
Pan-Cancer Analysis Reveals Common and Specific Relationships between Intragenic miRNAs and Their Host Genes


Abstract
:1. Introduction
2. Materials and Methods
2.1. mRNA and miRNA Sequencing Datasets
2.2. miRNA Classification
2.3. Analysis of Transcriptional Association between miRNAs and Host Genes across Cancer Types
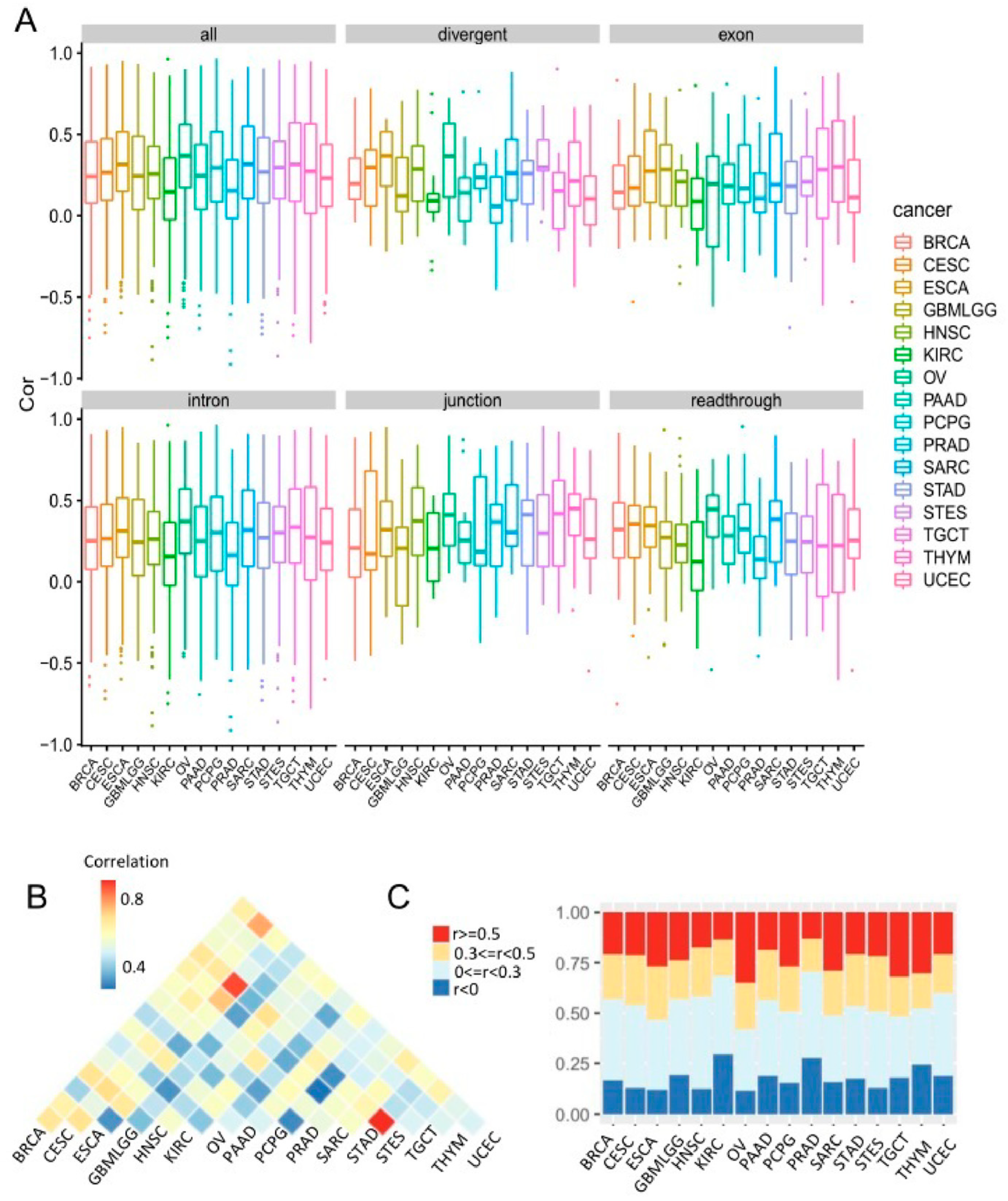
3. Results
3.1. Human miRNA Classification
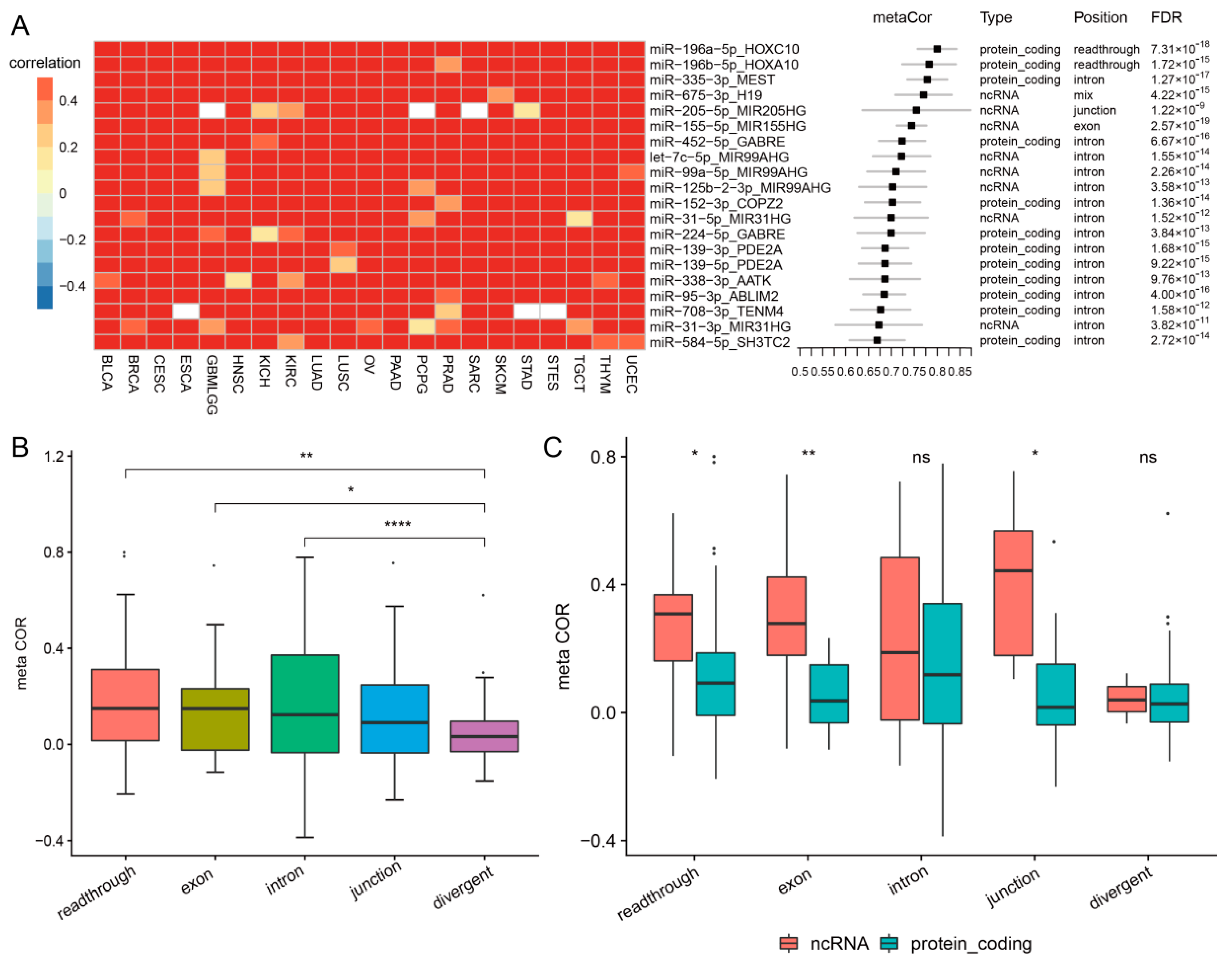
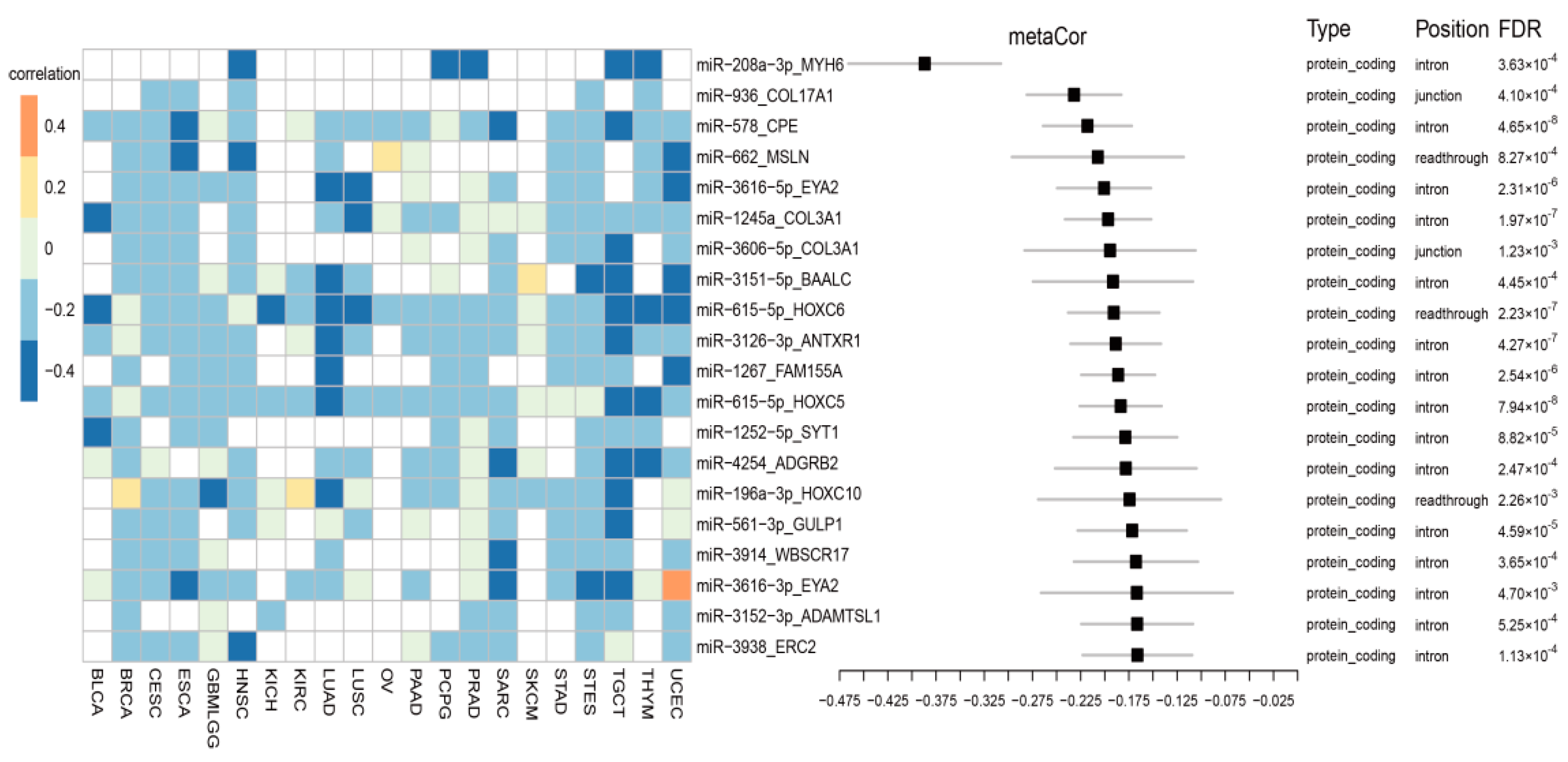
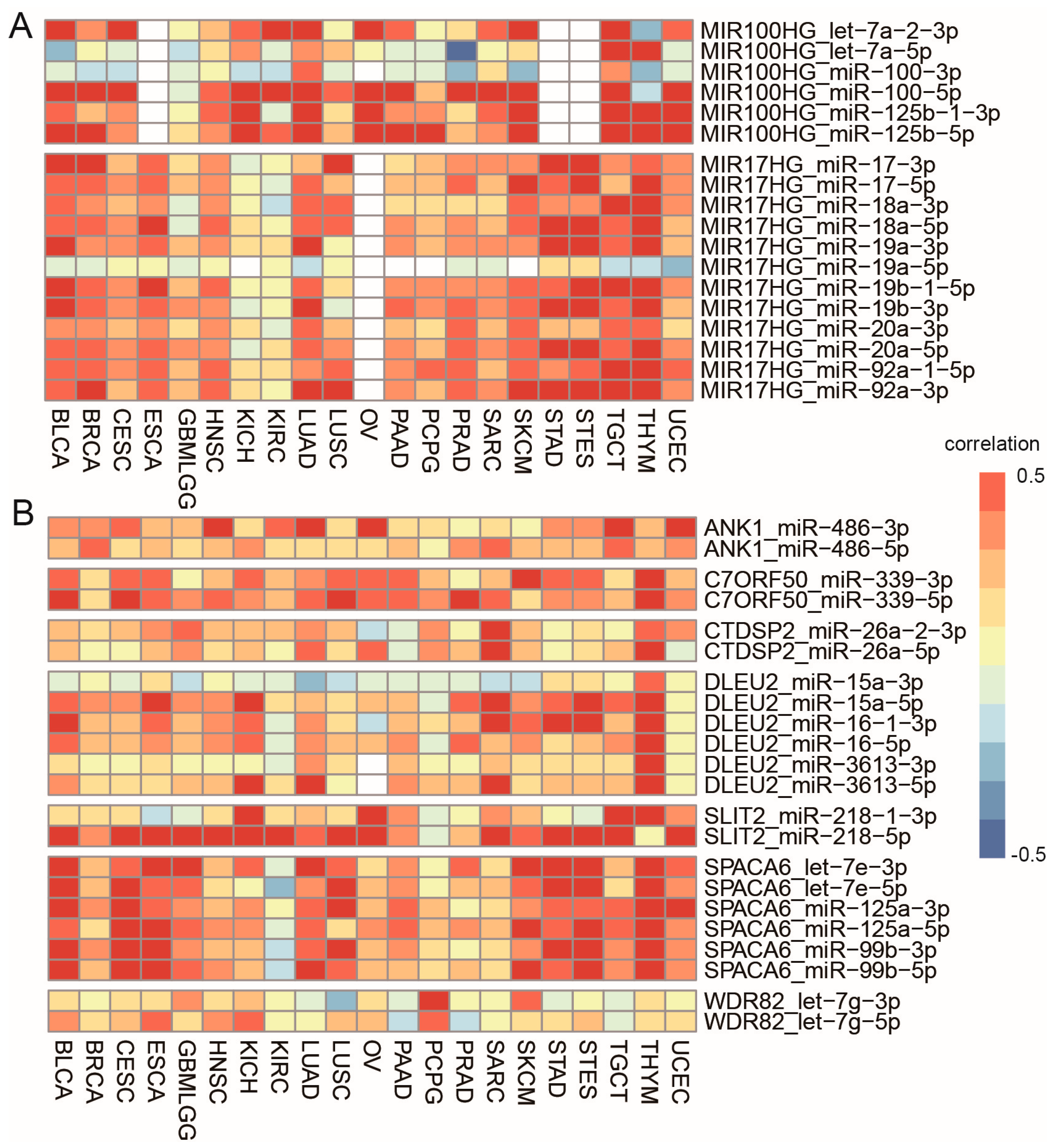
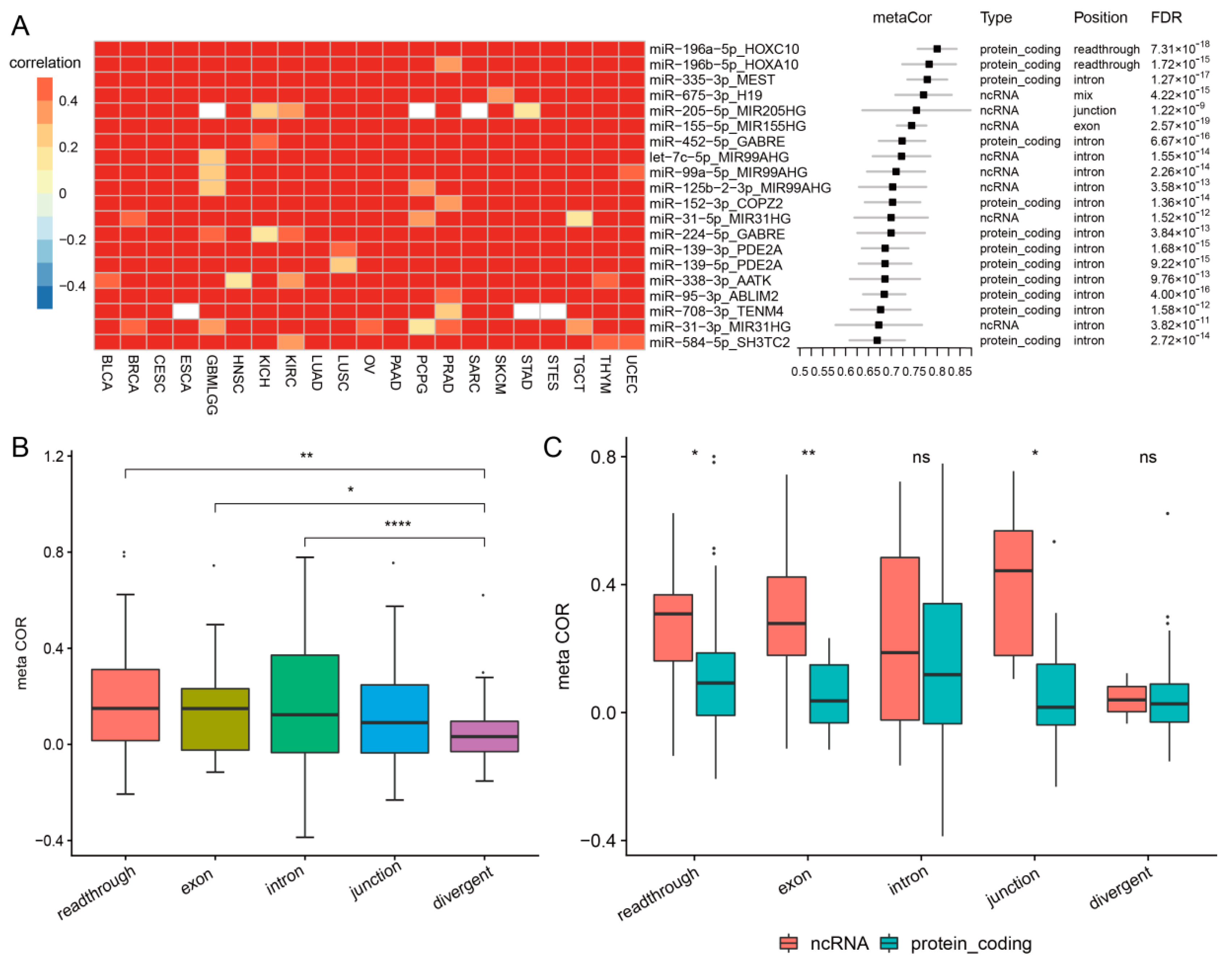
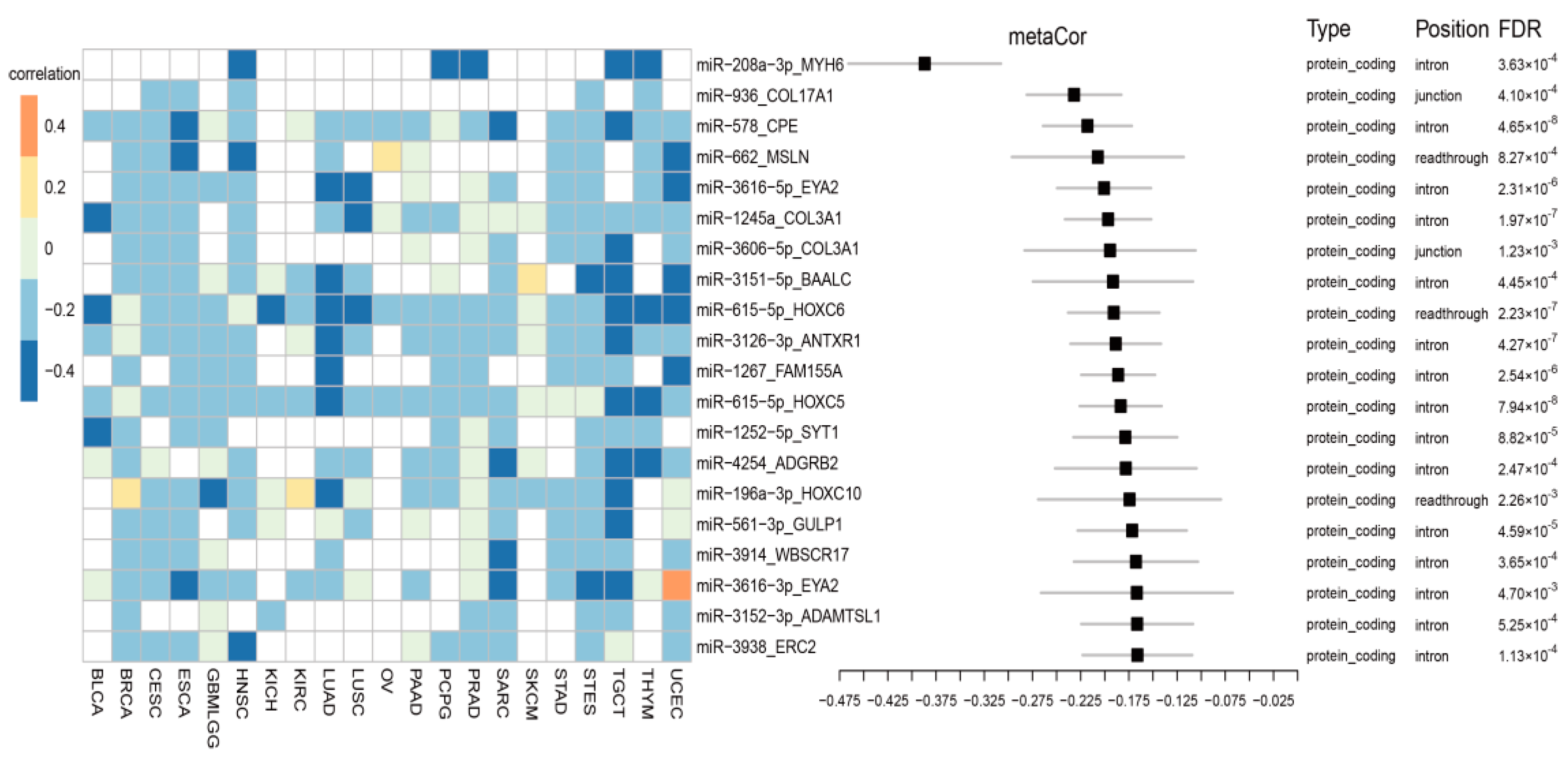
3.2. Cancer-Common Transcriptional Relationships between miRNAs and Host Genes
3.3. Cancer-Specific Co-Transcription between miRNAs and Host Genes
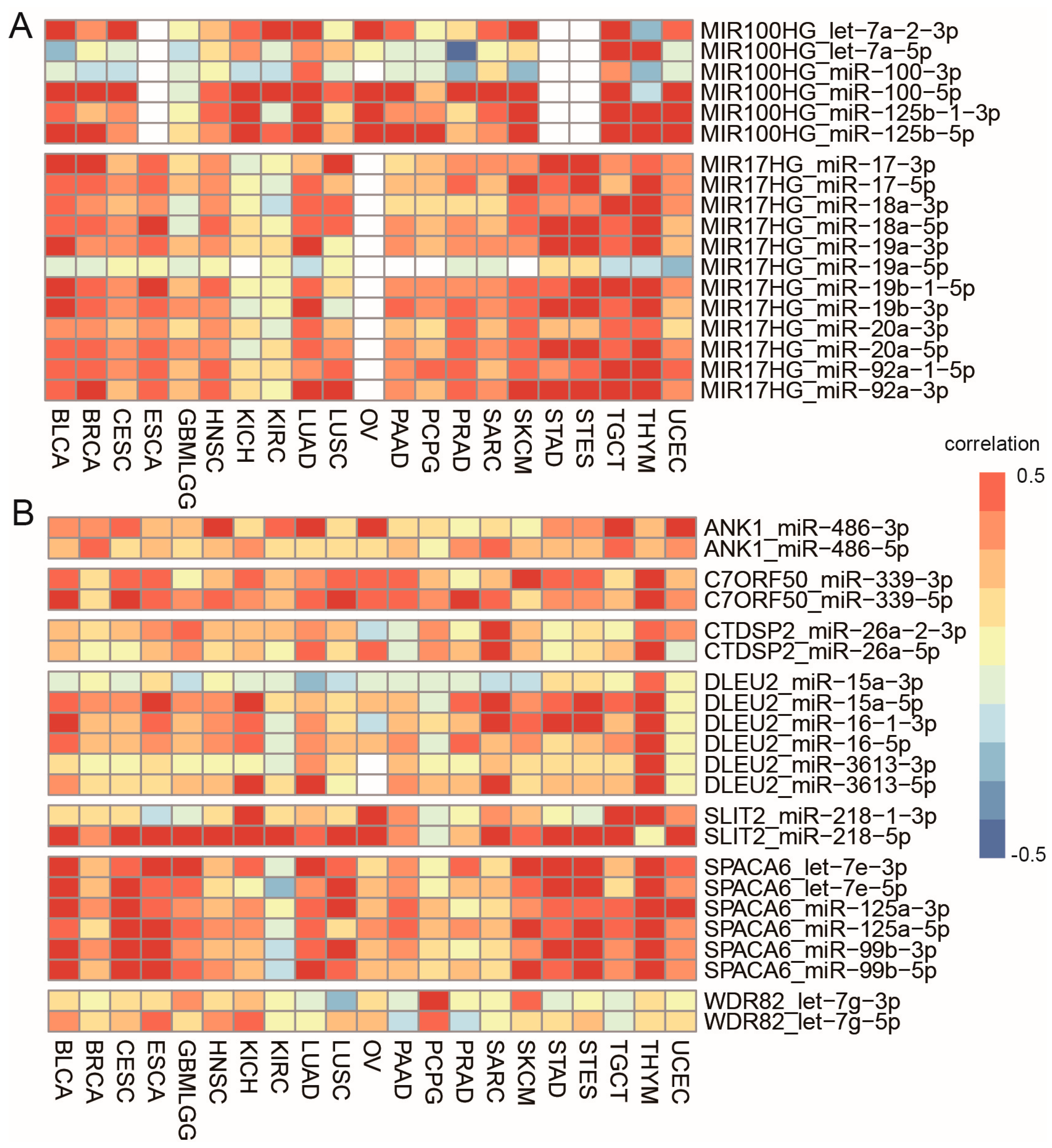
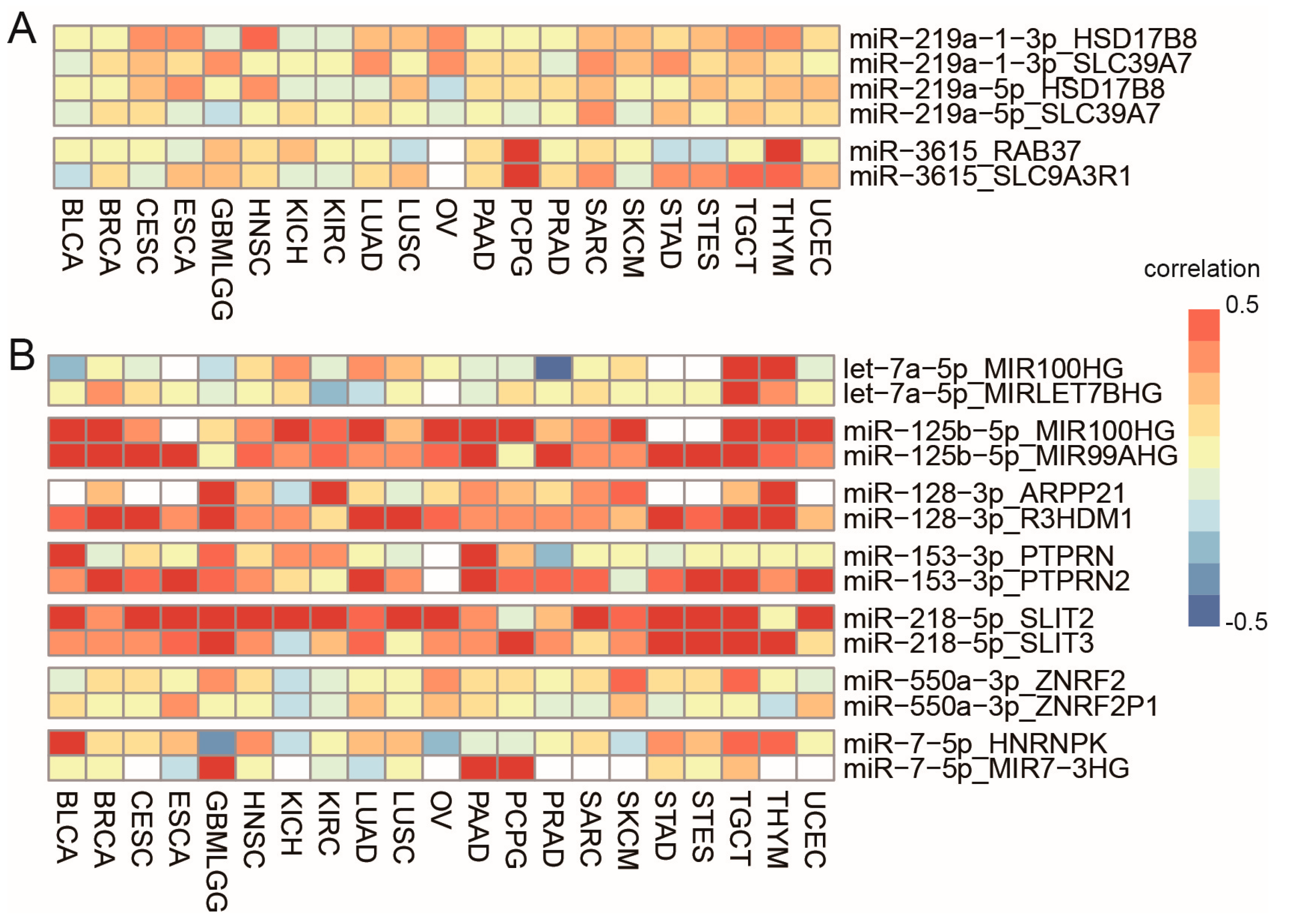
3.4. Cancer-Specific Selection of Sister miRNA Pairs
3.5. Cancer-Specific Selection of Host Genes and the Main Contributors
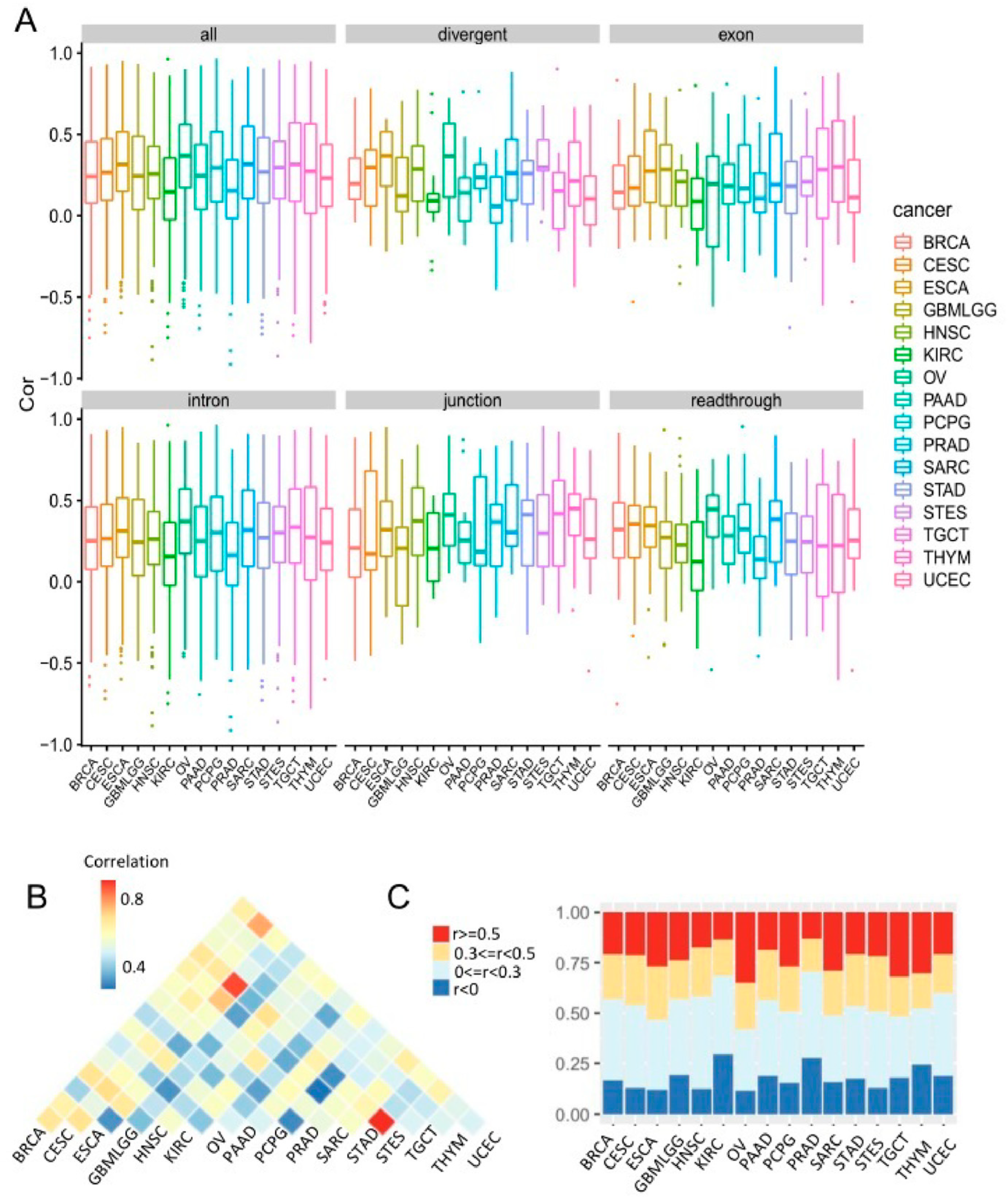
3.6. Comparison of Co-Transcription Patterns across Cancer Types
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, G.C.; Singh, J.; Barik, S. MicroRNAs: Processing, Maturation, Target Recognition and Regulatory Functions. Mol. Cell Pharmacol. 2011, 3, 83–92. [Google Scholar]
- Kloosterman, W.P.; Plasterk, R.H. The diverse functions of microRNAs in animal development and disease. Dev. Cell 2006, 11, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, M.; Ling, H.; Doki, Y.; Mori, M.; Calin, G.A. MicroRNA Processing and Human Cancer. J. Clin. Med. 2015, 4, 1651–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perron, M.P.; Provost, P. Protein interactions and complexes in human microRNA biogenesis and function. Front. Biosci. J. Virtual Libr. 2008, 13, 2537–2547. [Google Scholar] [CrossRef] [Green Version]
- Islam, A.; Mohammad, E.; Khan, M.A. Aberration of the modulatory functions of intronic microRNA hsa-miR-933 on its host gene ATF2 results in type II diabetes mellitus and neurodegenerative disease development. Hum. Genom. 2020, 14, 34. [Google Scholar] [CrossRef]
