
Multiple-Organ Complement Deposition on Vascular Endothelium in COVID-19 Patients

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Group
2.2. Tissue Sample Collection
2.3. Immunofluorescence Analysis
2.4. Western Blot Analysis
2.5. ELISA for C5b-9 Analysis
3. Results
3.1. Clinical Data
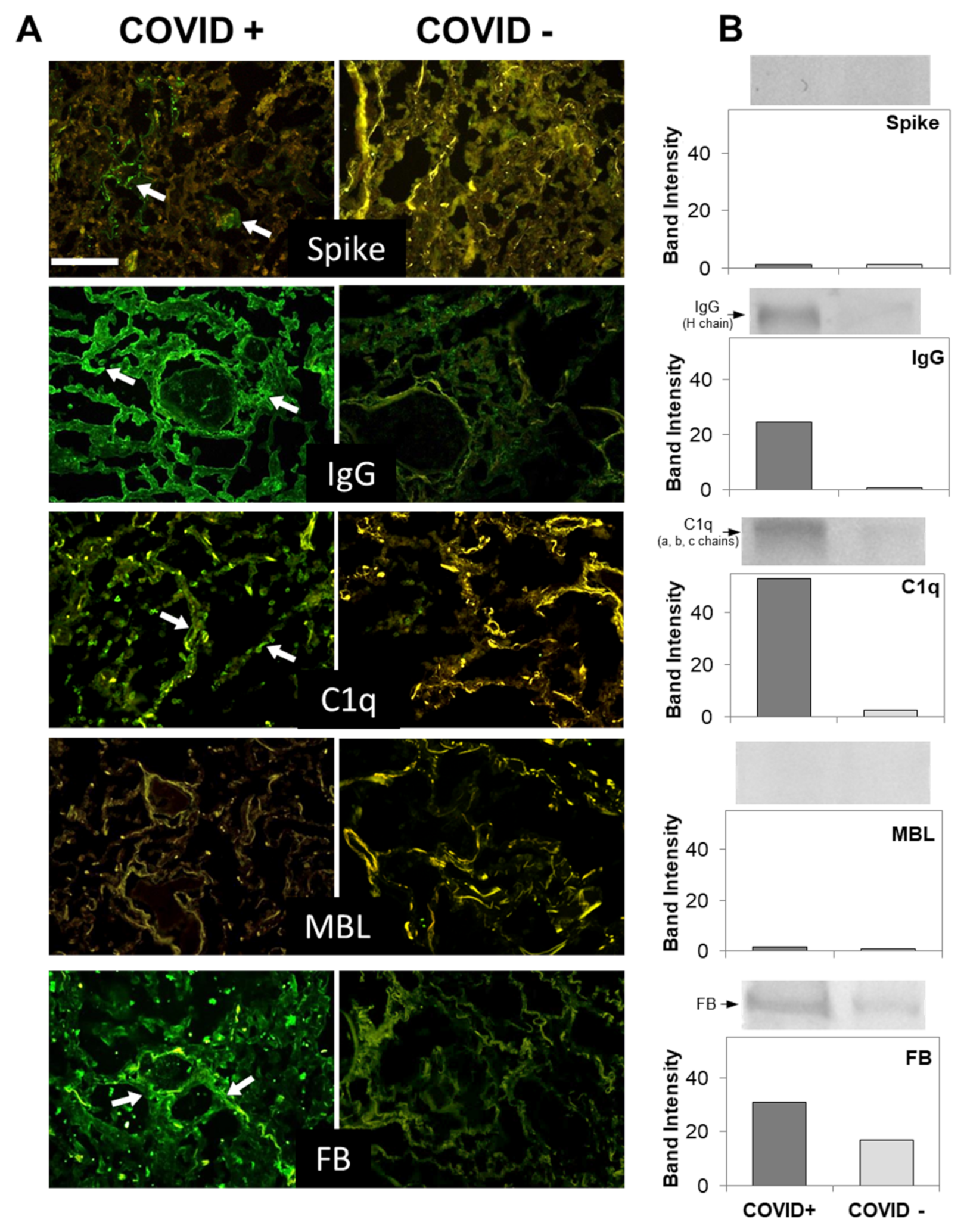
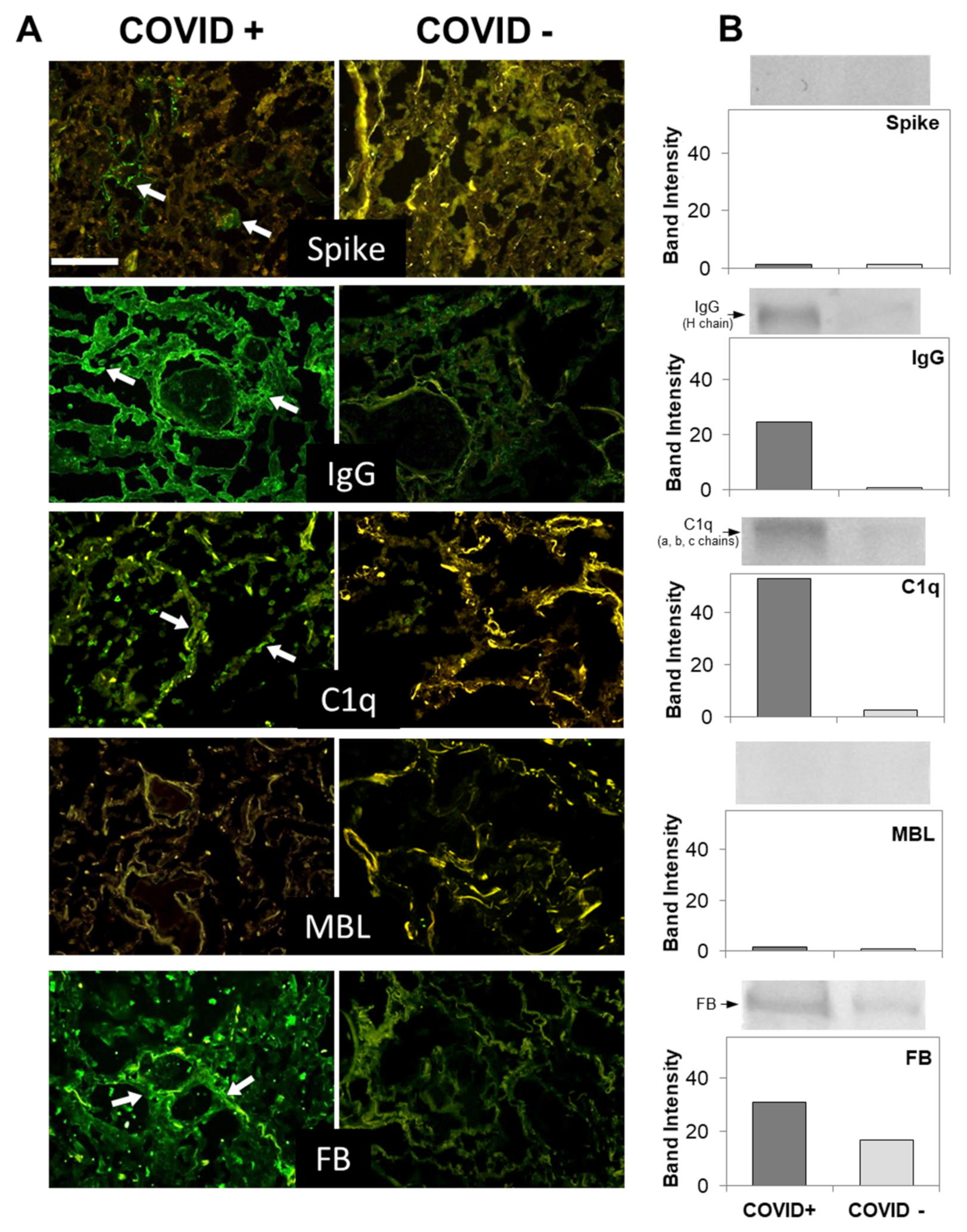
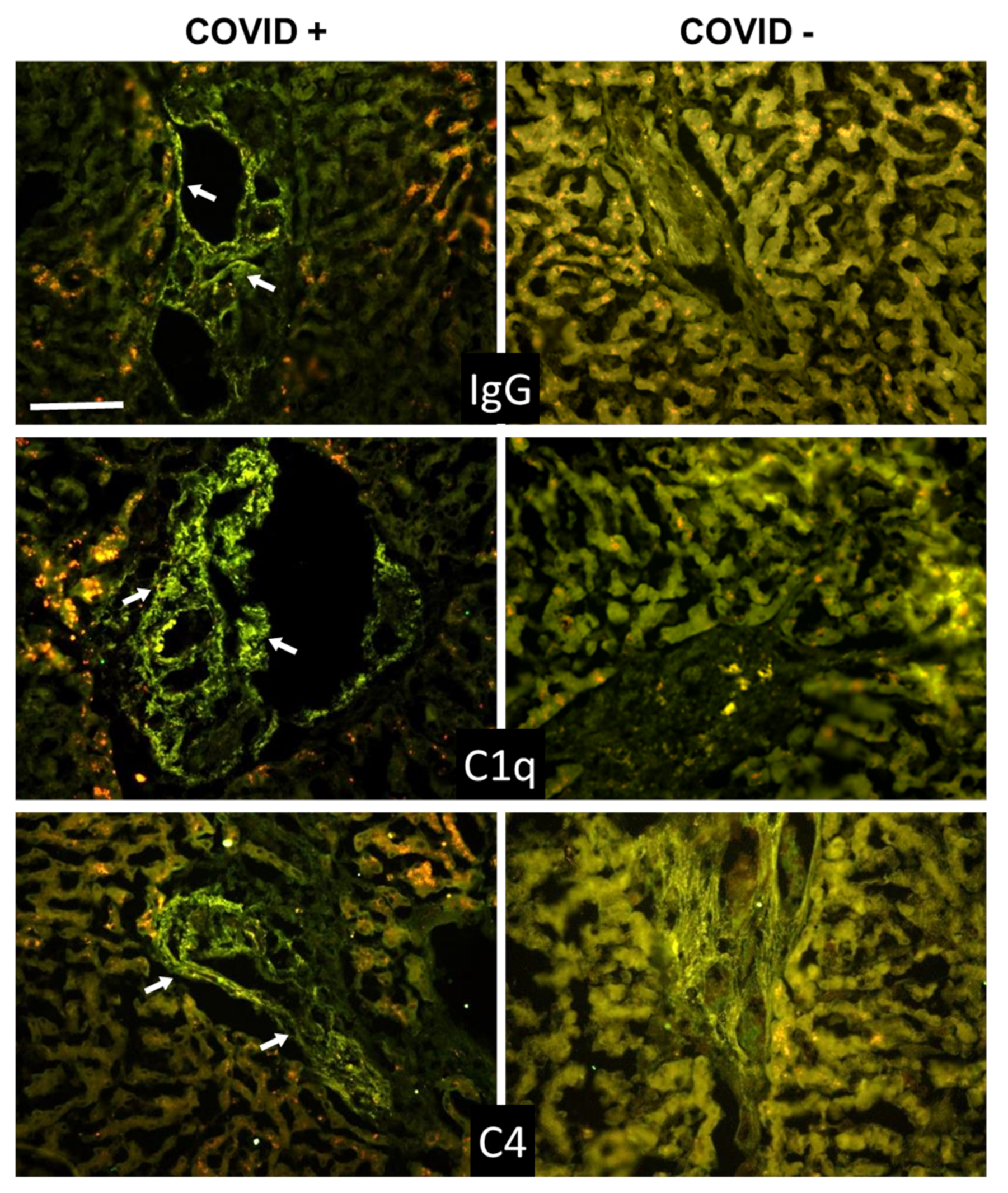
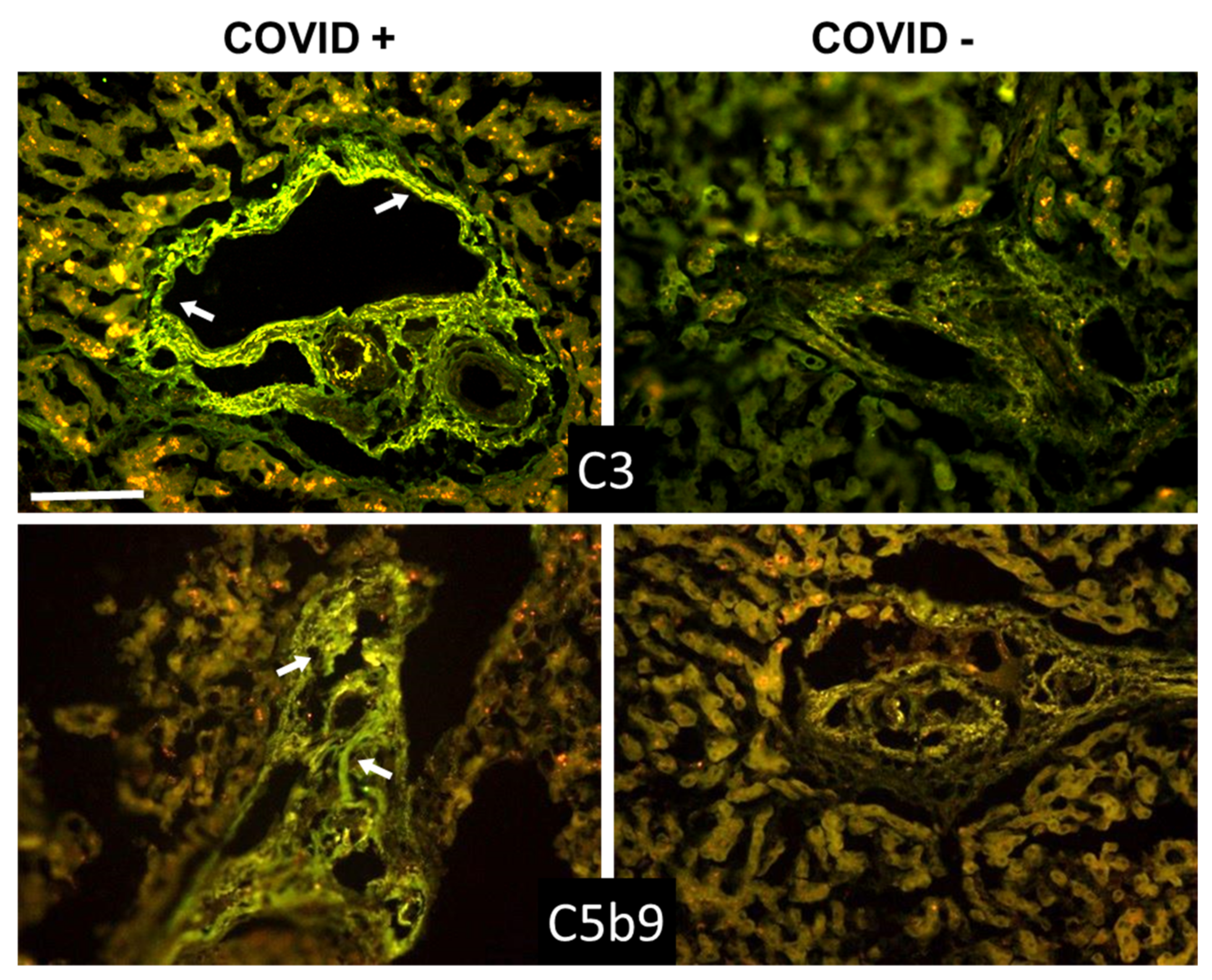
3.2. C Deposition in the Lungs
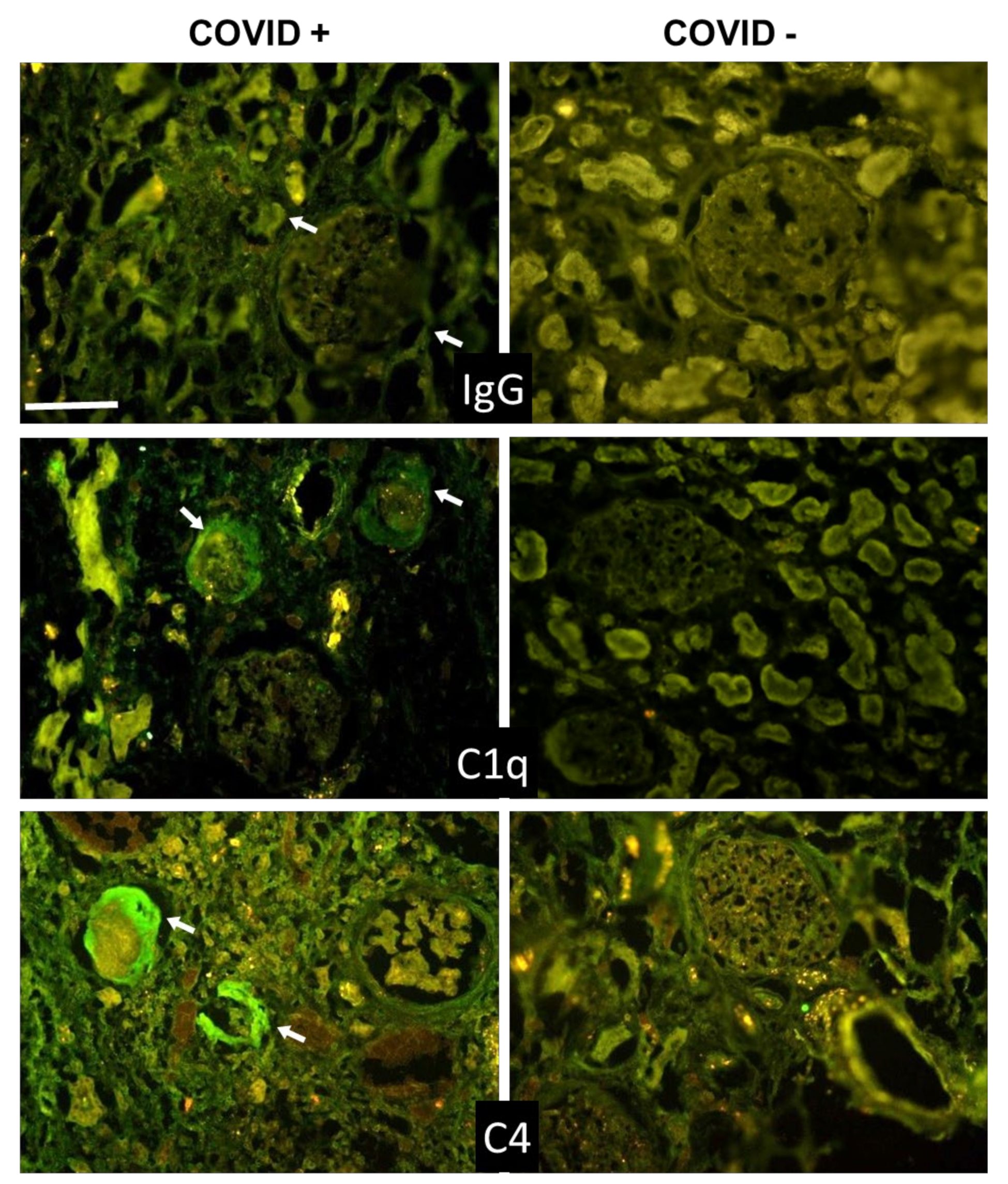
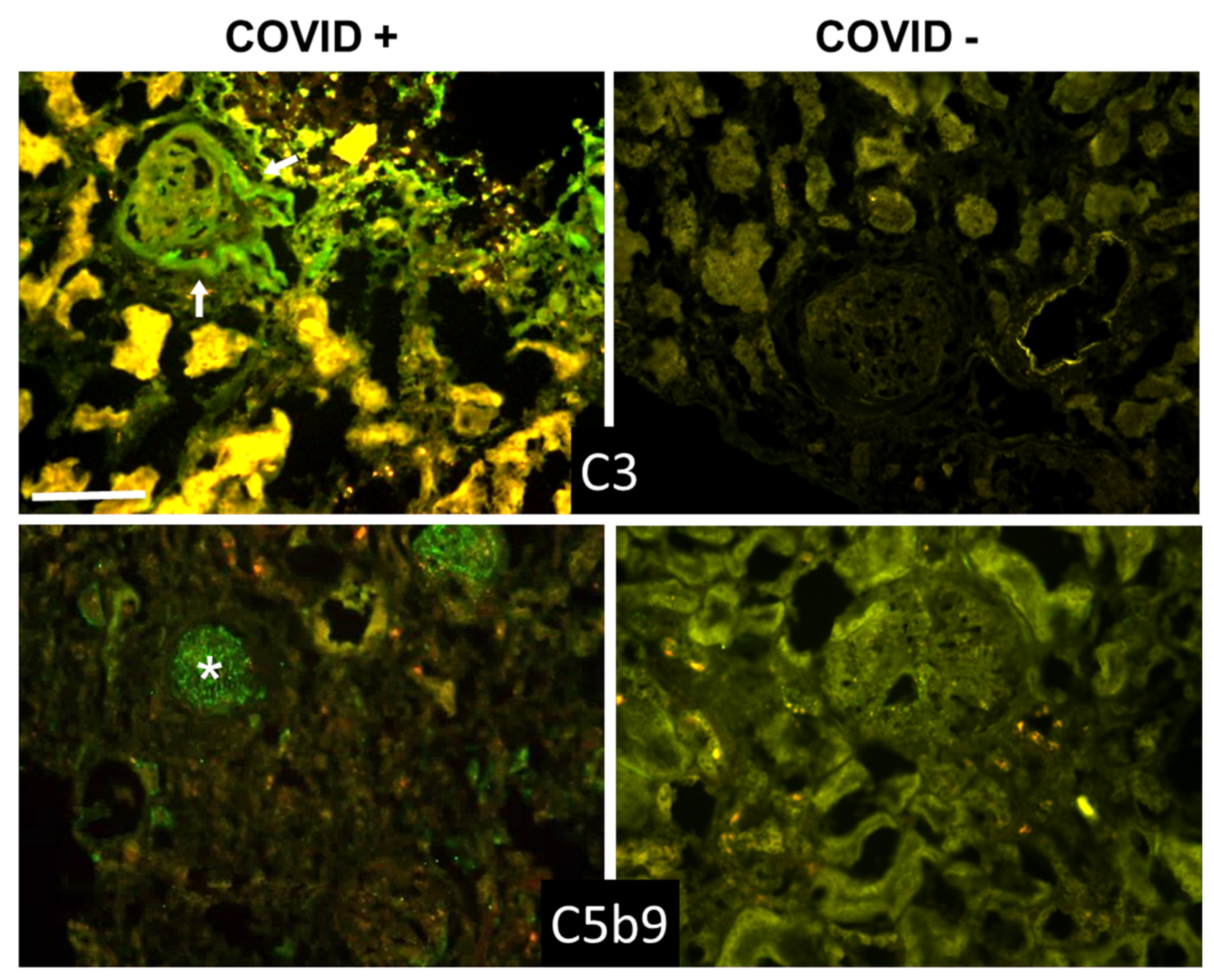
3.3. C Deposition in Kidney and Liver
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Remuzzi, A.; Remuzzi, G. COVID-19 and Italy: What next? Lancet 2020, 395, 1225–1228. [Google Scholar] [CrossRef]
- Fu, L.; Wang, B.; Yuan, T.; Chen, X.; Ao, Y.; Fitzpatrick, T.; Li, P.; Zhou, Y.; Lin, Y.F.; Duan, Q.; et al. Clinical characteristics of coronavirus disease 2019 (COVID-19) in China: A systematic review and meta-analysis. J. Infect. 2020, 80, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, I.M.; Padera, R.F.; Solomon, I.H.; Kanjilal, S.; Hammer, M.M.; Hornick, J.L.; Sholl, L.M. In situ detection of SARS-CoV-2 in lungs and airways of patients with COVID-19. Mod. Pathol. 2020, 33, 2104–2114. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Xie, M.; Zhang, W. Clinical characteristics and intrauterine vertical transmission potential of coronavirus disease 2019 infection in 9 pregnant women: A retrospective review of medical records. Am. J. Obstet. Gynecol. 2020, 223, 955–956. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Hikmet, F.; Mear, L.; Edvinsson, A.; Micke, P.; Uhlen, M.; Lindskog, C. The protein expression profile of ACE2 in human tissues. Mol. Syst. Biol. 2020, 16, e9610. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, Y.; Novelli, L.; Rojas, M.; De Santis, M.; Acosta-Ampudia, Y.; Monsalve, D.M.; Ramirez-Santana, C.; Costanzo, A.; Ridgway, W.M.; Ansari, A.A.; et al. Autoinflammatory and autoimmune conditions at the crossroad of COVID-19. J. Autoimmun. 