J.; Pakarinen, M.P. Repaired Oesophageal Atresia: Respiratory Morbidity and Pulmonary Function in Adults. Eur. Respir. J. 2010, 36, 1106–1112. [Google Scholar] [CrossRef]
- Donoso, F.; Hedenström, H.; Malinovschi, A.; Lilja, H.E. Pulmonary Function in Children and Adolescents after Esophageal Atresia Repair. Pediatr. Pulmonol. 2020, 55, 206–213. [Google Scholar] [CrossRef]
- Emery, J.L.; Haddadin, A.J. Squamous Epithelium in the Respiratory Tract of Children with Tracheo-Oesophageal Fistula, and “Retention Lung”. Arch. Dis. Child. 1971, 46, 884. [Google Scholar] [CrossRef]
- Wailoo, M.P.; Emery, J.L. The Trachea in Children with Tracheo-oesophageal Fistula. Histopathology 1979, 3, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Hackett, T.L.; Singhera, G.K.; Shaheen, F.; Hayden, P.; Jackson, G.R.; Hegele, R.G.; van Eeden, S.; Bai, T.R.; Dorscheid, D.R.; Knight, D.A. Intrinsic Phenotypic Differences of Asthmatic Epithelium and Its Inflammatory Responses to Respiratory Syncytial Virus and Air Pollution. Am. J. Respir. Cell Mol. Biol. 2011, 45, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Rao, W.; Wang, S.; Duleba, M.; Niroula, S.; Goller, K.; Xie, J.; Mahalingam, R.; Neupane, R.; Liew, A.A.; Vincent, M.; et al. Regenerative Metaplastic Clones in COPD Lung Drive Inflammation and Fibrosis. Cell 2020, 181, 848–864.e18. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Miller, Y.E.; Nakachi, I.; Kwon, J.B.; Barón, A.E.; Brantley, A.E.; Merrick, D.T.; Franklin, W.A.; Keith, R.L.; Vandivier, R.W. Exhaustion of Airway Basal Progenitor Cells in Early and Established Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2018, 197, 885–896. [Google Scholar] [CrossRef]
- Edwards, N.A.; Shacham-Silverberg, V.; Weitz, L.; Kingma, P.S.; Shen, Y.; Wells, J.M.; Chung, W.K.; Zorn, A.M. Developmental Basis of Trachea-Esophageal Birth Defects. Dev. Biol. 2021, 477, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.A.; Hever, A.M.; Rainger, J.; Rogers, R.C.; Magee, A.; Fiedler, Z.; Keng, W.T.; Sharkey, F.H.; McGill, N.; Hill, C.J.; et al. Mutations in SOX2 Cause Anophthalmia-Esophageal-Genital (AEG) Syndrome. Hum. Mol. Genet. 2006, 15, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Amatngalim, G.D.; Rodenburg, L.W.; Aalbers, B.L.; Raeven, H.H.M.; Aarts, E.M.; Sarhane, D.; Spelier, S.; Lefferts, J.W.; Silva, I.A.L.; Nijenhuis, W.; et al. Measuring Cystic Fibrosis Drug Responses in Organoids Derived from 2D Differentiated Nasal Epithelia. Life Sci. Alliance 2022, 5, e202101320. [Google Scholar] [CrossRef]
- Sachs, N.; Papaspyropoulos, A.; Zomer-van Ommen, D.D.; Heo, I.; Böttinger, L.; Klay, D.; Weeber, F.; Huelsz-Prince, G.; Iakobachvili, N.; Amatngalim, G.D.; et al. Long-term Expanding Human Airway Organoids for Disease Modeling. EMBO J. 2019, 38, e100300. [Google Scholar] [CrossRef] [PubMed]
- Mou, H.; Vinarsky, V.; Tata, P.R.; Brazauskas, K.; Choi, S.H.; Crooke, A.K.; Zhang, B.; Solomon, G.M.; Turner, B.; Bihler, H.; et al. Dual SMAD Signaling Inhibition Enables Long-Term Expansion of Diverse Epithelial Basal Cells. Cell Stem. Cell 2016, 19, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhu, X.; Shui, J.E.; Xiong, L.; Gierahn, T.; Zhang, C.; Wood, M.; Hally, S.; Love, J.C.; Li, H.; et al. Rho/SMAD/MTOR Triple Inhibition Enables Long-Term Expansion of Human Neonatal Tracheal Aspirate-Derived Airway Basal Cell-like Cells. Pediatr. Res. 2021, 89, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Paplinska-Goryca, M.; Misiukiewicz-Stepien, P.; Nejman-Gryz, P.; Proboszcz, M.; Mlacki, M.; Gorska, K.; Krenke, R. Epithelial-Macrophage-Dendritic Cell Interactions Impact Alarmins Expression in Asthma and COPD. Clin. Immunol. 2020, 215, 108421. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef] [PubMed]