- Gao, X.; Qiao, Y.; Han, D.; Zhang, Y.; Ma, N. Enemy or partner: Relationship between intronic micrornas and their host genes. IUBMB Life 2012, 64, 835–840. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Li, Z.; Chen, P.; Huang, H.; Hurst, L.D.; Chen, J. Young intragenic miRNAs are less coexpressed with host genes than old ones: Implications of miRNA–host gene coevolution. Nucleic Acids Res. 2012, 40, 4002–4012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Shyr, Y.; Cai, J.; Liu, Q. Interplay between miRNAs and host genes and their role in cancer. Brief. Funct. Genom. 2019, 18, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Wu, X.; Qian, W.; Cai, H.; Sun, X.; Zhang, W.; Tan, S.; Wu, Z.; Qian, P.; Ding, K.; et al. CCAR1 5′ UTR as a natural miRancer of miR-1254 overrides tamoxifen resistance. Cell Res. 2016, 26, 655–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, N.; Wang, X.; Qiao, Y.; Li, F.; Hui, Y.; Zou, C.; Jin, J.; Lv, G.; Peng, Y.; Wang, L.; et al. Coexpression of an intronic microRNA and its host gene reveals a potential role for miR-483-5p as an IGF2 partner. Mol. Cell Endocrinol. 2011, 333, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Hinske, L.C.G.; Galante, P.A.F.; Kuo, W.P.; Ohno-Machado, L. A potential role for intragenic miRNAs on their hosts’ interactome. BMC Genom. 2010, 11, 533. [Google Scholar] [CrossRef] [Green Version]
- Barron, N.; Sanchez, N.; Kelly, P.; Clynes, M. MicroRNAs: Tiny targets for engineering CHO cell phenotypes? Biotechnol. Lett. 2011, 33, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Baskerville, S.; Bartel, D.P. Microarray profiling of microRNAs reveals frequent coexpression with neighboring miRNAs and host genes. RNA 2005, 11, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Dohi, O.; Yasui, K.; Gen, Y.; Takada, H.; Endo, M.; Tsuji, K.; Konishi, C.; Yamada, N.; Mitsuyoshi, H.; Yagi, N.; et al. Epigenetic silencing of miR-335 and its host gene MEST in hepatocellular carcinoma. Int. J. Oncol. 2013, 42, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Georgakilas, G.; Vlachos, I.S.; Paraskevopoulou, M.D.; Yang, P.; Zhang, Y.; Economides, A.N.; Hatzigeorgiou, A.G. microTSS: Accurate microRNA transcription start site identification reveals a significant number of divergent pri-miRNAs. Nat. Commun. 2014, 5, 5700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seila, A.C.; Core, L.J.; Lis, J.T.; Sharp, P.A. Divergent transcription: A new feature of active promoters. Cell Cycle 2009, 8, 2557–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef]
