
Tcf4 Is Involved in Subset Specification of Mesodiencephalic Dopaminergic Neurons



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Experimental Section
2.1. Ethics Statement
2.2. Animals
2.3. Genotyping
2.4. In Situ Hybridization and Combined TH-DAB IHC
2.5. Immunohistochemistry
2.6. Quantitative PCR (qPCR)
3. Results
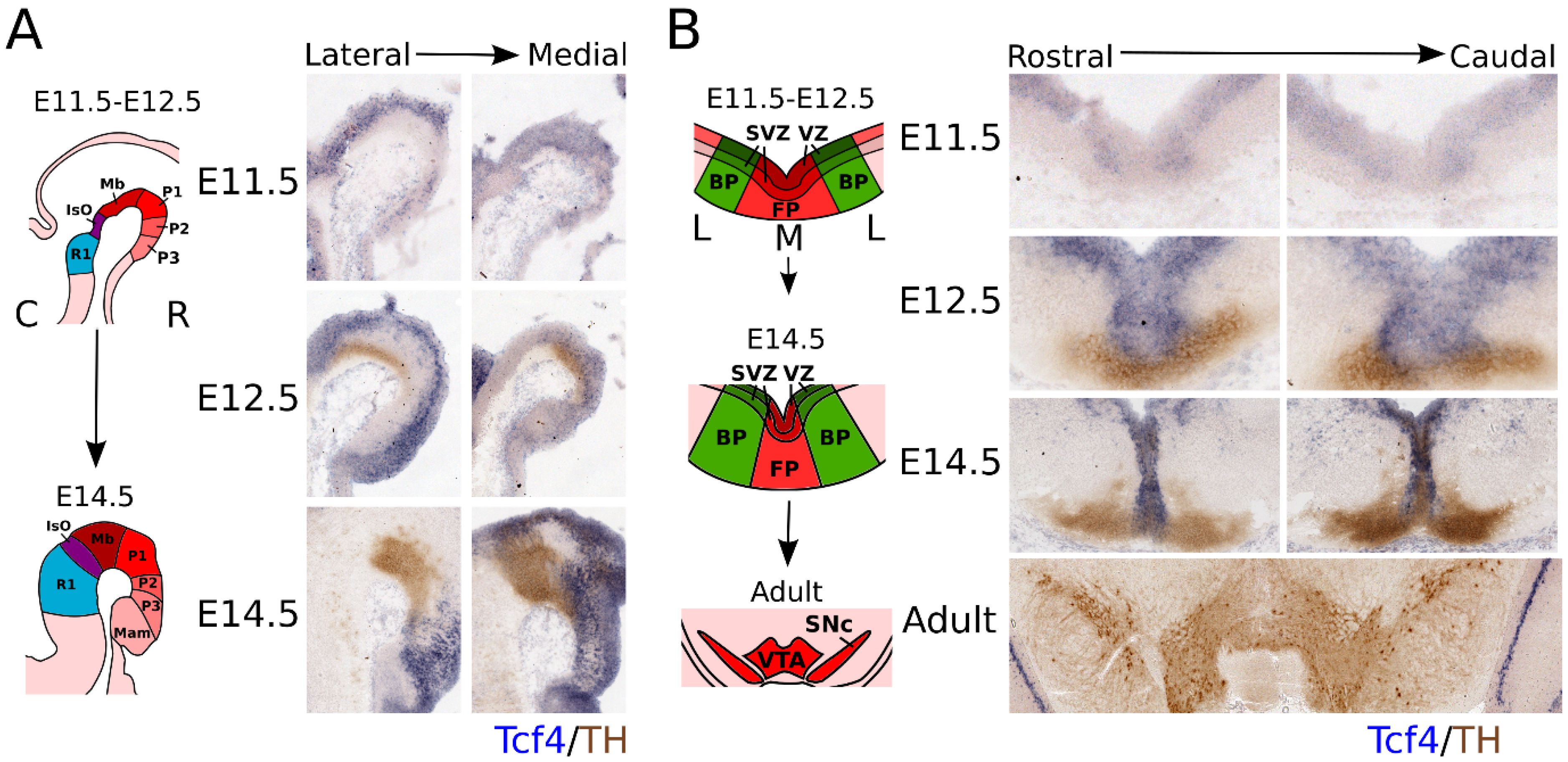
3.1. Tcf4 Is Expressed from E11.5 Onward and Partly Overlaps with the TH-Expressing Population
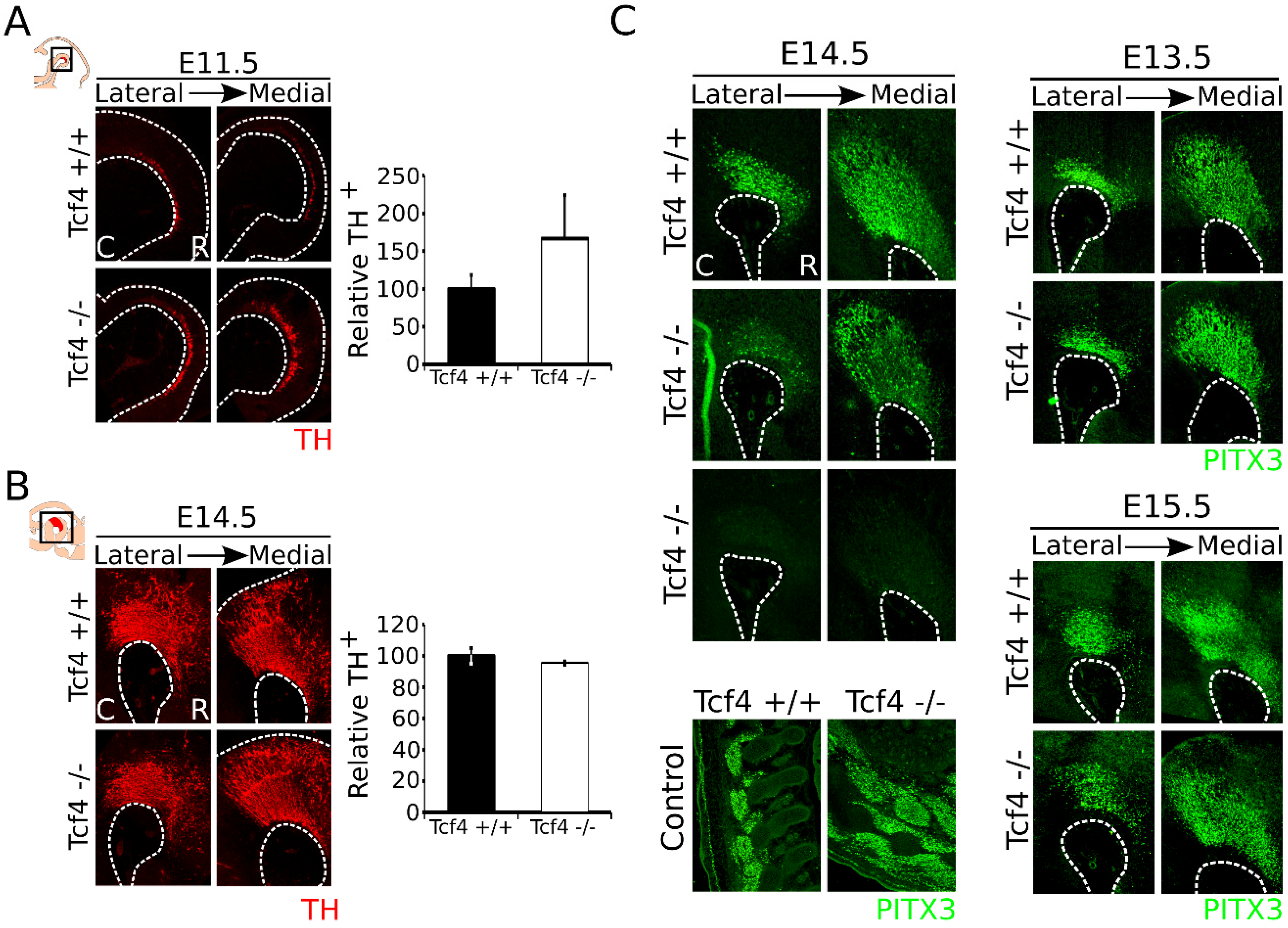
3.2. Loss of Tcf4 Leads to an Upward Trend in TH-Expressing Neurons at E11.5 but Not at E14.5, and Sporadic Loss of PITX3 at E14.5
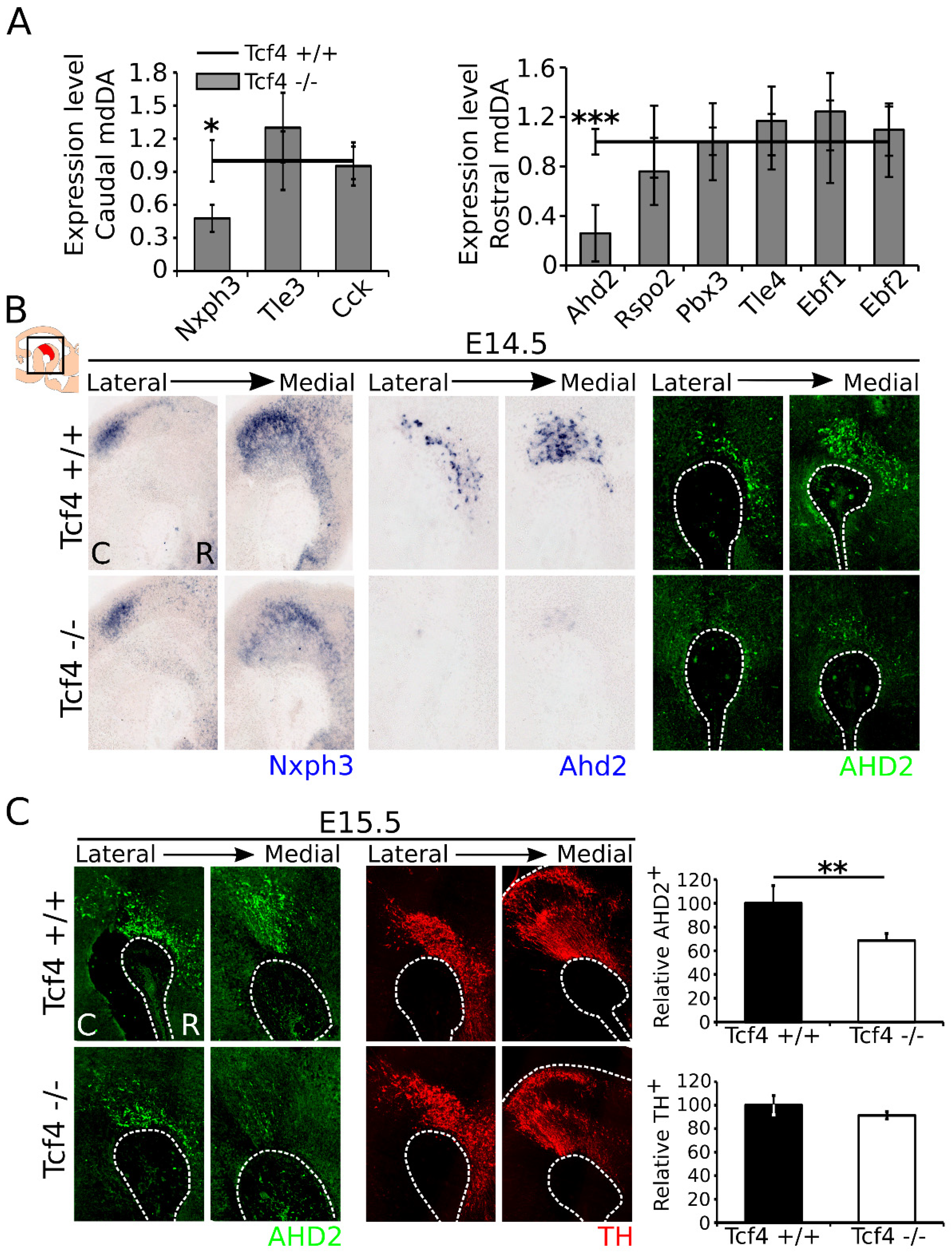
3.3. Correct mdDA Neuronal Subset Specification Is Affected in the Tcf4 Mutant
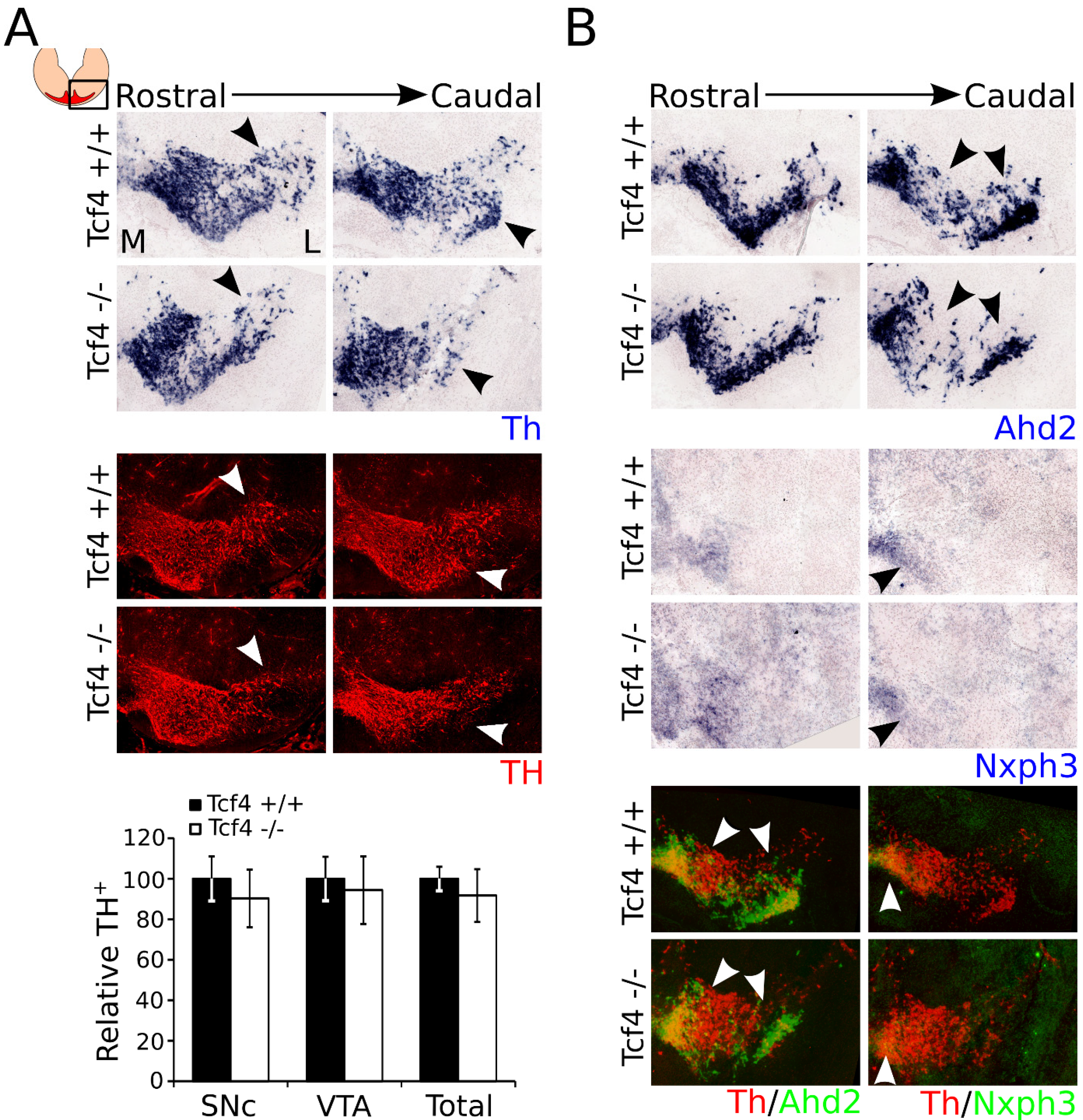
3.4. Ahd2 and Nxph3 Expression Partially Recovers Later in Development, Whereas Th Expression Becomes Slightly Affected at P0
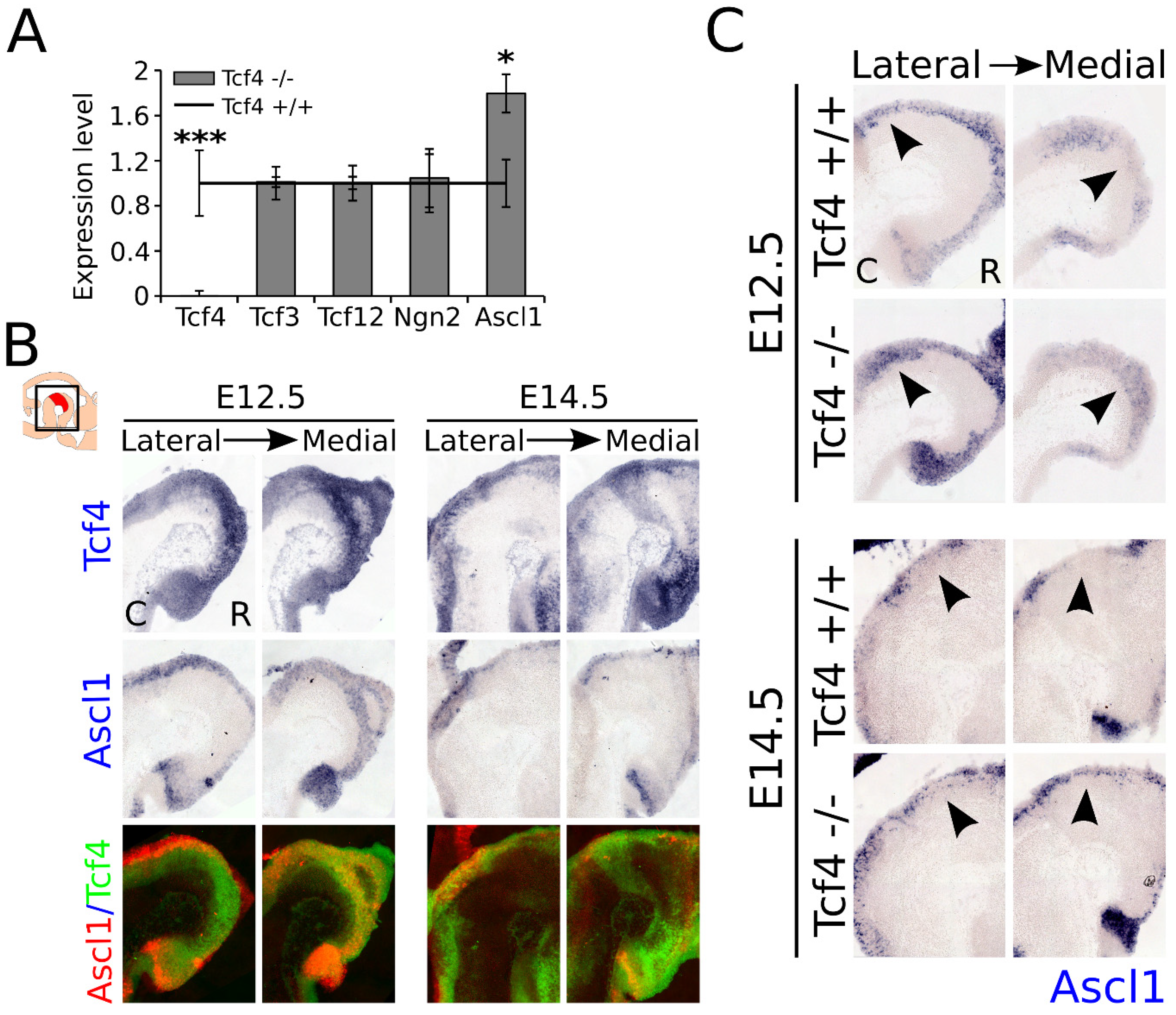
3.5. Ascl1 Is Upregulated in the Developing Midbrain of the Tcf4 Mutant
