
3D Collagen Hydrogel Promotes In Vitro Langerhans Islets Vascularization through ad-MVFs Angiogenic Activity

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of ad-MVFs from Rat Adipose Tissue
2.2. ad-MVF Quantification
2.3. ad-MVF Culture Conditions, Viability, and Proliferation
2.4. Isolation of Pancreatic Islets
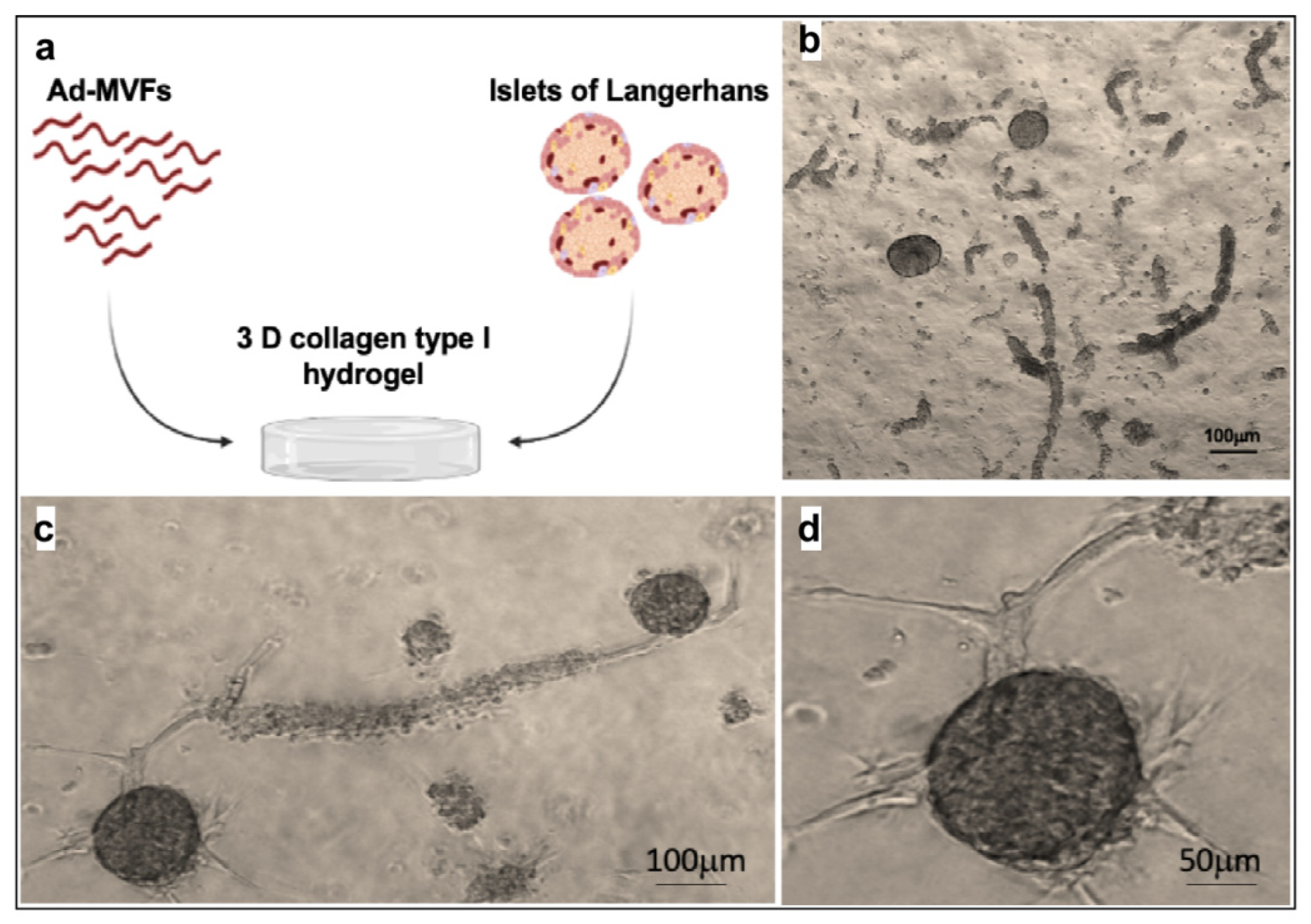
2.5. Coculture of Islets and ad-MFVs in Collagen Type-I Hydrogel
2.6. Glucose Stimulated Insulin Secretion (GSIS) Assay
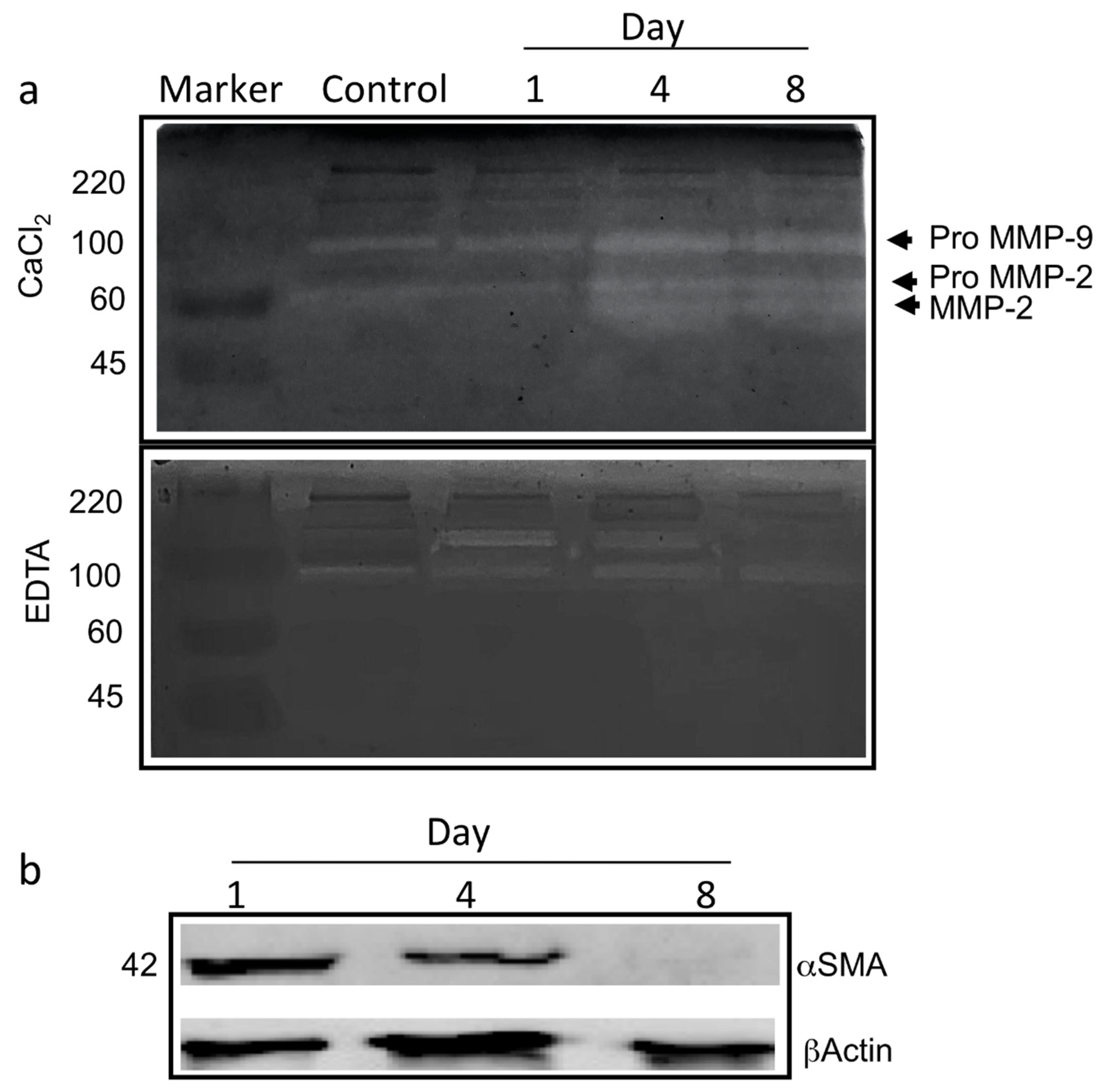
2.7. Western Blots
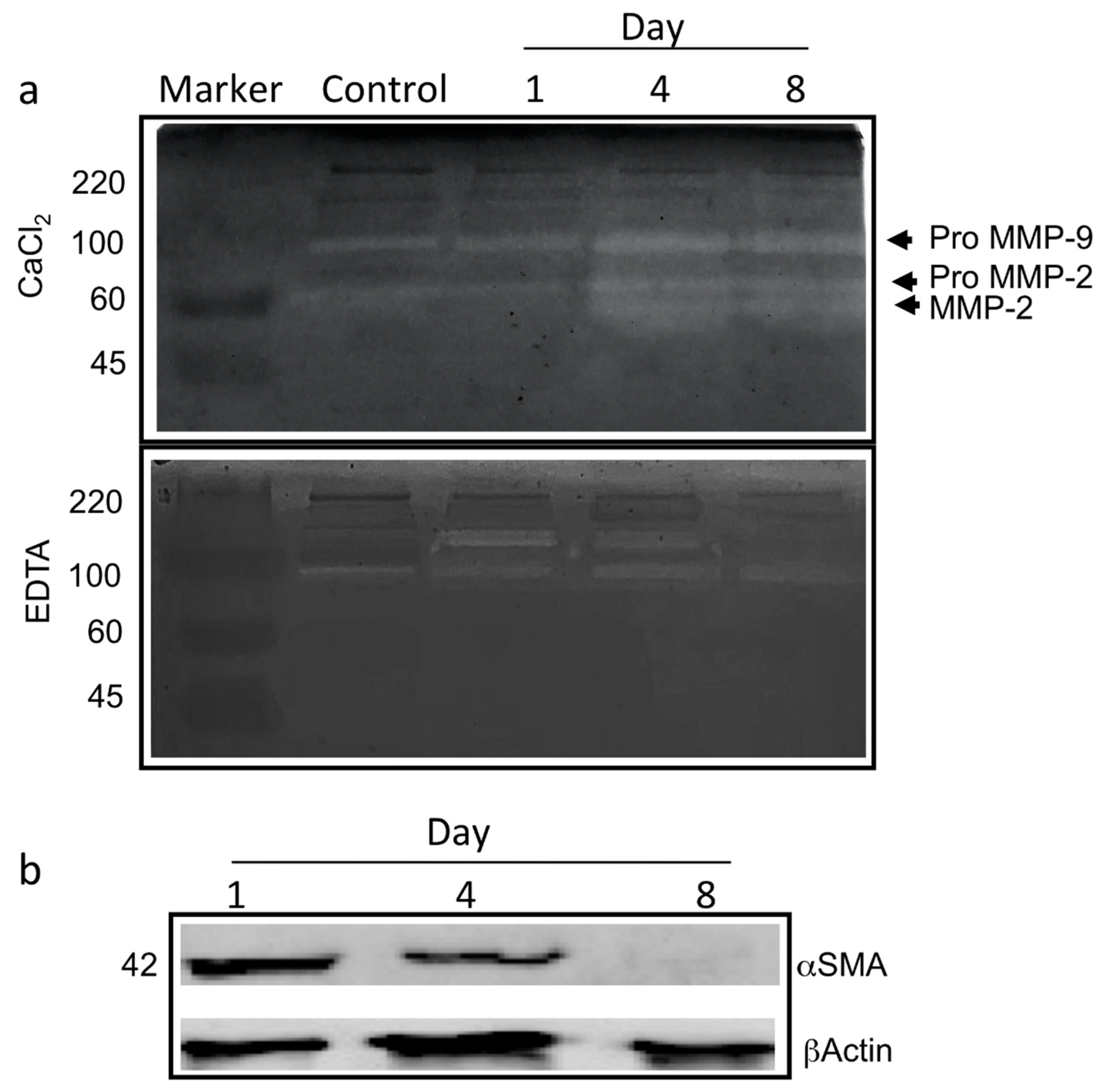
2.8. Zymography
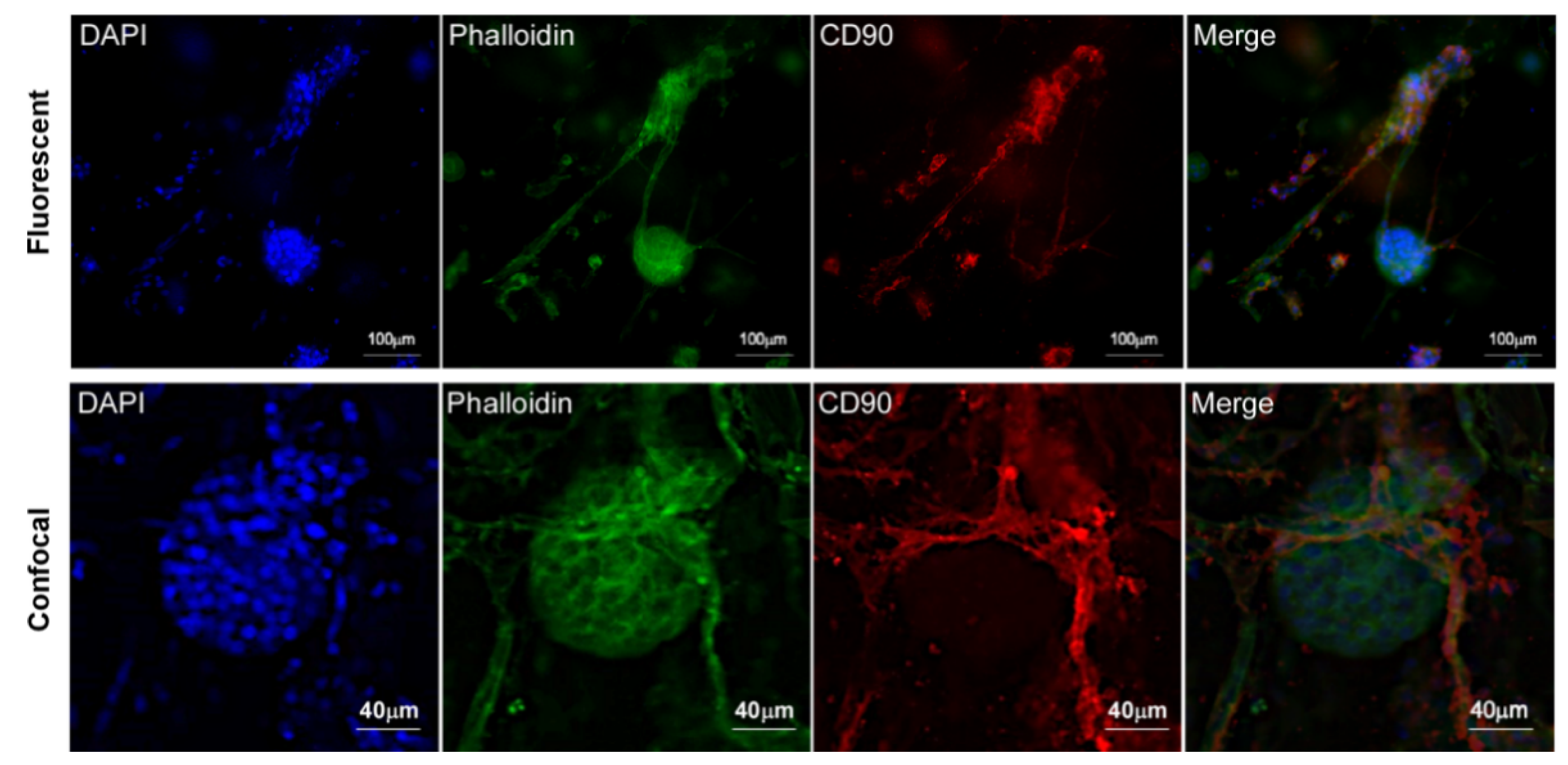
2.9. Immunofluorescence Assay
2.10. RNA Isolation and cDNA Synthesis
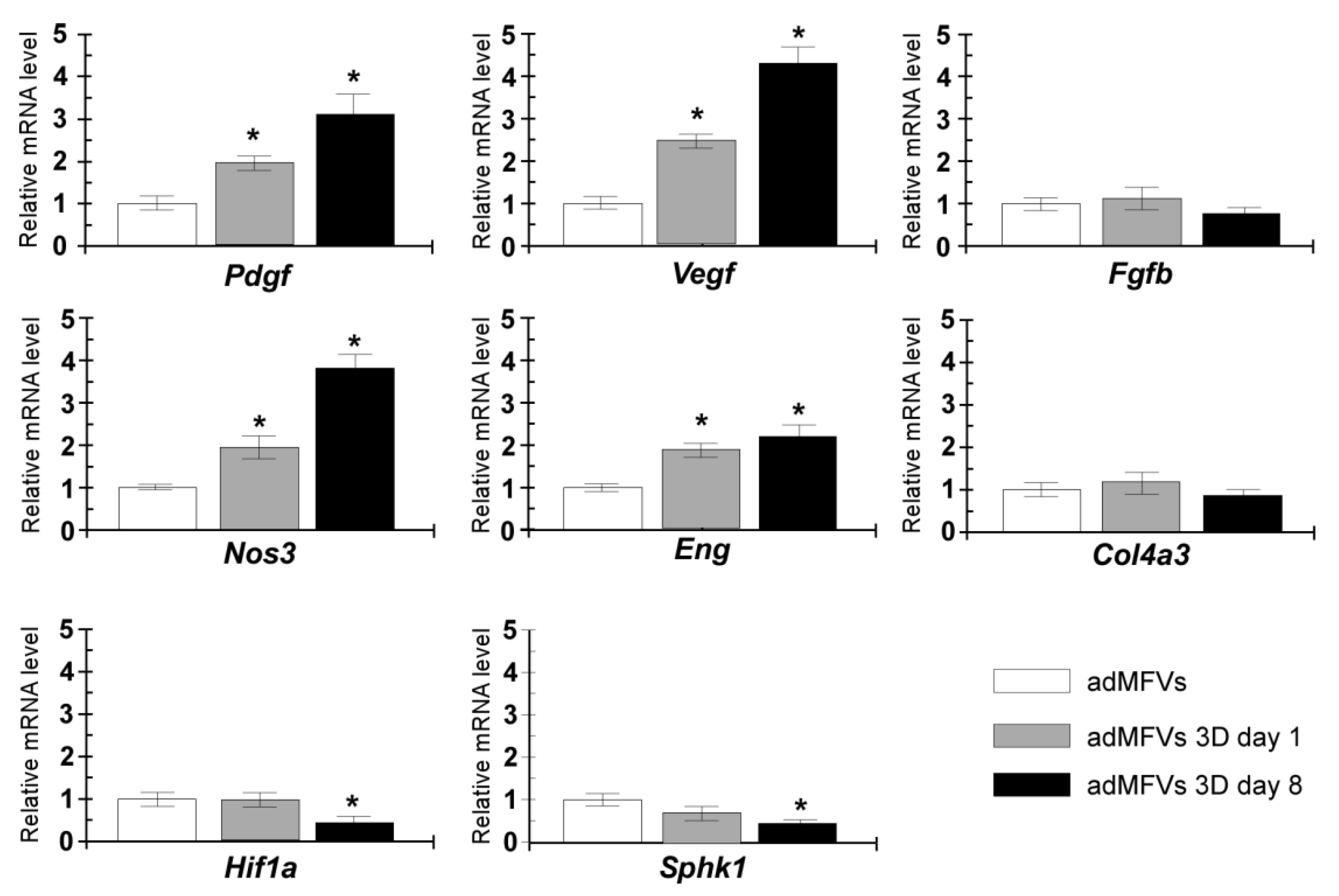
2.11. RT-qPCR Analyses
3. Results
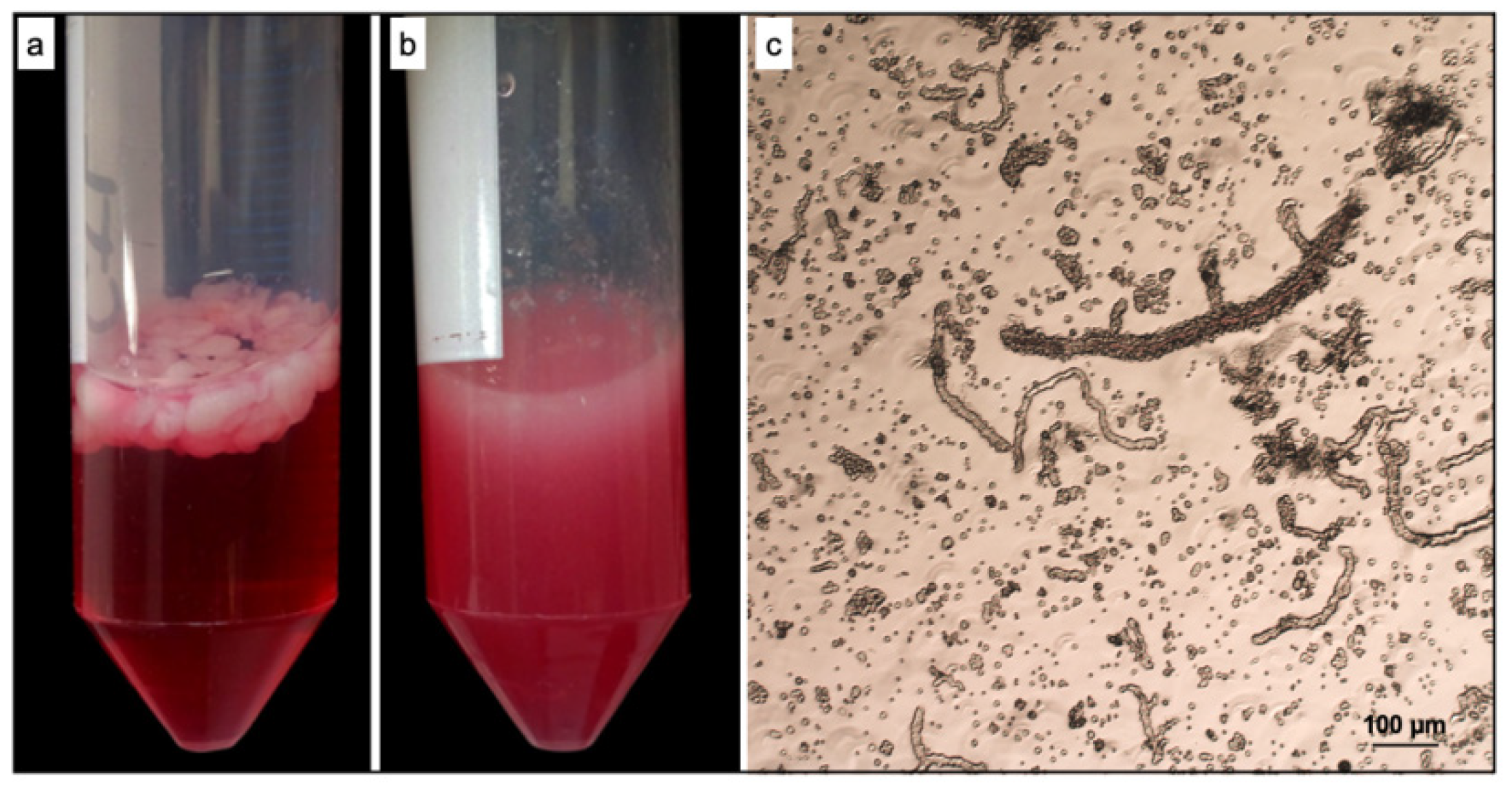
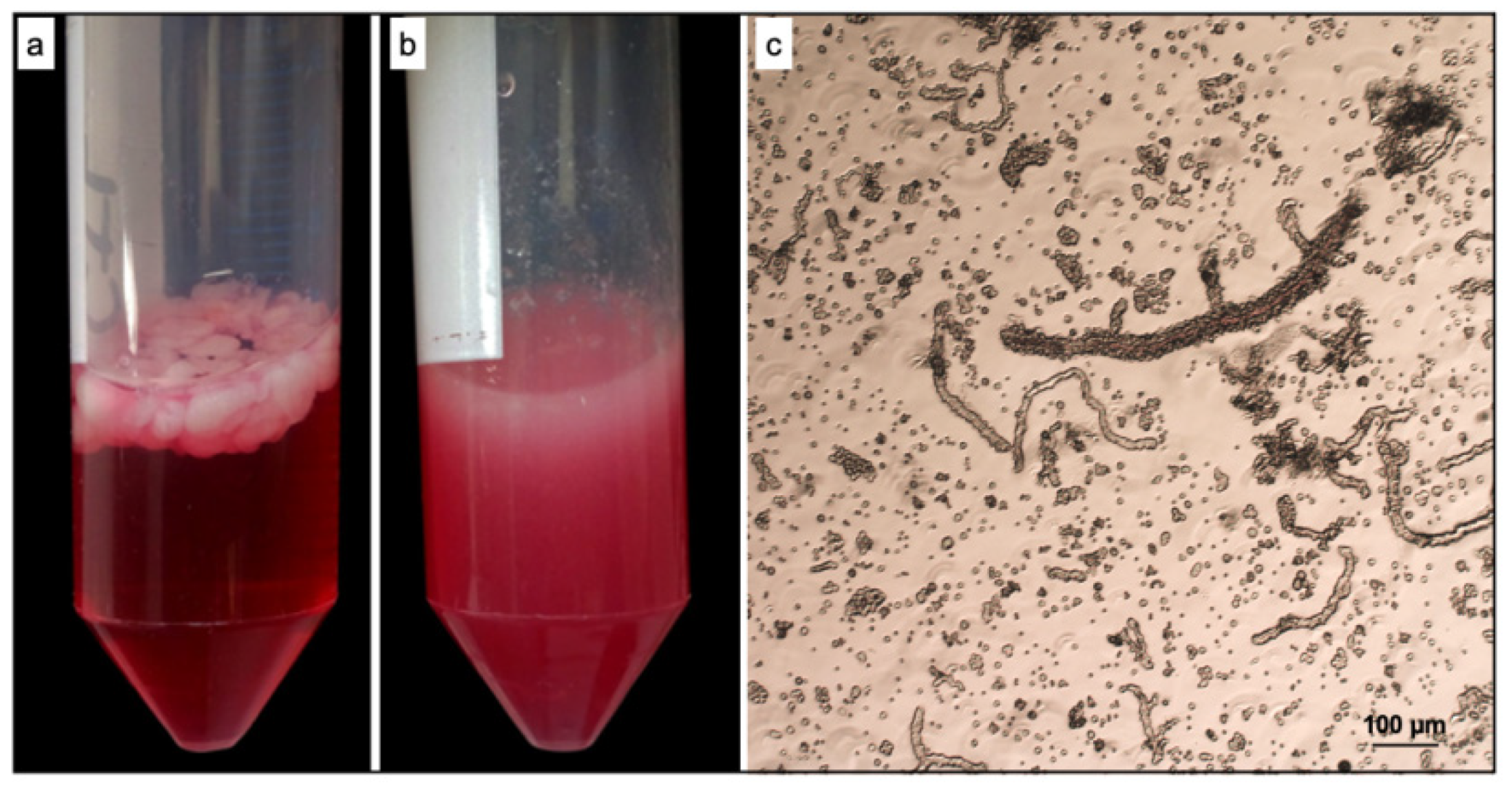
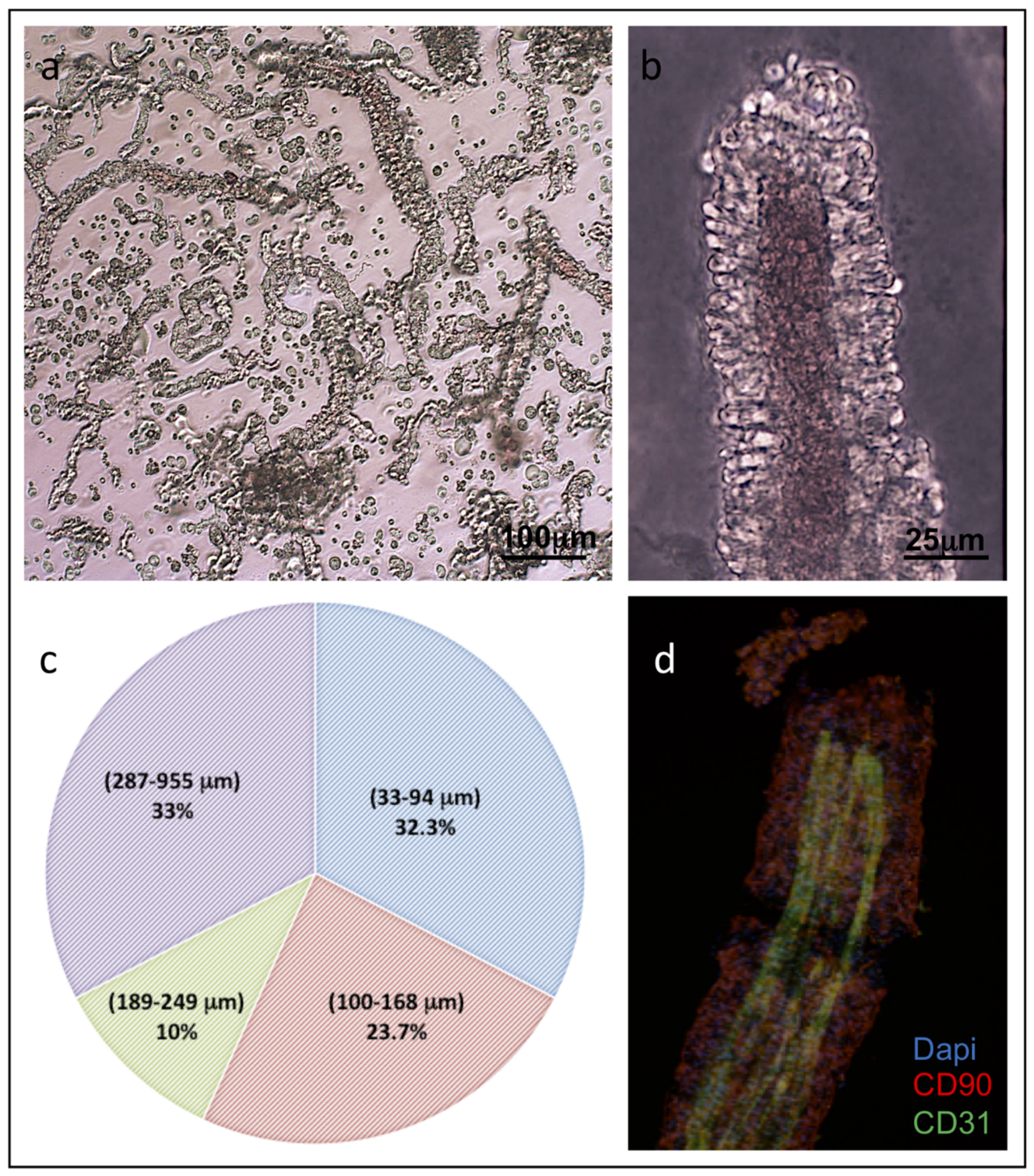
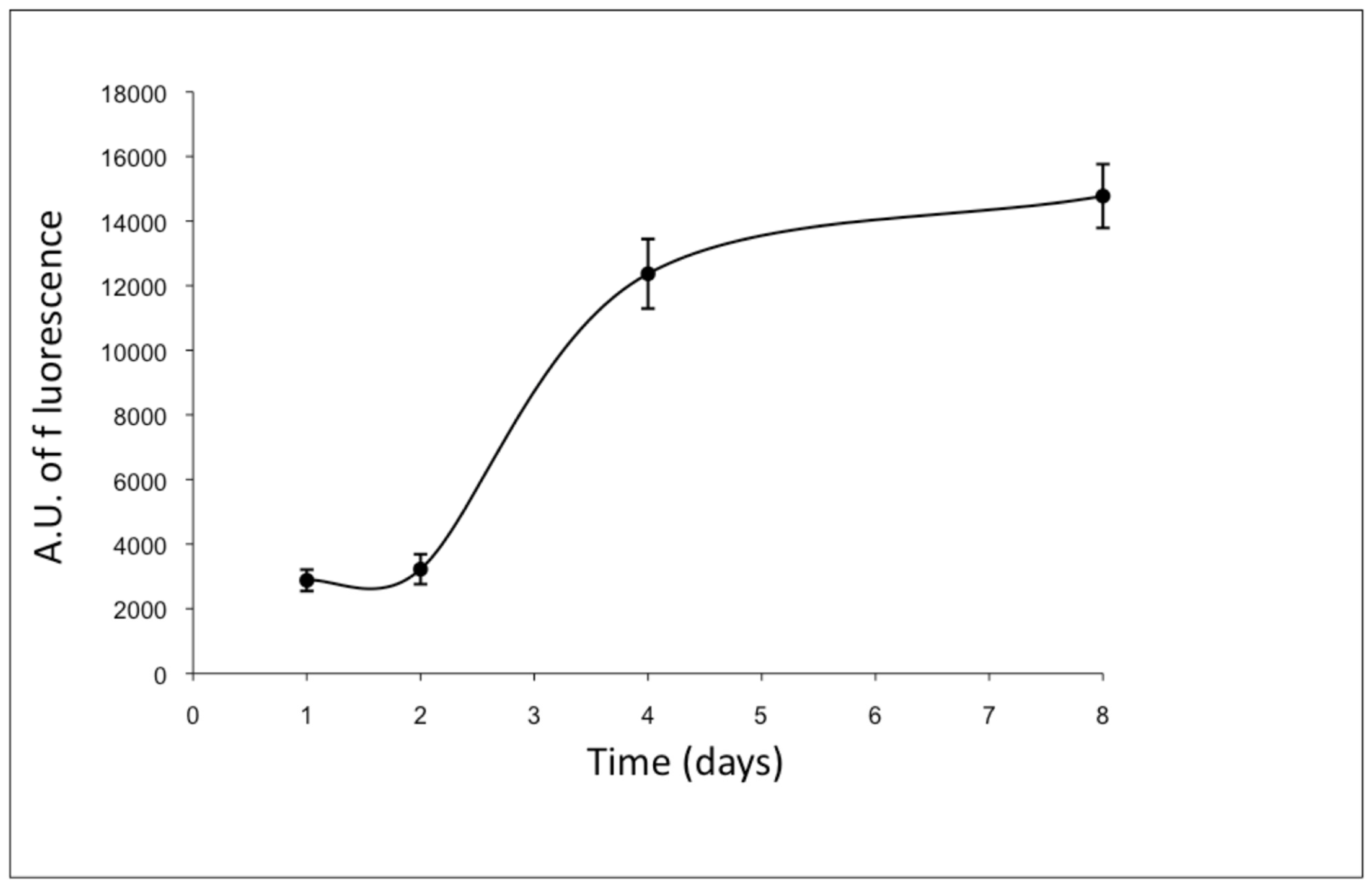
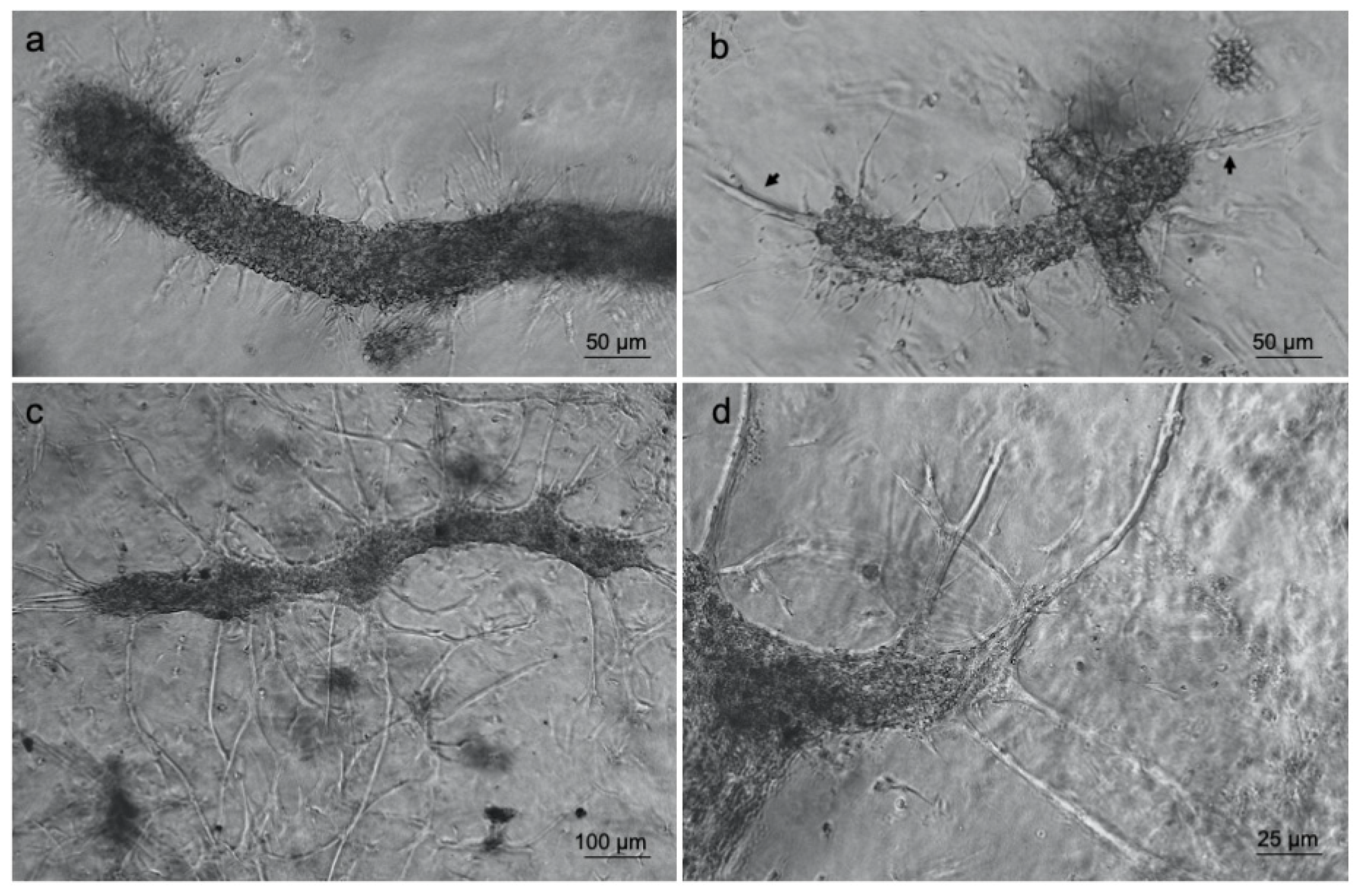
3.1. ad-MVF Isolation and Culture in 3D Collagen Type-I Hydrogel
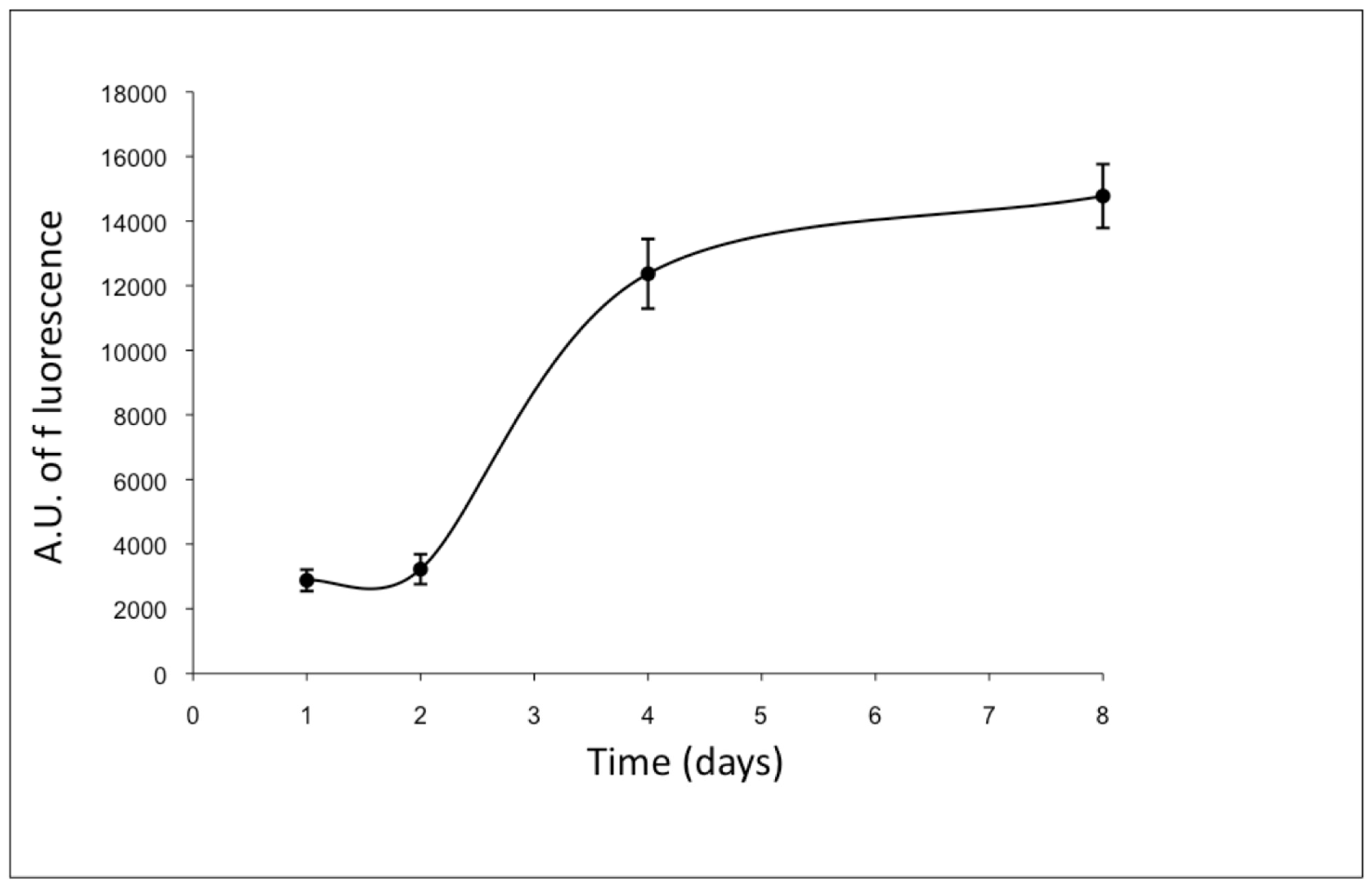
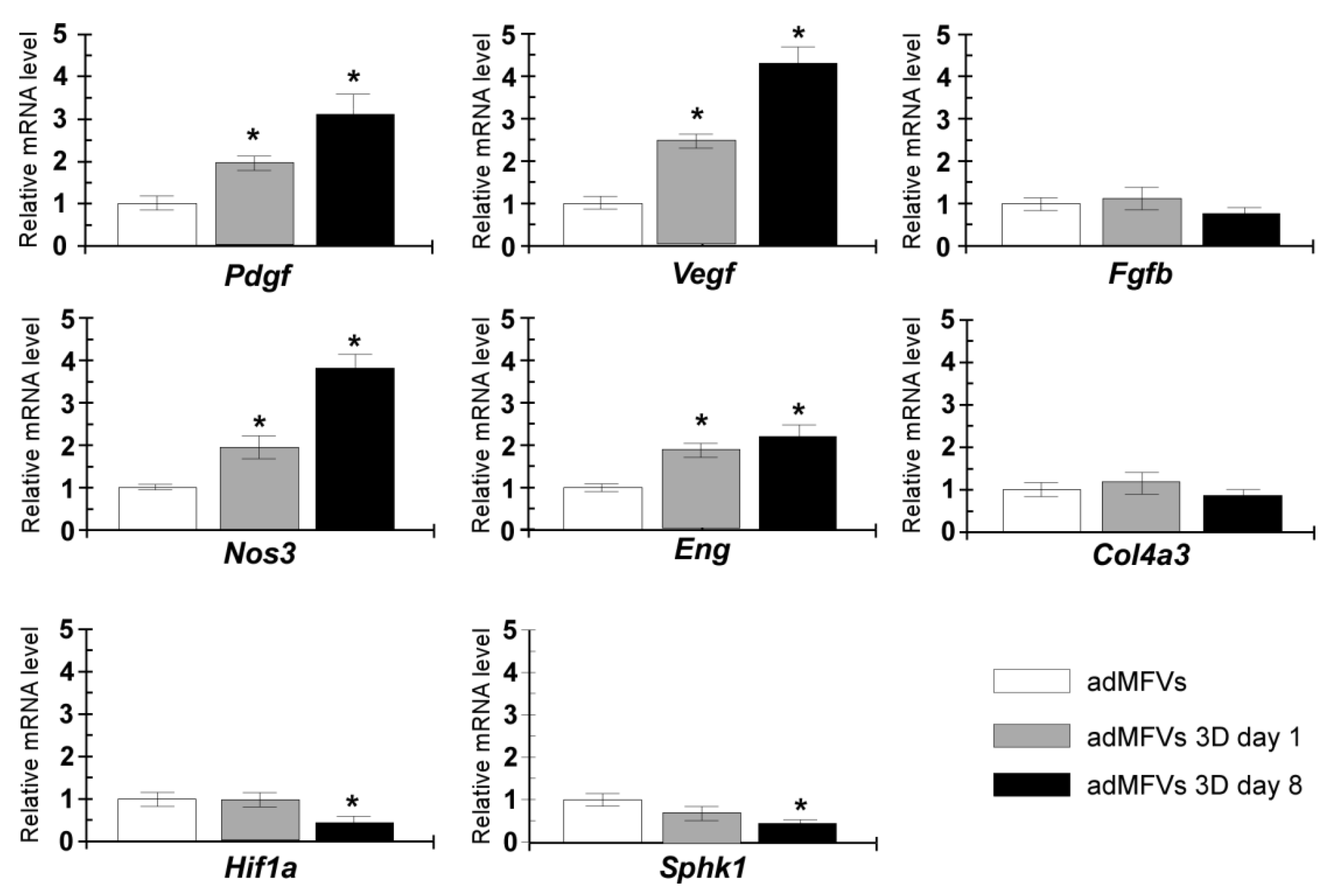
3.2. Gene Expression Analyses of ad-MVFs in 3D Collagen Type-I Hydrogel
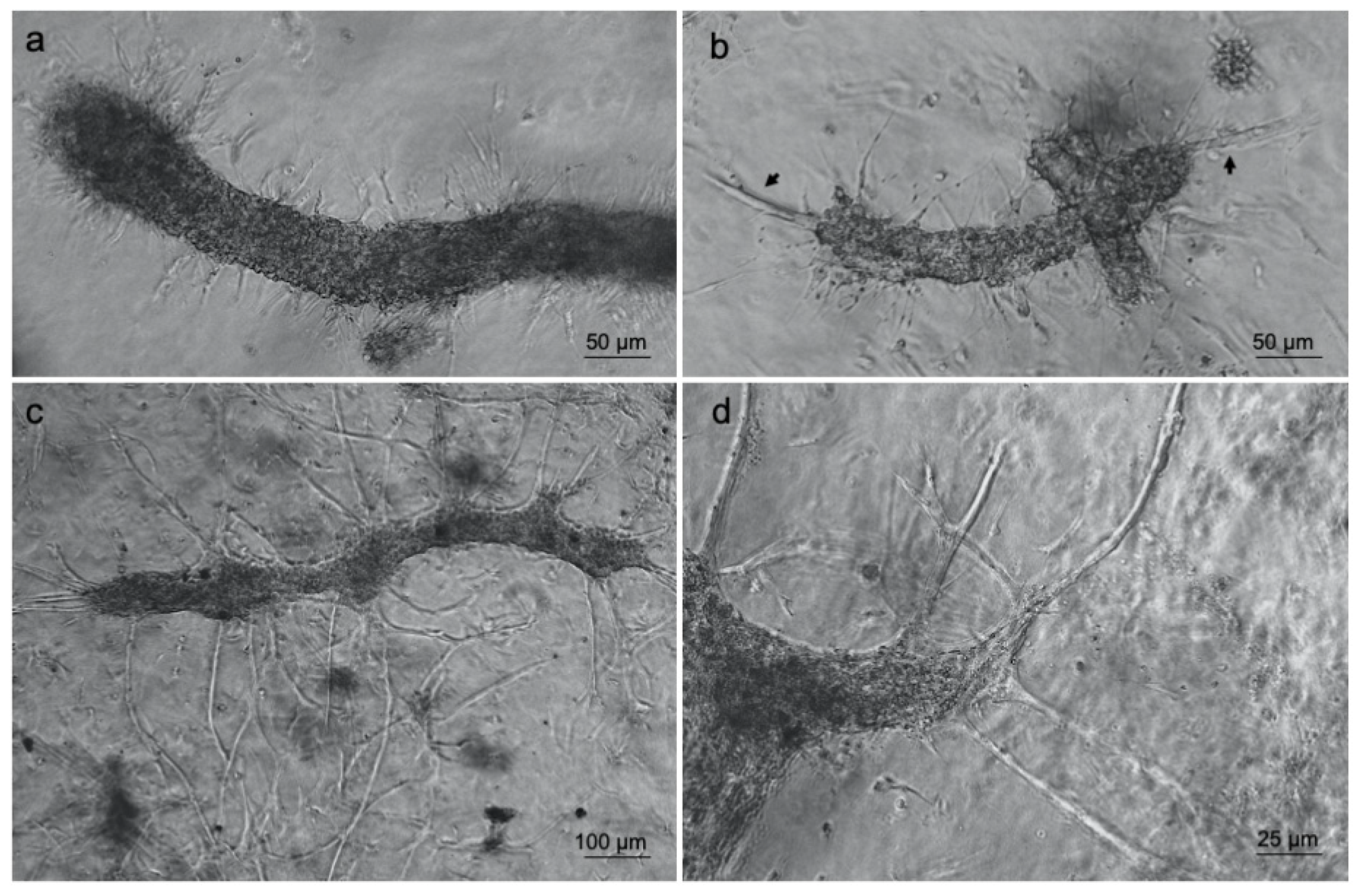
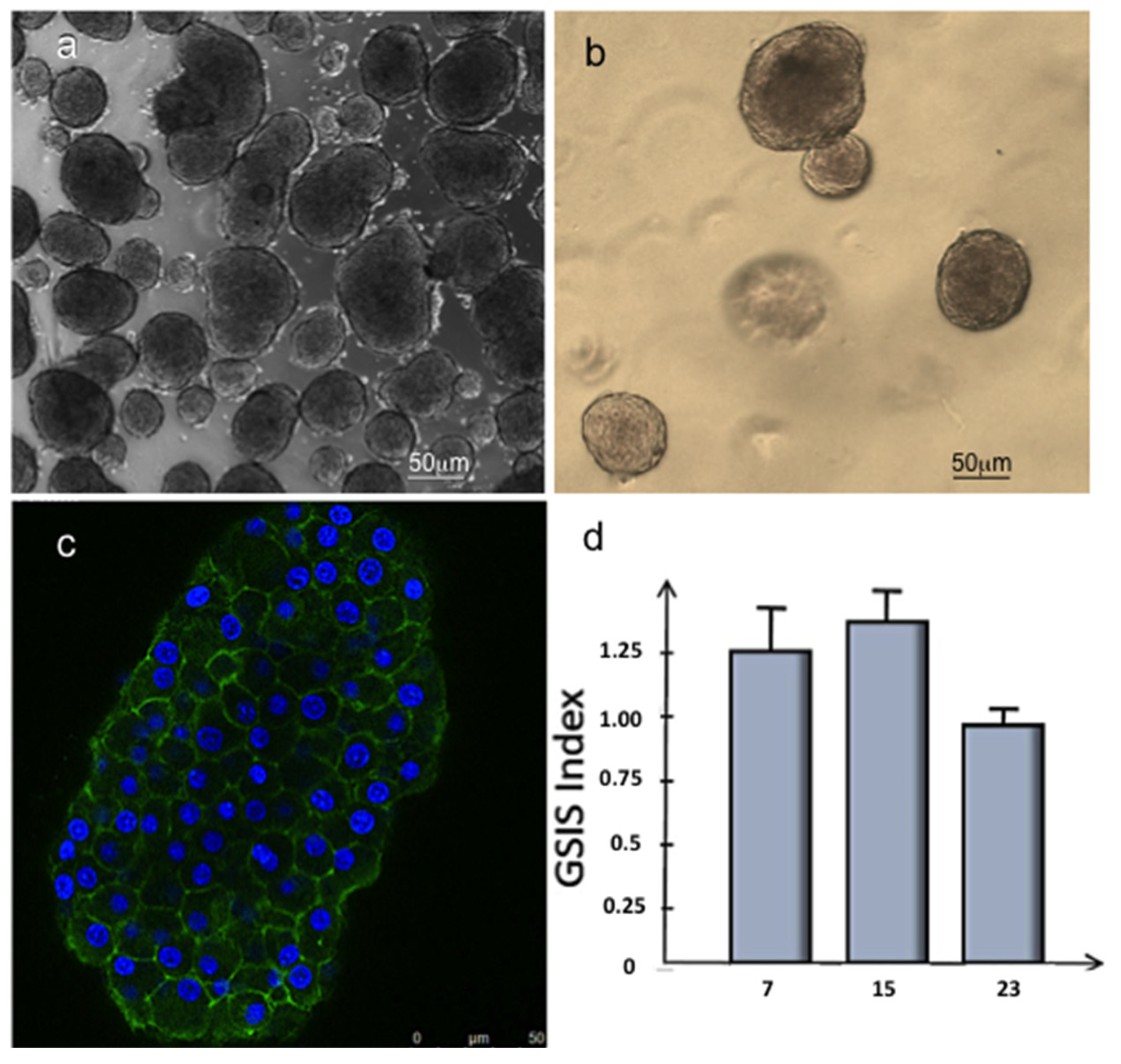
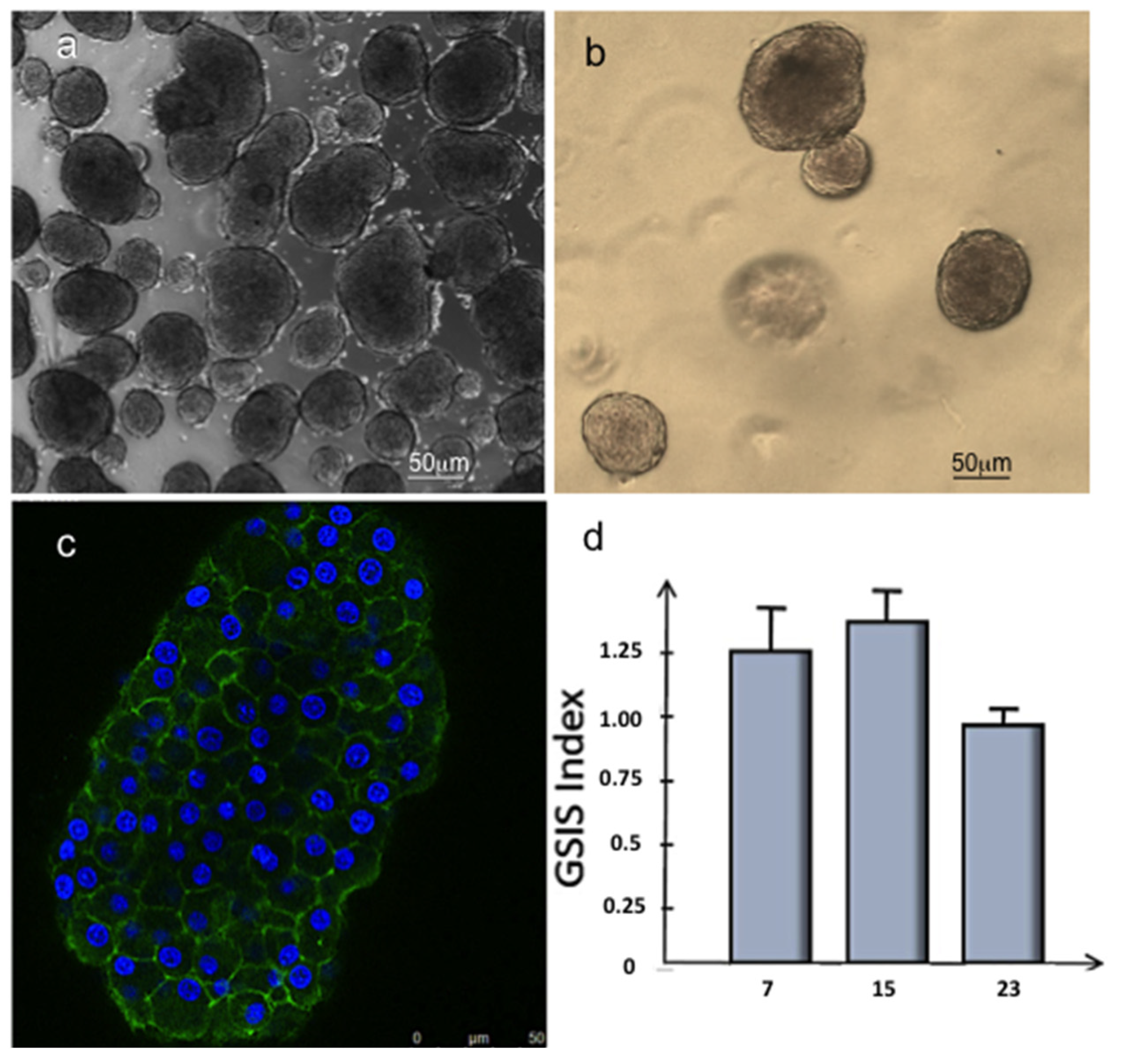
3.3. Islet of Langerhans and Ad-SVF Coculture into Type-I Collagen Hydrogel
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kinney, M.A.; Hookway, T.A.; Wang, Y.; McDevitt, T.C. Engineering three-dimensional stem cell morphogenesis for the development of tissue models and scalable regenerative therapeutics. Ann. Biomed. Eng. 2014, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Antoine, E.E.; Vlachos, P.P.; Rylander, M.N. Review of collagen i hydrogels for bioengineered tissue microenvironments: Characterization of mechanics, structure, and transport. Tissue Eng. Part B Rev. 2014, 20, 683–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, L.J.; Binner, M.; Freudenberg, U.; Werner, C. Hydrogel-based in vitro models of tumor angiogenesis. Methods Mol. Biol. 2017, 39–63. [Google Scholar] [CrossRef]
- Yang, J.A.; Yeom, J.; Hwang, B.W.; Hoffman, A.S.; Hahn, S.K. In situ-forming injectable hydrogels for regenerative medicine. Prog. Polym. Sci. 2014, 39, 1973–1986. [Google Scholar] [CrossRef]
- Rigogliuso, S.; Salamone, M.; Barbarino, E.; Barbarino, M.; Nicosia, A.; Ghersi, G. Production of Injectable Marine Collagen-Based Hydrogel for the Maintenance of Differentiated Chondrocytes in Tissue Engineering Applications. Int. J. Mol. Sci. 2020, 21, 5798. [Google Scholar] [CrossRef]