- Drusco, A.; Croce, C.M. MicroRNAs and Cancer: A Long Story for Short RNAs. Adv. Cancer Res. 2017, 135, 1–24. [Google Scholar]
- Markopoulos, G.S.; Roupakia, E.; Tokamani, M.; Chavdoula, E.; Hatziapostolou, M.; Polytarchou, C.; Marcu, K.B.; Papavassiliou, A.G.; Sandaltzopoulos, R.; Kolettas, E. A step-by-step microRNA guide to cancer development and metastasis. Cell. Oncol. 2017, 40, 303–339. [Google Scholar] [CrossRef]
- Dragomir, M.; Mafra, A.C.; Dias, S.M.G.; Vasilescu, C.; Calin, G.A. Using microRNA Networks to Understand Cancer. Int. J. Mol. Sci. 2018, 19, 1871. [Google Scholar] [CrossRef] [Green Version]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Goodall, G.J.; Wickramasinghe, V.O. RNA in cancer. Nat. Rev. Cancer 2021, 21, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Veneziano, D.; Acunzo, M.; Croce, C.M. Small non-coding RNA and cancer. Carcinogenesis 2017, 38, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shkurnikov, M.; Nikulin, S.; Nersisyan, S.; Poloznikov, A.; Zaidi, S.; Baranova, A.; Schumacher, U.; Wicklein, D.; Tonevitsky, A. LAMA4-Regulating miR-4274 and Its Host Gene SORCS2 Play a Role in IGFBP6-Dependent Effects on Phenotype of Basal-Like Breast Cancer. Front. Mol. Biosci. 2019, 6, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Ji, F.; Kumar, M.R.; Zheng, X.; Xiao, Y.; Liu, N.; Shi, J.; Wong, L.; Forgues, M.; Qin, L.X.; et al. Transcriptome integration analysis in hepatocellular carcinoma reveals discordant intronic miRNA-host gene pairs in expression. Int. J. Biol. Sci. 2017, 13, 1438–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, L.; Sun, T. Cohesive Regulation of Neural Progenitor Development by microRNA miR-26, Its Host Gene Ctdsp and Target Gene Emx2 in the Mouse Embryonic Cerebral Cortex. Front. Mol. Neurosci. 2018, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Cao, G.; Huang, B.; Liu, Z.; Zhang, J.; Xu, H.; Xia, W.; Li, J.; Li, S.; Chen, L.; Ding, H.; et al. Intronic miR-301 feedback regulates its host gene, ska2, in A549 cells by targeting MEOX2 to affect ERK/CREB pathways. Biochem. Biophys. Res. Commun. 