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bayer, S.A.; Wills, K.V.; Triarhou, L.C.; Ghetti, B. Time of neuron origin and gradients of neurogenesis in midbrain dopaminergic neurons in the mouse. Exp. Brain Res. Exp. Hirnforsch. Exp. Céréb. 1995, 105, 191–199. [Google Scholar] [CrossRef]
- Mesman, S.; von Oerthel, L.; Smidt, M.P. Mesodiencephalic dopaminergic neuronal differentiation does not involve GLI2A-mediated SHH-signaling and is under the direct influence of canonical WNT signaling. PLoS ONE 2014, 9, e97926. [Google Scholar] [CrossRef]
- Ono, Y.; Nakatani, T.; Sakamoto, Y.; Mizuhara, E.; Minaki, Y.; Kumai, M.; Hamaguchi, A.; Nishimura, M.; Inoue, Y.; Hayashi, H.; et al. Differences in neurogenic potential in floor plate cells along an anteroposterior location: Midbrain dopaminergic neurons originate from mesencephalic floor plate cells. Dev. Camb. Engl. 2007, 134, 3213–3225. [Google Scholar] [CrossRef]
- Di Salvio, M.; Di Giovannantonio, L.G.; Acampora, D.; Prosperi, R.; Omodei, D.; Prakash, N.; Wurst, W.; Simeone, A. Otx2 controls neuron subtype identity in ventral tegmental area and antagonizes vulnerability to MPTP. Nat. Neurosci. 2010, 13, 1481–1488. [Google Scholar] [CrossRef]