- Rastogi, P.; Kandasubramanian, B. Review of alginate-based hydrogel bioprinting for application in tissue engineering. Biofabrication 2019, 11. [Google Scholar] [CrossRef]
- Sharma, S.; Tiwari, S. A review on biomacromolecular hydrogel classification and its applications. Int. J. Biol. Macromol. 2020, 162, 737–747. [Google Scholar] [CrossRef]
- Chen, S.; Jang, T.-S.; Pan, H.M.; Jung, H.-D.; Sia, M.W.; Xie, S.; Hang, Y.; Chong, S.K.M.; Wang, D.; Song, J. 3D Freeform Printing of Nanocomposite Hydrogels through in situ Precipitation in Reactive Viscous Fluid. Int. J. Bioprinting 2020, 6, 258. [Google Scholar] [CrossRef]
- Cai, M.H.; Chen, X.Y.; Fu, L.Q.; Du, W.L.; Yang, X.; Mou, X.Z.; Hu, P.Y. Design and Development of Hybrid Hydrogels for Biomedical Applications: Recent Trends in Anticancer Drug Delivery and Tissue Engineering. Front. Bioeng. Biotechnol. 2021, 9, 630943. [Google Scholar] [CrossRef]
- Goers, L.; Freemont, P.; Polizzi, K.M. Co-culture systems and technologies: Taking synthetic biology to the next level. J.R. Soc. Interface. 2014, 11. [Google Scholar] [CrossRef] [Green Version]
- Tam, R.Y.; Smith, L.J.; Shoichet, M.S. Engineering Cellular Microenvironments with Photo- and Enzymatically Responsive Hydrogels: Toward Biomimetic 3D Cell Culture Models. Acc. Chem. Res. 2017, 50, 703–713. [Google Scholar] [CrossRef]
- Wang, L.S.; Chung, J.E.; Chan, P.P.; Kurisawa, M. Injectable biodegradable hydrogels with tunable mechanical properties for the stimulation of neurogenesic differentiation of human mesenchymal stem cells in 3D culture. Biomaterials 2010, 31, 1148–1157. [Google Scholar] [CrossRef]
- Parenteau-Bareil, R.; Gauvin, R.; Berthod, F. Collagen-Based Biomaterials for Tissue Engineering Applications. Materials 2010, 3, 1863–1887. [Google Scholar] [CrossRef] [Green Version]
- Laschke, M.W.; Menger, M.D. Prevascularization in tissue engineering: Current concepts and future directions. Biotechnol. Adv. 2016, 34, 112–121. [Google Scholar] [CrossRef]
- Vaithilingam, J.; Sanjuan-Alberte, P.; Campora, S.; Rance, G.A.; Jiang, L.; Thorpe, J.; Burroughs, L.; Tuck, C.J.; Denning, C.; Wildman, R.D.; et al. Multifunctional Bioinstructive 3D Architectures to Modulate Cellular Behavior. Adv. Funct. Mater. 2019. [Google Scholar] [CrossRef] [Green Version]