2010, 396, 978–982. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Osokine, I.; Hsu, R.; Loeb, G.B.; McManus, M.T. Unintentional miRNA Ablation Is a Risk Factor in Gene Knockout Studies: A Short Report. PLoS Genet. 2008, 4, e34. [Google Scholar] [CrossRef]
- Gennarino, V.A.; Sardiello, M.; Avellino, R.; Meola, N.; Maselli, V.; Anand, S.; Cutillo, L.; Ballabio, A.; Banfi, S. MicroRNA target prediction by expression analysis of host genes. Genome Res. 2008, 19, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Balduzzi, S.; Rucker, G.; Schwarzer, G. How to perform a meta-analysis with R: A practical tutorial. Evid. Based Ment. Health 2019, 22, 153–160. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, A.T.; Katsyv, I.; Song, W.-M.; Wang, M.; Zhang, B. DGCA: A comprehensive R package for Differential Gene Correlation Analysis. BMC Syst. Biol. 2016, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Franca, G.S.; Vibranovski, M.D.; Galante, P.A. Host gene constraints and genomic context impact the expression and evolution of human microRNAs. Nat. Commun. 2016, 7, 11438. [Google Scholar] [CrossRef]
- França, G.S.; Hinske, L.C.; Galante, P.A.F.; Vibranovski, M. Unveiling the Impact of the Genomic Architecture on the Evolution of Vertebrate microRNAs. Front. Genet. 2017, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasiolo, M.; Sales, G.; Lionetti, M.; Agnelli, L.; Todoerti, K.; Bisognin, A.; Coppe, A.; Romualdi, C.; Neri, A.; Bortoluzzi, S. Impact of Host Genes and Strand Selection on miRNA and miRNA Expression. PLoS ONE 2011, 6, e23854. [Google Scholar] [CrossRef]
- Ribas, J.; Ni, X.; Castanares, M.; Liu, M.M.; Esopi, D.; Yegnasubramanian, S.; Rodriguez, R.; Mendell, J.T.; Lupold, S.E. A novel source for miR-21 expression through the alternative polyadenylation of VMP1 gene transcripts. Nucleic Acids Res. 2012, 40, 6821–6833. [Google Scholar] [CrossRef] [Green Version]
- Agranat-Tamir, L.; Shomron, N.; Sperling, J.; Sperling, R. Interplay between pre-mRNA splicing and microRNA biogenesis within the supraspliceosome. Nucleic Acids Res. 2014, 42, 4640–4651. [Google Scholar] [CrossRef]