- Hoekstra, E.J.; Mesman, S.; de Munnik, W.A.; Smidt, M.P. LMX1B is part of a transcriptional complex with PSPC1 and PSF. PLoS ONE 2013, 8, e53122. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, F.M.J.; Veenvliet, J.V.; Almirza, W.H.; Hoekstra, E.J.; von Oerthel, L.; van der Linden, A.J.A.; Neijts, R.; Koerkamp, M.G.; van Leenen, D.; Holstege, F.C.P.; et al. Retinoic acid-dependent and -independent gene-regulatory pathways of Pitx3 in meso-diencephalic dopaminergic neurons. Dev. Camb. Engl. 2011, 138, 5213–5222. [Google Scholar] [CrossRef] [PubMed]
- Smits, S.M.; von Oerthel, L.; Hoekstra, E.J.; Burbach, J.P.H.; Smidt, M.P. Molecular Marker Differences Relate to Developmental Position and Subsets of Mesodiencephalic Dopaminergic Neurons. PLoS ONE 2013, 8, e76037. [Google Scholar] [CrossRef] [PubMed]
- Veenvliet, J.V.; Dos Santos, M.T.M.A.; Kouwenhoven, W.M.; von Oerthel, L.; Lim, J.L.; van der Linden, A.J.A.; Koerkamp, M.J.A.G.; Holstege, F.C.P.; Smidt, M.P. Specification of dopaminergic subsets involves interplay of En1 and Pitx3. Dev. Camb. Engl. 2013, 140, 3373–3384. [Google Scholar] [CrossRef]