- Traktuev, D.O.; Merfeld-Clauss, S.; Li, J.; Kolonin, M.; Arap, W.; Pasqualini, R.; Johnstone, B.H.; March, K.L. A population of multipotent CD34-positive adipose stromal cells share pericyte and mesenchymal surface markers, reside in a periendothelial location, and stabilize endothelial networks. Circ. Res. 2008, 102, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pill, K.; Melke, J.; Mühleder, S.; Pultar, M.; Rohringer, S.; Priglinger, E.; Redl, H.R.; Hofmann, S.; Holnthoner, W. Microvascular Networks From Endothelial Cells and Mesenchymal Stromal Cells From Adipose Tissue and Bone Marrow: A. Comparison. Front. Bioeng. Biotechnol. 2018, 6, 156. [Google Scholar] [CrossRef] [PubMed]
- Levenberg, S.; Rouwkema, J.; Macdonald, M.; Garfein, E.S.; Kohane, D.S.; Darland, D.C.; Marini, R.; van Blitterswijk, C.A.; Mulligan, R.C.; D’Amore, P.A.; et al. Engineering vascularized skeletal muscle tissue. Nat. Biotechnol. 2005, 23, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Blinder, Y.J.; Freiman, A.; Raindel, N.; Mooney, D.J.; Levenberg, S. Vasculogenic dynamics in 3D engineered tissue constructs. Sci. Rep. 2015, 5, 17840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamat, P.; Frueh, F.S.; McLuckie, M.; Sanchez-Macedo, N.; Wolint, P.; Lindenblatt, N.; Plock, J.A.; Calcagni, M.; Buschmann, J. Adipose tissue and the vascularization of biomaterials: Stem cells, microvascular fragments and nanofat—A review. Cytotherapy 2020, 22, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Cerino, G.; Gaudiello, E.; Muraro, M.G.; Eckstein, F.; Martin, I.; Scherberich, A.; Marsano, A. Engineering of an angiogenic niche by perfusion culture of adipose-derived stromal vascular fraction cells. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Laschke, M.W.; Menger, M.D. Adipose tissue-derived microvascular fragments: Natural vascularization units for regenerative medicine. Trends Biotechnol. 2015, 33, 442–448. [Google Scholar] [CrossRef]
- Später, T.; Körbel, C.; Frueh, F.S.; Nickels, R.M.; Menger, M.D.; Laschke, M.W. Seeding density is a crucial determinant for the in vivo vascularisation capacity of adipose tissue-derived microvascular fragments. Eur. Cells Mater. 2017, 34, 55–69. [Google Scholar] [CrossRef]
- McDaniel, J.S.; Pilia, M.; Ward, C.L.; Pollot, B.E.; Rathbone, C.R. Characterization and multilineage potential of cells derived from isolated microvascular fragments. J. Surg. Res. 2014, 192, 214–222. [Google Scholar] [CrossRef]
- Grässer, C.; Scheuer, C.; Parakenings, J.; Tschernig, T.; Eglin, D.; Menger, M.D.; Laschke, M.W. Effects of macrophage-activating lipopeptide-2 (MALP-2) on the vascularisation of implanted polyurethane scaffolds seeded with microvascular fragments. Eur. Cells Mater. 2016, 32, 74–86. [Google Scholar] [CrossRef]