2020, 114, 102506. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, J.; Wang, D.; Guan, W.; Qin, J.; Xu, X.; Fang, J.; Fu, B.; Zheng, X.; Wang, D.; et al. Profiling of the immune repertoire in COVID-19 patients with mild, severe, convalescent, or retesting-positive status. J. Autoimmun. 2021, 118, 102596. [Google Scholar] [CrossRef]
- Vardhana, S.A.; Wolchok, J.D. The many faces of the anti-COVID immune response. J. Exp. Med. 2020, 217, e20200678. [Google Scholar] [CrossRef]
- Luo, W.; Zhang, J.W.; Zhang, W.; Lin, Y.L.; Wang, Q. Circulating levels of IL-2, IL-4, TNF-alpha, IFN-gamma, and C-reactive protein are not associated with severity of COVID-19 symptoms. J. Med. Virol. 2021, 93, 89–91. [Google Scholar] [CrossRef]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Cugno, M.; Meroni, P.L.; Gualtierotti, R.; Griffini, S.; Grovetti, E.; Torri, A.; Lonati, P.; Grossi, C.; Borghi, M.O.; Novembrino, C.; et al. Complement activation and endothelial perturbation parallel COVID-19 severity and activity. J. Autoimmun. 2021, 116, 102560. [Google Scholar] [CrossRef] [PubMed]
- Cugno, M.; Meroni, P.L.; Gualtierotti, R.; Griffini, S.; Grovetti, E.; Torri, A.; Panigada, M.; Aliberti, S.; Blasi, F.; Tedesco, F.; et al. Complement activation in patients with COVID-19: A novel therapeutic target. J. Allergy Clin. Immunol. 2020, 146, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Hu, M.; Zhang, X.; Li, H.; Zhu, L.; Liu, H.; Dong, Q.; Zhang, Z.; Wang, Z.; Hu, Y.; et al. Highly pathogenic coronavirus N protein aggravates lung injury by MASP-2-mediated complement over-activation. medRxiv 2020, 25. [Google Scholar] [CrossRef]
- Valenti, L.; Griffini, S.; Lamorte, G.; Grovetti, E.; Uceda Renteria, S.C.; Malvestiti, F.; Scudeller, L.; Bandera, A.; Peyvandi, F.; Prati, D.; et al. Chromosome 3 cluster rs11385942 variant links complement activation with severe COVID-19. J. Autoimmun. 2021, 117, 102595. [Google Scholar] [CrossRef]