- Meenakshisundaram, G.; Common, J.; Gopal, F.E.; Srikanta, S.; Lakshman, K.; Lunny, D.; Lim, T.C.; Tanavde, V.; Lane, E.B.; Sampath, P. ‘See-saw’ expression of microRNA-198 and FSTL1 from a single transcript in wound healing. Nature 2013, 495, 103–106. [Google Scholar] [CrossRef]
- Batista, L.; Bourachot, B.; Mateescu, B.; Reyal, F.; Mechta-Grigoriou, F. Regulation of miR-200c/141 expression by intergenic DNA-looping and transcriptional read-through. Nat. Commun. 2016, 7, 8959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sestito, R.; Cianfrocca, R.; Tocci, P.; Rosano, L.; Sacconi, A.; Blandino, G.; Bagnato, A. Targeting endothelin 1 receptor-miR-200b/c-ZEB1 circuitry blunts metastatic progression in ovarian cancer. Commun. Biol. 2020, 3, 677. [Google Scholar] [CrossRef]
- Sulaiman, S.A.; Ab Mutalib, N.S.; Jamal, R. miR-200c Regulation of Metastases in Ovarian Cancer: Potential Role in Epithelial and Mesenchymal Transition. Front. Pharmacol. 2016, 7, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modur, V.; Singh, N.; Mohanty, V.; Chung, E.; Muhammad, B.; Choi, K.; Chen, X.; Chetal, K.; Ratner, N.; Salomonis, N.; et al. Defective transcription elongation in a subset of cancers confers immunotherapy resistance. Nat. Commun. 2018, 9, 4410. [Google Scholar] [CrossRef] [Green Version]
- Duan, D.R.; Pause, A.; Burgess, W.H.; Aso, T.; Chen, D.Y.; Garrett, K.P.; Conaway, R.C.; Linehan, W.M.; Klausner, R.D. Inhibition of transcription elongation by the VHL tumor suppressor protein. Science 1995, 269, 1402–1406. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Li, K.; Asaduzzaman, M.; Cuella, R.; Shi, H.; Raguz, S.; Coombes, R.C.; Zhou, Y.; Yagüe, E. miR-106b~25 cluster regulates multidrug resistance in an ABC transporter-independent manner via downregulation of EP300. Oncol. Rep. 2015, 35, 1170–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pidikova, P.; Herichova, I. miRNA Clusters with Up-Regulated Expression in Colorectal Cancer. Cancers 2021, 13, 2979. [Google Scholar] [CrossRef]