- Frueh, F.S.; Später, T.; Lindenblatt, N.; Calcagni, M.; Giovanoli, P.; Scheuer, C.; Menger, M.D.; Laschke, M.W. Adipose Tissue-Derived Microvascular Fragments Improve Vascularization, Lymphangiogenesis, and Integration of Dermal Skin Substitutes. J. Invest. Dermatol. 2017, 137, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, B.R.; Hoying, J.B.; Williams, S.K. Microvascular transplantation after acute myocardial infarction. Tissue Eng. 2007, 13, 2871–2879. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.; Rathbone, C.R. Microvascular fragment transplantation improves rat dorsal skin flap survival. Plast. Reconstr. Surg. Glob. Open. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Pilia, M.; McDaniel, J.S.; Guda, T.; Chen, X.K.; Rhoads, R.P.; Allen, R.E.; Corona, B.T.; Rathbone, C.R. Transplantation and perfusion of microvascular fragments in a rodent model of volumetric muscle loss injury. Eur. Cells Mater. 2014, 28, 11–24. [Google Scholar] [CrossRef]
- Hiscox, A.M.; Stone, A.L.; Limesand, S.; Hoying, J.B.; Williams, S.K. An islet-stabilizing implant constructed using a preformed vasculature. Tissue Eng. Part A 2008, 14, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Rickels, M.R.; Robertson, R.P. Pancreatic islet transplantation in humans: Recent progress and future directions. Endocr. Rev. 2019, 40, 631–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, J.D.; Headen, D.M.; Aquart, J.; Johnson, C.T.; Shea, L.D.; Shirwan, H.; García, A.J. Vasculogenic hydrogel enhances islet survival, engraftment, and function in leading extrahepatic sites. Sci. Adv. 2017, 3, e1700184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarreal, D.; Pradhan, G.; Wu, C.S.; Allred, C.D.; Guo, S.; Sun, Y. A simple high efficiency protocol for pancreatic islet isolation from mice. J. Vis. Exp. 2019, 2019, e57048. [Google Scholar] [CrossRef] [PubMed]
- Salamone, M.; Cuttitta, A.; Bertuzzi, F.; Ricordi, C.; Ghersi, G.; Seidita, G. Biochemical comparison between Clostridium hystoliticum collagenases G and H obtained by DNA recombinant and extractive procedures. Chem. Eng. Trans. 2012, 259–264. [Google Scholar] [CrossRef]
- Campora, S.; Alberte, P.S.; Bruno, C.; Ghersi, G. Isolation of adult rat cardiomyocytes using recombinant collagenases. Chem. Eng. Trans. 2018. [Google Scholar] [CrossRef]
- Salamone, M.; Nicosia, A.; Bennici, C.; Quatrini, P.; Catania, V.; Mazzola, S.; Ghersi, G.; Cuttitta, A. Comprehensive analysis of a Vibrio parahaemolyticus strain extracellular serine protease VpSP37. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Salamone, M.; Saladino, S.; Pampalone, M.; Campora, S.; Ghersi, G. Tissue dissociation and primary cells isolation using recombinant collagenases class i and II. Chem. Eng. Trans. 2014. [Google Scholar] [CrossRef]