- Barzilai, A.; Melamed, E. Molecular mechanisms of selective dopaminergic neuronal death in Parkinson’s disease. Trends Mol. Med. 2003, 9, 126–132. [Google Scholar] [CrossRef]
- Mesman, S.; Smidt, M.P. Acquisition of the Midbrain Dopaminergic Neuronal Identity. Int. J. Mol. Sci. 2020, 21, 4638. [Google Scholar] [CrossRef]
- Skinner, M.K.; Rawls, A.; Wilson-Rawls, J.; Roalson, E.H. Basic helix-loop-helix transcription factor gene family phylogenetics and nomenclature. Differ. Res. Biol. Divers. 2010, 80, 1–8. [Google Scholar] [CrossRef]
- Murre, C.; McCaw, P.S.; Vaessin, H.; Caudy, M.; Jan, L.Y.; Jan, Y.N.; Cabrera, C.V.; Buskin, J.N.; Hauschka, S.D.; Lassar, A.B. Interactions between heterologous helix-loop-helix proteins generate complexes that bind specifically to a common DNA sequence. Cell 1989, 58, 537–544. [Google Scholar] [CrossRef]
- Powell, L.M.; Jarman, A.P. Context dependence of proneural bHLH proteins. Curr. Opin. Genet. Dev. 2008, 18, 411–417. [Google Scholar] [CrossRef]