- Carvelli, J.; Demaria, O.; Vely, F.; Batista, L.; Chouaki Benmansour, N.; Fares, J.; Carpentier, S.; Thibult, M.L.; Morel, A.; Remark, R.; et al. Association of COVID-19 inflammation with activation of the C5a-C5aR1 axis. Nature 2020, 588, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Holter, J.C.; Pischke, S.E.; de Boer, E.; Lind, A.; Jenum, S.; Holten, A.R.; Tonby, K.; Barratt-Due, A.; Sokolova, M.; Schjalm, C.; et al. Systemic complement activation is associated with respiratory failure in COVID-19 hospitalized patients. Proc. Natl. Acad. Sci. USA 2020, 117, 25018–25025. [Google Scholar] [CrossRef]
- Magro, C.; Mulvey, J.J.; Berlin, D.; Nuovo, G.; Salvatore, S.; Harp, J.; Baxter-Stoltzfus, A.; Laurence, J. Complement associated microvascular injury and thrombosis in the pathogenesis of severe COVID-19 infection: A report of five cases. Transl. Res. 2020, 220, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bussani, R.; Schneider, E.; Zentilin, L.; Collesi, C.; Ali, H.; Braga, L.; Volpe, M.C.; Colliva, A.; Zanconati, F.; Berlot, G.; et al. Persistence of viral RNA, pneumocyte syncytia and thrombosis are hallmarks of advanced COVID-19 pathology. EBioMedicine 2020, 61, 103104. [Google Scholar] [CrossRef] [PubMed]
- Colombo, F.; Durigutto, P.; De Maso, L.; Biffi, S.; Belmonte, B.; Tripodo, C.; Oliva, R.; Bardini, P.; Marini, G.M.; Terreno, E.; et al. Targeting CD34+ cells of the inflamed synovial endothelium by guided nanoparticles for the treatment of rheumatoid arthritis. J. Autoimmun. 2019, 103, 102288. [Google Scholar] [CrossRef]
- Capolla, S.; Mezzaroba, N.; Zorzet, S.; Tripodo, C.; Mendoza-Maldonado, R.; Granzotto, M.; Vita, F.; Spretz, R.; Larsen, G.; Noriega, S.; et al. A new approach for the treatment of CLL using chlorambucil/hydroxychloroquine-loaded anti-CD20 nanoparticles. Nano Res. 2016, 9, 11. [Google Scholar] [CrossRef]