- Volpe, L.; Salamone, M.; Giardina, A.; Ghersi, G. Optimization of a Biotechnological Process for Production and Purification of Two Recombinant Proteins: Col, G. and Col H. Chem. Eng. Trans. 2016, 49, 61–66. [Google Scholar] [CrossRef]
- Salamone, M.; Seidita, G.; Cuttitta, A.; Rigogliuso, S.; Mazzola, S.; Bertuzzi, F.; Ghersi, G. A new method to value efficiency of enzyme blends for pancreatic tissue digestion. Transpl. Proc. 2010, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Tabatabaei, A.; Jalili, R.B.; Hartwell, R.; Salimi, S.; Kilani, R.T.; Ghahary, A. Embedding Islet in a Liquid Scaffold Increases Islet Viability and Function. Can. J. Diabetes. 2013, 37, 27–35. [Google Scholar] [CrossRef]
- Frueh, F.S.; Später, T.; Scheuer, C.; Menger, M.D.; Laschke, M.W. Isolation of murine adipose tissue-derived microvascular fragments as vascularization units for tissue engineering. J. Vis. Exp. 2017, 2017, 55721. [Google Scholar] [CrossRef] [PubMed]
- Salamone, M.; Rigogliuso, S.; Nicosia, A.; Tagliavia, M.; Campora, S.; Cinà, P.; Bruno, C.; Ghersi, G. Neural Crest-Derived Chondrocytes Isolation for Tissue Engineering in Regenerative Medicine. Cells 2020, 9, 962. [Google Scholar] [CrossRef] [PubMed]
- Knight, E.; Przyborski, S. Advances in 3D cell culture technologies enabling tissue-like structures to be created In Vitro. J. Anat. 2015, 227, 746–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandekeere, S.; Dewerchin, M.; Carmeliet, P. Angiogenesis Revisited: An Overlooked Role of Endothelial Cell Metabolism in Vessel Sprouting. Microcirculation 2015, 22, 509–517. [Google Scholar] [CrossRef]
- Folkman, J.; Shing, Y. Angiogenesis. J. Biol. Chem. 1992, 267, 10931–10934. [Google Scholar] [CrossRef]
- Arpino, V.; Brock, M.; Gill, S.E. The role of TIMPs in regulation of extracellular matrix proteolysis. Matrix Biol. 2015, 46, 247–254. [Google Scholar] [CrossRef]
- Rundhaug, J.E. Matrix metalloproteinases and angiogenesis. J. Cell. Mol. Med. 2005, 9, 267–285. [Google Scholar] [CrossRef]
- Nicosia, A.; Maggio, T.; Costa, S.; Salamone, M.; Tagliavia, M.; Mazzola, S.; Gianguzza, F.; Cuttitta, A. Maintenance of a protein structure in the dynamic evolution of TIMPs over 600 million years. Genome Biol. Evol. 2016, 8, 1056–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salamone, M.; Pavia, F.C.; Ghersi, G. Proteolytic Enzymes Clustered in Specialized Plasma-Membrane Domains Drive Endothelial Cells’ Migration. PLoS ONE 2016, 11, e0154709. [Google Scholar] [CrossRef] [Green Version]