- Andersson, E.; Jensen, J.B.; Parmar, M.; Guillemot, F.; Björklund, A. Development of the mesencephalic dopaminergic neuron system is compromised in the absence of neurogenin 2. Dev. Camb. Engl. 2006, 133, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Kele, J.; Simplicio, N.; Ferri, A.L.M.; Mira, H.; Guillemot, F.; Arenas, E.; Ang, S.-L. Neurogenin 2 is required for the development of ventral midbrain dopaminergic neurons. Dev. Camb. Engl. 2006, 133, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Mesman, S.; Smidt, M.P. Tcf12 Is Involved in Early Cell-Fate Determination and Subset Specification of Midbrain Dopamine Neurons. Front. Mol. Neurosci. 2017, 10, 353. [Google Scholar] [CrossRef]
- Peterson, D.J.; Marckini, D.N.; Straight, J.L.; King, E.M.; Johnson, W.; Sarah, S.S.; Chowdhary, P.K.; DeLano-Taylor, M.K. The Basic Helix-Loop-Helix Gene Nato3 Drives Expression of Dopaminergic Neuron Transcription Factors in Neural Progenitors. Neuroscience 2019, 421, 176–191. [Google Scholar] [CrossRef]
- Massari, M.E.; Murre, C. Helix-loop-helix proteins: Regulators of transcription in eucaryotic organisms. Mol. Cell. Biol. 2000, 20, 429–440. [Google Scholar] [CrossRef]