- Li, N.; Miao, Y.; Shan, Y.; Liu, B.; Li, Y.; Zhao, L.; Jia, L. MiR-106b and miR-93 regulate cell progression by suppression of PTEN via PI3K/Akt pathway in breast cancer. Cell Death Dis. 2017, 8, e2796. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Ho, J.-N.; Jin, H.; Lee, S.C.; Hong, S.-K.; Lee, J.W.; Lee, E.-S.; Byun, S.-S. Upregulated expression of BCL2, MCM7, and CCNE1 indicate cisplatin-resistance in the set of two human bladder cancer cell lines: T24 cisplatin sensitive and T24R2 cisplatin resistant bladder cancer cell lines. Investig. Clin. Urol. 2016, 57, 63–72. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Jiang, Y.Y.; Hao, J.J.; Lu, S.S.; Zhang, T.T.; Shang, L.; Cao, J.; Song, X.; Wang, B.S.; Cai, Y.; et al. Prognostic significance of MCM7 expression in the bronchial brushings of patients with non-small cell lung cancer (NSCLC). Lung Cancer 2012, 77, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lan, P.; Hou, Z.; Guan, Y.; Zhang, J.; Xu, W.; Tian, Z.; Zhang, C. Histone deacetylase inhibitor SAHA epigenetically regulates miR-17-92 cluster and MCM7 to upregulate MICA expression in hepatoma. Br. J. Cancer 2014, 112, 112–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalingam, P.; Palanichamy, J.K.; Singh, A.; Das, P.; Bhagat, M.; Kassab, M.A.; Sinha, S.; Chattopadhyay, P. Biogenesis of intronic miRNAs located in clusters by independent transcription and alternative splicing. RNA 2013, 20, 76–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schanen, B.C.; Li, X. Transcriptional regulation of mammalian miRNA genes. Genomics 2011, 97, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Shin, C.H.; Lee, H.; Choi, K.H.; Nam, D.-H.; Ohn, T.; Kim, H.H. MicroRNA-1908-5p contributes to the oncogenic function of the splicing factor SRSF3. Oncotarget 2016, 8, 8342–8355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bublik, D.R.; Bursac, S.; Sheffer, M.; Orsolic, I.; Shalit, T.; Tarcic, O.; Kotler, E.; Mouhadeb, O.; Hoffman, Y.; Fuchs, G.; et al. Regulatory module involving FGF13, miR-504, and p53 regulates ribosomal biogenesis and supports cancer cell survival. Proc. Natl. Acad Sci. USA 2017, 114, E496–E505. [Google Scholar] [CrossRef] [Green Version]
- Cui, R.; Guan, Y.; Sun, C.; Chen, L.; Bao, Y.; Li, G.; Qiu, B.; Meng, X.; Pang, C.; Wang, Y. A tumor-suppressive microRNA, miR-504, inhibits cell proliferation and promotes apoptosis by targeting FOXP1 in human glioma. Cancer Lett. 2016, 374, 1–11. [Google Scholar] [CrossRef]