- Taverna, S.; Rigogliuso, S.; Salamone, M.; Vittorelli, M.L. Intracellular trafficking of endogenous fibroblast growth factor-2. FEBS J. 2008, 275, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Ghersi, G.; Zhao, Q.; Salamone, M.; Yeh, Y.; Zucker, S.; Chen, W.T. The protease complex consisting of dipeptidyl peptidase IV and seprase plays a role in the migration and invasion of human endothelial cells in collagenous matrices. Cancer Res. 2006, 66, 4652–4661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saladino, S.; Salamone, M.; Ghersi, G. MDA-MB-231 and 8701BC breast cancer lines promote the migration and invasiveness of ECV304 cells on 2D and 3D type-I collagen matrix. Cell Biol. Int. 2017, 41, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Später, T.; Menger, M.M.; Nickels, R.M.; Menger, M.D.; Laschke, M.W. Macrophages promote network formation and maturation of transplanted adipose tissue-derived microvascular fragments. J. Tissue Eng. 2020, 11, 2041731420911816. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, T.; Mukouyama, Y.S. Tissue Specific Origin, Development, and Pathological Perspectives of Pericytes. Front. Cardiovasc. Med. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurz, H.; Fehr, J.; Nitschke, R.; Burkhardt, H. Pericytes in the mature chorioallantoic membrane capillary plexus contain desmin and α-smooth muscle actin: Relevance for non-sprouting angiogenesis. Histochem. Cell Biol. 2008, 130, 1027–1040. [Google Scholar] [CrossRef]
- Nyberg, P.; Xie, L.; Kalluri, R. Endogenous inhibitors of angiogenesis. Cancer Res. 2005, 65, 3967–3979. [Google Scholar] [CrossRef] [Green Version]
- Folkman, J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat. Med. 1995, 1, 27–30. [Google Scholar] [CrossRef]
- Marech, I.; Leporini, C.; Ammendola, M.; Porcelli, M.; Gadaleta, C.D.; Russo, E.; de Sarro, G.; Ranieri, G. Classical and non-classical proangiogenic factors as a target of antiangiogenic therapy in tumor microenvironment. Cancer Lett. 2016, 380, 216–226. [Google Scholar] [CrossRef]
- Tsai, J.C.; Goldman, C.K.; Gillespie, G.Y. Vascular endothelial growth factor in human glioma cell lines: Induced secretion by EGF, PDGF-BB, and bFGF. J. Neurosurg. 1995, 82, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Hu, B.; Gu, W.; Xu, L.; Wang, D.; Huang, H.J.S.; Cavenee, W.K.; Cheng, S.Y. Platelet-derived growth factor-B enhances glioma angiogenesis by stimulating vascular endothelial growth factor expression in tumor endothelia and by promoting pericyte recruitment. Am. J. Pathol. 2003, 162, 1083–1093. [Google Scholar] [CrossRef] [Green Version]