- Wojciechowski, J.; Lai, A.; Kondo, M.; Zhuang, Y. E2A and HEB are required to block thymocyte proliferation prior to pre-TCR expression. J. Immunol. Baltim. Md 1950 2007, 178, 5717–5726. [Google Scholar] [CrossRef]
- Wikström, I.; Forssell, J.; Penha-Goncalves, M.N.; Bergqvist, I.; Holmberg, D. A role for E2-2 at the DN3 stage of early thymopoiesis. Mol. Immunol. 2008, 45, 3302–3311. [Google Scholar] [CrossRef]
- Fischer, B.; Azim, K.; Hurtado-Chong, A.; Ramelli, S.; Fernández, M.; Raineteau, O. E-proteins orchestrate the progression of neural stem cell differentiation in the postnatal forebrain. Neural Develop. 2014, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Ravanpay, A.C.; Olson, J.M. E protein dosage influences brain development more than family member identity. J. Neurosci. Res. 2008, 86, 1472–1481. [Google Scholar] [CrossRef] [PubMed]
- Sweatt, J.D. Pitt-Hopkins Syndrome: Intellectual disability due to loss of TCF4-regulated gene transcription. Exp. Mol. Med. 2013, 45, e21. [Google Scholar] [CrossRef] [PubMed]