- Chiu, M.C.; Li, C.; Liu, X.; Song, W.; Wan, Z.; Yu, Y.; Huang, J.; Xiao, D.; Chu, H.; Cai, J.P.; et al. Human Nasal Organoids Model SARS-CoV-2 Upper Respiratory Infection and Recapitulate the Differential Infectivity of Emerging Variants. mBio 2022, 13, e01944-22. [Google Scholar] [CrossRef] [PubMed]
- Eenjes, E.; van Riet, S.; Kroon, A.A.; Slats, A.M.; Khedoe, P.P.S.J.; Boerema-De Munck, A.; Buscop-Van Kempen, M.; Ninaber, D.K.; Reiss, I.K.M.; Clevers, H.; et al. Disease Modeling Following Organoid-Based Expansion of Airway Epithelial Cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 321, L775–L786. [Google Scholar] [CrossRef] [PubMed]
- Salahudeen, A.A.; Choi, S.S.; Rustagi, A.; Zhu, J.; van Unen, V.; de la O, S.M.; Flynn, R.A.; Margalef-Català, M.; Santos, A.J.M.; Ju, J.; et al. Progenitor Identification and SARS-CoV-2 Infection in Human Distal Lung Organoids. Nature 2020, 588, 670–675. [Google Scholar] [CrossRef]
- Rajan, A.; Weaver, A.M.; Aloisio, G.M.; Jelinski, J.; Johnson, H.L.; Venable, S.F.; Mcbride, T.; Aideyan, L.; Piedra, F.-A.; Ye, X.; et al. The Human Nose Organoid Respiratory Virus Model: An Ex Vivo Human Challenge Model To Study Respiratory Syncytial Virus (RSV) and Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Pathogenesis and Evaluate Therapeutics. Mbio 2023, 13, e03511-21. [Google Scholar] [CrossRef] [PubMed]
- Gohy, S.T.; Detry, B.R.; Lecocq, M.; Bouzin, C.; Weynand, B.A.; Amatngalim, G.D.; Sibille, Y.M.; Pilette, C. Polymeric Immunoglobulin Receptor Down-Regulation in Chronic Obstructive Pulmonary Disease: Persistence in the Cultured Epithelium and Role of Transforming Growth Factor-β. Am. J. Respir. Crit. Care Med. 2014, 190, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Engesaeth, V.G.; Warner, J.O.; Bush, A. New Associations of Primary Ciliary Dyskinesia Syndrome. Pediatr. Pulmonol. 1993, 16, 9–12. [Google Scholar] [CrossRef]
- Nagasaki, T.; Schuyler, A.J.; Zhao, J.; Samovich, S.N.; Yamada, K.; Deng, Y.; Ginebaugh, S.P.; Christenson, S.A.; Woodruff, P.G.; Fahy, J.V.; et al. 15LO1 Dictates Glutathione Redox Changes in Asthmatic Airway Epithelium to Worsen Type 2 Inflammation. J. Clin. Investig. 2022, 132, e151685. [Google Scholar] [CrossRef] [PubMed]
- Saito, N.; Kikuchi, A.; Yamaya, M.; Deng, X.; Sugawara, M.; Takayama, S.; Nagatomi, R.; Ishii, T. Kakkonto Inhibits Cytokine Production Induced by Rhinovirus Infection in Primary Cultures of Human Nasal Epithelial Cells. Front. Pharm. 2021, 12, 687818. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.D.H.; Petris, A.; Hynds, R.E.; O’Callaghan, C. Ciliated Epithelial Cell Differentiation at Air-Liquid Interface Using Commercially Available Culture Media. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2020; Volume 2109, pp. 275–291. [Google Scholar]
- Lee, D.D.H.; Cardinale, D.; Nigro, E.; Butler, C.R.; Rutman, A.; Fassad, M.R.; Hirst, R.A.; Moulding, D.; Agrotis, A.; Forsythe, E.; et al. Higher Throughput Drug Screening for Rare Respiratory Diseases: Readthrough Therapy in Primary Ciliary Dyskinesia. Eur. Respir. J. 2021, 58, 2000455. [Google Scholar] [CrossRef]
- Coles, J.L.; Thompson, J.; Horton, K.L.; Hirst, R.A.; Griffin, P.; Williams, G.M.; Goggin, P.; Doherty, R.; Lackie, P.M.; Harris, A.; et al. A Revised Protocol for Culture of Airway Epithelial Cells as a Diagnostic Tool for Primary Ciliary Dyskinesia. J. Clin. Med. 2020, 9, 3753. [Google Scholar] [CrossRef] [PubMed]