- Seghezzi, G.; Patel, S.; Ren, C.J.; Gualandris, A.; Pintucci, G.; Robbins, E.S.; Shapiro, R.L.; Galloway, A.C.; Rifkin, D.B.; Mignatti, P. Fibroblast growth factor-2 (FGF-2) induces vascular endothelial growth factor (VEGF) expression in the endothelial cells of forming capillaries: An autocrine mechanism contributing to angiogenesis. J. Cell Biol. 1998, 141, 1659–1673. [Google Scholar] [CrossRef]
- Hamano, Y.; Zeisberg, M.; Sugimoto, H.; Lively, J.C.; Maeshima, Y.; Yang, C.; Hynes, R.O.; Werb, Z.; Sudhakar, A.; Kalluri, R. Physiological levels of tumstatin, a fragment of collagen IV α3 chain, are generated by MMP-9 proteolysis and suppress angiogenesis via αVβ3 integrin. Cancer Cell. 2003, 3, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Duda, D.G.; Fukumura, D.; Jain, R.K. Role of eNOS in neovascularization: NO for endothelial progenitor cells. Trends Mol. Med. 2004, 10, 143–145. [Google Scholar] [CrossRef]
- Kroll, J.; Waltenberger, J. VEGF-A induces expression of eNOS and iNOS in endothelial cells via VEGF receptor-2 (KDR). Biochem. Biophys. Res. Commun. 1998, 252, 743–746. [Google Scholar] [CrossRef]
- Liu, Z.; Lebrin, F.; Maring, J.A.; van den Driesche, S.; van der Brink, S.; van Dinther, M.; Thorikay, M.; Martin, S.; Kobayashi, K.; Hawinkels, L.J.A.C.; et al. ENDOGLIN Is Dispensable for Vasculogenesis, but Required for Vascular Endothelial Growth Factor-Induced Angiogenesis. PLoS ONE 2014, 9, e86273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ader, I.; Brizuela, L.; Bouquerel, P.; Malavaud, B.; Cuvillier, O. Sphingosine kinase 1: A new modulator of hypoxia inducible factor 1α during hypoxia in human cancer cells. Cancer Res. 2008, 68, 8635–8642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anelli, V.; Gault, C.R.; Cheng, A.B.; Obeid, L.M. Sphingosine kinase 1 is up-regulated during hypoxia in U87MG glioma cells: Role of hypoxia-inducible factors 1 and 2. J. Biol. Chem. 2008, 283, 3365–3375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalbach, L.; Roma, L.P.; Schmitt, B.M.; Becker, V.; Körbel, C.; Wrublewsky, S.; Pack, M.; Später, T.; Metzger, W.; Menger, M.M.; et al. Improvement of islet transplantation by the fusion of islet cells with functional blood vessels. EMBO Mol. Med. 2021, 13, e12616. [Google Scholar] [CrossRef]