- Ruffatti, A.; Tarzia, V.; Fedrigo, M.; Calligaro, A.; Favaro, M.; Macor, P.; Tison, T.; Cucchini, U.; Cosmi, E.; Tedesco, F.; et al. Evidence of complement activation in the thrombotic small vessels of a patient with catastrophic antiphospholipid syndrome treated with eculizumab. Autoimmun. Rev. 2019, 18, 561–563. [Google Scholar] [CrossRef]
- Chen, T.; Wu, D.; Chen, H.; Yan, W.; Yang, D.; Chen, G.; Ma, K.; Xu, D.; Yu, H.; Wang, H.; et al. Clinical characteristics of 113 deceased patients with coronavirus disease 2019: Retrospective study. BMJ 2020, 368, m1091. [Google Scholar] [CrossRef] [Green Version]
- Carsana, L.; Sonzogni, A.; Nasr, A.; Rossi, R.S.; Pellegrinelli, A.; Zerbi, P.; Rech, R.; Colombo, R.; Antinori, S.; Corbellino, M.; et al. Pulmonary post-mortem findings in a series of COVID-19 cases from northern Italy: A two-centre descriptive study. Lancet Infect. Dis. 2020, 20, 1135–1140. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Conway, E.M.; Pryzdial, E.L.G. Is the COVID-19 thrombotic catastrophe complement-connected? J. Thromb. Haemost. 2020, 18, 2812–2822. [Google Scholar] [CrossRef] [PubMed]
- Polycarpou, A.; Howard, M.; Farrar, C.A.; Greenlaw, R.; Fanelli, G.; Wallis, R.; Klavinskis, L.S.; Sacks, S. Rationale for targeting complement in COVID-19. EMBO Mol. Med. 2020, 12, e12642. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Yuan, X.; Chen, H.; Chaturvedi, S.; Braunstein, E.M.; Brodsky, R.A. Direct activation of the alternative complement pathway by SARS-CoV-2 spike proteins is blocked by factor D inhibition. Blood 2020, 136, 2080–2089. [Google Scholar] [CrossRef]
- Eriksson, O.; Hultstrom, M.; Persson, B.; Lipcsey, M.; Ekdahl, K.N.; Nilsson, B.; Frithiof, R. Mannose-Binding Lectin is Associated with Thrombosis and Coagulopathy in Critically Ill COVID-19 Patients. Thromb. Haemost. 2020, 120, 1720–1724. [Google Scholar] [CrossRef]
- Durigutto, P.; Macor, P.; Pozzi, N.; Agostinis, C.; Bossi, F.; Meroni, P.L.; Grossi, C.; Borghi, M.O.; Planer, W.; Garred, P.; et al. Complement Activation and Thrombin Generation by MBL Bound to beta2-Glycoprotein I. J. Immunol. 2020, 205, 1385–1392. [Google Scholar] [CrossRef]