- Ravi, A.; Chowdhury, S.; Dijkhuis, A.; Dierdorp, B.S.; Dekker, T.; Kruize, R.; Piñeros, Y.S.S.; Majoor, C.J.; Sterk, P.J.; Lutter, R. Imprinting of Bronchial Epithelial Cells upon in Vivo Rhinovirus Infection in People with Asthma. ERJ Open Res. 2022, 8, 00522-2021. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guerra, S.; Ledford, J.G.; Kraft, M.; Li, H.; Hastie, A.T.; Castro, M.; Denlinger, L.C.; Erzurum, S.C.; Fahy, J.V.; et al. Low CC16 MRNA Expression Levels in Bronchial Epithelial Cells Are Associated with Asthma Severity. Am. J. Respir. Crit. Care Med. 2023, 207, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Radzikowska, U.; Ding, M.; Tan, G.; Zhakparov, D.; Peng, Y.; Wawrzyniak, P.; Wang, M.; Li, S.; Morita, H.; Altunbulakli, C.; et al. Distribution of ACE2, CD147, CD26, and Other SARS-CoV-2 Associated Molecules in Tissues and Immune Cells in Health and in Asthma, COPD, Obesity, Hypertension, and COVID-19 Risk Factors. Allergy Eur. J. Allergy Clin. Immunol. 2020, 75, 2829–2845. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Chen, L.; Guo, J.; Shi, F.; Yang, Q.; Xie, L.; Lu, D.; Li, Y.; Luo, J.; Wang, L.; et al. Multiomics Analysis of a DNAH5-Mutated PCD Organoid Model Revealed the Key Role of the TGF-β/BMP and Notch Pathways in Epithelial Differentiation and the Immune Response in DNAH5-Mutated Patients. Cells 2022, 11, 4013. [Google Scholar] [CrossRef]



| Gender, Age | Type EA | Associated Anomalies | Tracheomalacia | Respiratory History |
|---|---|---|---|---|
| 1, female, 6.4 months | C | VACTERL | Yes, severe | No infections, mild respiratory symptoms |
| 2, male, 4 days | A | Macrocefalie | None | No symptoms |
| 3, male, 1.6 years | C | None | Yes, mild | Mild respiratory symptoms, cough |
| 4, male, 4.8 years | C | None | Yes, mild | Multiple RTIs, cough |
| 5, male, 1.2 months | C | VACTERL | Yes, mild | Mild respiratory symptoms, cough |
| 6, female, 1 year | C | VACTERL | Yes, moderate | PPT, Cough |
| 7, female, 7 days | A | Hypoplastic rib | None | Mild |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dreyer, H.H.M.; van Tuyll van Serooskerken, E.S.; Rodenburg, L.W.; Bittermann, A.J.N.; Arets, H.G.M.; Reuling, E.M.B.P.; Verweij, J.W.; Haarman, E.G.; van der Zee, D.C.; Tytgat, S.H.A.J.; et al. Airway Epithelial Cultures of Children with Esophageal Atresia as a Model to Study Respiratory Tract Disorders. Children 2023, 10, 1020. https://doi.org/10.3390/children10061020
Dreyer HHM, van Tuyll van Serooskerken ES, Rodenburg LW, Bittermann AJN, Arets HGM, Reuling EMBP, Verweij JW, Haarman EG, van der Zee DC, Tytgat SHAJ, et al. Airway Epithelial Cultures of Children with Esophageal Atresia as a Model to Study Respiratory Tract Disorders. Children. 2023; 10(6):1020. https://doi.org/10.3390/children10061020
Chicago/Turabian StyleDreyer, Henriette H. M., Eleonora Sofie van Tuyll van Serooskerken, Lisa W. Rodenburg, Arnold J. N. Bittermann, Hubertus G. M. Arets, Ellen M. B. P. Reuling, Johannes W. Verweij, Eric G. Haarman, David C. van der Zee, Stefaan H. A. J. Tytgat, and et al. 2023. "Airway Epithelial Cultures of Children with Esophageal Atresia as a Model to Study Respiratory Tract Disorders" Children 10, no. 6: 1020. https://doi.org/10.3390/children10061020
APA StyleDreyer, H. H. M., van Tuyll van Serooskerken, E. S., Rodenburg, L. W., Bittermann, A. J. N., Arets, H. G. M., Reuling, E. M. B. P., Verweij, J. W., Haarman, E. G., van der Zee, D. C., Tytgat, S. H. A. J., van der Ent, C. K., Beekman, J. M., Amatngalim, G. D., & Lindeboom, M. Y. A. (2023). Airway Epithelial Cultures of Children with Esophageal Atresia as a Model to Study Respiratory Tract Disorders. Children, 10(6), 1020. https://doi.org/10.3390/children10061020