- Melamed, Z.; Levy, A.; Ashwal-Fluss, R.; Lev-Maor, G.; Mekahel, K.; Atias, N.; Gilad, S.; Sharan, R.; Levy, C.; Kadener, S.; et al. Alternative splicing regulates biogenesis of miRNAs located across exon-intron junctions. Mol. Cell 2013, 50, 869–881. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wang, Y.; Yu, T.; Nie, E.; Hu, Q.; Wu, W.; Zhi, T.; Jiang, K.; Wang, X.; Lu, X.; et al. Blocking MIR155HG/miR-155 axis inhibits mesenchymal transition in glioma. Neuro Oncol. 2017, 19, 1195–1205. [Google Scholar] [CrossRef]
- Wu, X.; Sharp, P.A. Divergent transcription: A driving force for new gene origination? Cell 2013, 155, 990–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Peng, R.; Zeng, M.; Zhang, Z.; Liu, S.; Jiang, D.; Lu, Y.; Zou, F. An autoregulatory feedback loop of miR-21/VMP1 is responsible for the abnormal expression of miR-21 in colorectal cancer cells. Cell Death Dis. 2020, 11, 1067. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ma, R.; Yi, B.; Riker, A.I.; Xi, Y. MicroRNAs are involved in the development and progression of gastric cancer. Acta Pharmacol. Sin. 2020, 42, 1018–1026. [Google Scholar] [CrossRef]
- Palmero, E.I.; de Campos, S.G.; Campos, M.; de Souza, N.C.; Guerreiro, I.D.; Carvalho, A.L.; Marques, M.M. Mechanisms and role of microRNA deregulation in cancer onset and progression. Genet. Mol. Biol. 2011, 34, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Zarrilli, G.; Galuppini, F.; Angerilli, V.; Munari, G.; Sabbadin, M.; Lazzarin, V.; Nicolè, L.; Biancotti, R.; Fassan, M. miRNAs Involved in Esophageal Carcinogenesis and miRNA-Related Therapeutic Perspectives in Esophageal Carcinoma. Int. J. Mol. Sci. 2021, 22, 3640. [Google Scholar] [CrossRef] [PubMed]
- Gambari, R.; Brognara, E.; Spandidos, D.A.; Fabbri, E. Targeting oncomiRNAs and mimicking tumor suppressor miRNAs: Nuew trends in the development of miRNA therapeutic strategies in oncology (Review). Int. J. Oncol. 2016, 49, 5–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Cui, L.; Ahmed, S.; Zainab, F.; Wu, Q.; Wang, X.; Yuan, Z. An overview of epigenetic agents and natural nutrition products targeting DNA methyltransferase, histone deacetylases and microRNAs. Food Chem. Toxicol. 2018, 123, 574–594. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer | Description | Paired Samples |