- Rambaldi, A.; Gritti, G.; Mico, M.C.; Frigeni, M.; Borleri, G.; Salvi, A.; Landi, F.; Pavoni, C.; Sonzogni, A.; Gianatti, A.; et al. Endothelial injury and thrombotic microangiopathy in COVID-19: Treatment with the lectin-pathway inhibitor narsoplimab. Immunobiology 2020, 225, 152001. [Google Scholar] [CrossRef] [PubMed]
- Kanduc, D.; Shoenfeld, Y. Molecular mimicry between SARS-CoV-2 spike glycoprotein and mammalian proteomes: Implications for the vaccine. Immunol. Res. 2020, 68, 310–313. [Google Scholar] [CrossRef]
- Lyons-Weiler, J. Pathogenic priming likely contributes to serious and critical illness and mortality in COVID-19 via autoimmunity. J. Transl. Autoimmun. 2020, 3, 100051. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Kharrazian, D. Reaction of Human Monoclonal Antibodies to SARS-CoV-2 Proteins With Tissue Antigens: Implications for Autoimmune Diseases. Front. Immunol. 2020, 11, 617089. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-H.; Huang, Y.-H.; Chuang, Y.-H.; Peng, C.-M.; Wang, L.-C.; Lin, Y.-T.; Chiang, B.-L. Autoantibodies against human epithelial cells and endothelial cells after severe acute respiratory syndrome (SARS)-associated coronavirus infection. J. Med. Virol. 2005, 77, 1–7. [Google Scholar] [CrossRef]
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Liu, F.; Zheng, N.S.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse Functional Autoantibodies in Patients with COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Pfister, F.; Vonbrunn, E.; Ries, T.; Jack, H.M.; Uberla, K.; Lochnit, G.; Sheriff, A.; Herrmann, M.; Buttner-Herold, M.; Amann, K.; et al. Complement Activation in Kidneys of Patients With COVID-19. Front. Immunol. 2020, 11, 594849. [Google Scholar] [CrossRef]