- de Souza, B.M.; Bouças, A.P.; de Oliveira, F.d.S.; Reis, K.P.; Ziegelmann, P.; Bauer, A.C.; Crispim, D. Effect of co-culture of mesenchymal stem/stromal cells with pancreatic islets on viability and function outcomes: A systematic review and meta-analysis. Islets 2017, 9, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Aamodt, K.I.; Powers, A.C. Signals in the pancreatic islet microenvironment influence β-cell proliferation. Diabetes Obes. Metab. 2017, 19, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Cohrs, C.M.; Chen, C.; Jahn, S.R.; Stertmann, J.; Chmelova, H.; Weitz, J.; Bähr, A.; Klymiuk, N.; Steffen, A.; Ludwig, B.; et al. Vessel network architecture of adult human islets promotes distinct cell-cell interactions in situ and is altered after transplantation. Endocrinology 2017, 158, 1373–1385. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences (5′–3′) | Accession Number |
|---|---|---|
| Gapdh | CAGCCTCGTCTCATAGACAAGATG a AAGGCAGCCCTGGTAACCA b | AF106860 |
| bFgf | GAGAGAGGAGTTGTGTCCATCAAG a GCAGCCGTCCATCTTCCTT b | X61697 |
| Pdgfb | TGGAGTCGAGTCGGAAAGCT a GAAGTTGGCATTGGTGCGAT b | NM_031524.1 |
| Vegfb | GAGGAAAGGGAAAGGGTCAAAA a CACAGTGAACGCTCCAGGATT b | AF062644 |
| Hif1a | GTTTACTAAAGGACAAGTCACC a TTCTGTTTGTTGAAGGGAG b | NM024359 |
| Nos3 | GACCCTCACCGATACAACATAC a CATACAGGATAGTCGCCTTCAC b | NM_021838 |
| SphK1 | TCAGTCTGTCCTGGGGTTTC a TCCTCCAGAGGAACGAGGTA b | NM_001270811.1 |
| Col4a3 | CCCTTGAGCCCTACGTTAGCA a CCTCAGAGCCTGCACTTGTAAACA b | XM_343607 |
| Eng | TGCTCCCTCTGGTCATTACC a CCTGGCTGGTGGTGTATGTC b | NM_001010968 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salamone, M.; Rigogliuso, S.; Nicosia, A.; Campora, S.; Bruno, C.M.; Ghersi, G. 3D Collagen Hydrogel Promotes In Vitro Langerhans Islets Vascularization through ad-MVFs Angiogenic Activity. Biomedicines 2021, 9, 739. https://doi.org/10.3390/biomedicines9070739
Salamone M, Rigogliuso S, Nicosia A, Campora S, Bruno CM, Ghersi G. 3D Collagen Hydrogel Promotes In Vitro Langerhans Islets Vascularization through ad-MVFs Angiogenic Activity. Biomedicines. 2021; 9(7):739. https://doi.org/10.3390/biomedicines9070739
Chicago/Turabian StyleSalamone, Monica, Salvatrice Rigogliuso, Aldo Nicosia, Simona Campora, Carmelo Marco Bruno, and Giulio Ghersi. 2021. "3D Collagen Hydrogel Promotes In Vitro Langerhans Islets Vascularization through ad-MVFs Angiogenic Activity" Biomedicines 9, no. 7: 739. https://doi.org/10.3390/biomedicines9070739
APA StyleSalamone, M., Rigogliuso, S., Nicosia, A., Campora, S., Bruno, C. M., & Ghersi, G. (2021). 3D Collagen Hydrogel Promotes In Vitro Langerhans Islets Vascularization through ad-MVFs Angiogenic Activity. Biomedicines, 9(7), 739. https://doi.org/10.3390/biomedicines9070739