|---|---|---|
| BLCA | Bladder urothelial carcinoma | 56 |
| BRCA | Breast invasive carcinoma | 458 |
| CESC | Cervical and endocervical cancers | 304 |
| ESCA | Esophageal carcinoma | 183 |
| GBMLGG | Glioma (Brain Lower Grade Glioma) | 512 |
| HNSC | Head and Neck squamous cell carcinoma | 228 |
| KICH | Kidney Chromophobe | 66 |
| KIRC | Kidney renal clear cell carcinoma | 204 |
| LUAD | Lung adenocarcinoma | 79 |
| LUSC | Lung squamous cell carcinoma | 69 |
| OV | Ovarian serous cystadenocarcinoma | 288 |
| PAAD | Pancreatic adenocarcinoma | 178 |
| PCPG | Pheochromocytoma and Paraganglioma | 179 |
| PRAD | Prostate adenocarcinoma | 493 |
| SARC | Sarcoma | 257 |
| SKCM | Skin Cutaneous Melanoma | 97 |
| STAD | Stomach adenocarcinoma | 232 |
| STES | Stomach and Esophageal carcinoma | 415 |
| TGCT | Testicular Germ Cell Tumors | 150 |
| THYM | Thymoma | 120 |
| UCEC | Uterine Corpus Endometrial Carcinoma | 139 |
| Type | Number | Percent (%) |
|---|---|---|
| Intronic | 918 | 48.9 |
| Exonic | 74 | 3.9 |
| Junction | 45 | 2.4 |
| Readthrough | 59 | 3.1 |
| Divergent | 50 | 2.6 |
| Mixed | 90 | 4.8 |
| Antisense | 198 | 10.5 |
| Intergenic | 447 | 23.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Shyr, Y.; Liu, Q. Pan-Cancer Analysis Reveals Common and Specific Relationships between Intragenic miRNAs and Their Host Genes. Biomedicines 2021, 9, 1263. https://doi.org/10.3390/biomedicines9091263
Liu B, Shyr Y, Liu Q. Pan-Cancer Analysis Reveals Common and Specific Relationships between Intragenic miRNAs and Their Host Genes. Biomedicines. 2021; 9(9):1263. https://doi.org/10.3390/biomedicines9091263
Chicago/Turabian StyleLiu, Baohong, Yu Shyr, and Qi Liu. 2021. "Pan-Cancer Analysis Reveals Common and Specific Relationships between Intragenic miRNAs and Their Host Genes" Biomedicines 9, no. 9: 1263. https://doi.org/10.3390/biomedicines9091263
APA StyleLiu, B., Shyr, Y., & Liu, Q. (2021). Pan-Cancer Analysis Reveals Common and Specific Relationships between Intragenic miRNAs and Their Host Genes. Biomedicines, 9(9), 1263. https://doi.org/10.3390/biomedicines9091263