- Su, H.; Yang, M.; Wan, C.; Yi, L.X.; Tang, F.; Zhu, H.Y.; Yi, F.; Yang, H.C.; Fogo, A.B.; Nie, X.; et al. Renal histopathological analysis of 26 postmortem findings of patients with COVID-19 in China. Kidney Int. 2020, 98, 219–227. [Google Scholar] [CrossRef]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Sonzogni, A.; Previtali, G.; Seghezzi, M.; Grazia Alessio, M.; Gianatti, A.; Licini, L.; Morotti, D.; Zerbi, P.; Carsana, L.; Rossi, R.; et al. Liver histopathology in severe COVID 19 respiratory failure is suggestive of vascular alterations. Liver Int. 2020, 40, 2110–2116. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cases | Age | Sex | Respiratory Signs * | Comorbidities | Radiologic Findings | Time from Diagnosis to Death (Days) | Hospitalization (Days) | Postmortem (Days) |
|---|---|---|---|---|---|---|---|---|
| 1 | 73 | M | yes | Prostate cancer; Lung metastases; Pyonephrosis; Heart failure | Bilateral airspace opacites | 24 | 23 | 1 |
| 2 | 82 | M | yes | Diabetes; Bladder cancer; Hypercholesterolemia | Bilateral airspace opacites | 49 | 18 | 2 |
| 3 | 80 | M | yes | Encephalopathy; Urosepsis; Seizure | Bilateral airspace opacites | 30 | 30 | 1 |
| 4 | 97 | F | yes | Colon cancer | NA | 3 | 14 | 1 |
| 5 | 90 | F | yes | Hypertension; Diabetes | Pleural effusion left lung | 48 | 48 | 1 |
| 6 | 77 | F | yes | Dementia; Stroke; Melanoma | Bilateral airspace opacites | 4 | 3 | 3 |
| 7 | 74 | F | yes | Seizure; Paraplegia; Atrial fibrillation | Ground glass right lung; left lung lower lobe consolidation | ND | 14 | 2 |
| 8 | 72 | M | yes | BPCO; heart failure; Diabetes; stroke | Bilateral airspace opacites | 27 | 27 | 2 |
| 9 | 79 | F | yes | Heart failure; Bilateral fibrotorax | Emphysema; Airspace opacities right lung | 27 | 27 | 2 |
| 10 | 80 | F | yes | Relapsing urinary infection; Stroke with left hemiparesis; Heart failure; FA; Hypercholesterolemia | Pleural effusion | 5 | 11 | 1 |
| 11 | 77 | F | yes | Bilateral carotid stenosis; Hypothyroidism; Obesity; Bladder and right breast cancer; Chronic vascular encephalopathy | Bilateral airspace opacites | 23 | 20 | 2 |
| 12 | 94 | M | yes | BPCO; Asbestosis; Hypertension; FA; History of relapsing falls | Bilateral calcific fibrothorax; Ground glass right lung; Interstitial pneumonia | 25 | 24 | 2 |
| Control A | 84 | F | yes | Anemia; Obesity; Previous DVT with massive pulmonary embolism; Chronic cholestasis; Vascular encephalopathy with cognitive impairment | Pulmonary embolism; Pleural effusion; Right lung consolidation | ND | 5 | 1 |
| Control B | 79 | F | yes | Hypertension; Hypertensive-ischemic heart disease; Diabetes; Stroke; Esophagitis | Mild and widespread interstitial disease | ND | 7 | 2 |
| Control C | 95 | M | yes | COPD; Hypertension; FA; Cognitive impairment; MGUS | Right lung consolidation; Bilateral pleural effusion | ND | 24 | 2 |
| Cases | D-Dimer (µg/L) | CRP (mg/dL) | Ferritin (µg/L) | White Cells (n × 103/µL) | Neutrophils (n × 103/µL) | Lymphocytes (n × 103/µL) | Platelets (n × 103/µL) |
|---|---|---|---|---|---|---|---|
| 1 | 17,180 | 15.6 | 2059 | 14.59 | 6.72 | 0.52 | 173 |
| 2 | 3020 | 30.6 | 2892 | 15.13 | 11.25 | 1.65 | 256 |
| 3 | 1330 | 30.2 | 90.9 | 12.98 | 9.59 | 2.45 | 2.84 |
| 4 | 1330 | 302.8 | 352 | 20.83 | 6.13 | 0.82 | 239 |
| 5 | NA | 9.1 | 85 | 12.68 | 11.21 | 0.43 | 461 |
| 6 | 6800 | 16.08 | 1608 | 5.57 | 3.89 | 0.76 | 231 |
| 7 | 2120 | 3.82 | 882 | 14.03 | 6.57 | 1.15 | 259 |
| 8 | 1800 | 210 | 755 | 14.19 | 13.51 | 0.31 | 83 |
| 9 | 430 | 15.7 | 1035 | 23.03 | 3.52 | 1.82 | 326 |
| 10 | NA | 4.97 | 244.6 | 7.69 | 5.74 | 2.19 | 355 |
| 11 | 1046 | 30.48 | 2146 | 17.79 | 19.91 | 1.26 | 222 |
| 12 | 1536 | 1.85 | 294.8 | 4.8 | 7.34 | 0.34 | 104 |
| Controls | <500 | <0.05 | 30–400 | 4.8–10.8 | 1.5–6.5 | 1.2–3.4 | 130–430 |
| P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 | P12 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IgG | ||||||||||||||||||||
| C1q | P | Postive | ||||||||||||||||||
| MBL | ||||||||||||||||||||
| Factor B | W | Weakly positive | ||||||||||||||||||
| C4 | ||||||||||||||||||||
| C3 | N | Negative | ||||||||||||||||||
| C5b-9 | ||||||||||||||||||||
| General C activation | ||||||||||||||||||||
| P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 | P12 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IgG | P | Postive | ||||||||||||||||||
| C1q | ||||||||||||||||||||
| C4 | W | Weakly positive | ||||||||||||||||||
| C3 | ||||||||||||||||||||
| C5b-9 | N | Negative | ||||||||||||||||||
| General C activation | ||||||||||||||||||||
| P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 | P12 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IgG | P | Postive | ||||||||||||||||||
| C1q | ||||||||||||||||||||
| C4 | W | Weakly positive | ||||||||||||||||||
| C3 | ||||||||||||||||||||
| C5b-9 | N | Negative | ||||||||||||||||||
| General C activation | ||||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macor, P.; Durigutto, P.; Mangogna, A.; Bussani, R.; De Maso, L.; D’Errico, S.; Zanon, M.; Pozzi, N.; Meroni, P.L.; Tedesco, F. Multiple-Organ Complement Deposition on Vascular Endothelium in COVID-19 Patients. Biomedicines 2021, 9, 1003. https://doi.org/10.3390/biomedicines9081003
Macor P, Durigutto P, Mangogna A, Bussani R, De Maso L, D’Errico S, Zanon M, Pozzi N, Meroni PL, Tedesco F. Multiple-Organ Complement Deposition on Vascular Endothelium in COVID-19 Patients. Biomedicines. 2021; 9(8):1003. https://doi.org/10.3390/biomedicines9081003
Chicago/Turabian StyleMacor, Paolo, Paolo Durigutto, Alessandro Mangogna, Rossana Bussani, Luca De Maso, Stefano D’Errico, Martina Zanon, Nicola Pozzi, Pier Luigi Meroni, and Francesco Tedesco. 2021. "Multiple-Organ Complement Deposition on Vascular Endothelium in COVID-19 Patients" Biomedicines 9, no. 8: 1003. https://doi.org/10.3390/biomedicines9081003
APA StyleMacor, P., Durigutto, P., Mangogna, A., Bussani, R., De Maso, L., D’Errico, S., Zanon, M., Pozzi, N., Meroni, P. L., & Tedesco, F. (2021). Multiple-Organ Complement Deposition on Vascular Endothelium in COVID-19 Patients. Biomedicines, 9(8), 1003. https://doi.org/10.3390/biomedicines9081003