- Zollino, M.; Zweier, C.; Van Balkom, I.D.; Sweetser, D.A.; Alaimo, J.; Bijlsma, E.K.; Cody, J.; Elsea, S.H.; Giurgea, I.; Macchiaiolo, M.; et al. Diagnosis and management in Pitt-Hopkins syndrome: First international consensus statement. Clin. Genet. 2019, 95, 462–478. [Google Scholar] [CrossRef]
- Flora, A.; Garcia, J.J.; Thaller, C.; Zoghbi, H.Y. The E-protein Tcf4 interacts with Math1 to regulate differentiation of a specific subset of neuronal progenitors. Proc. Natl. Acad. Sci. USA 2007, 104, 15382–15387. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Y.; Morozov, Y.M.; Chen, X.; Page, S.C.; Rannals, M.D.; Maher, B.J.; Rakic, P. Disruption of TCF4 regulatory networks leads to abnormal cortical development and mental disabilities. Mol. Psychiatry 2019, 24, 1235–1246. [Google Scholar] [CrossRef]
- Mesman, S.; Bakker, R.; Smidt, M.P. Tcf4 is required for correct brain development during embryogenesis. Mol. Cell. Neurosci. 2020, 106, 103502. [Google Scholar] [CrossRef]
- Yoon, S.O.; Chikaraishi, D.M. Isolation of two E-box binding factors that interact with the rat tyrosine hydroxylase enhancer. J. Biol. Chem. 1994, 269, 18453–18462. [Google Scholar] [CrossRef]
- Smidt, M.P.; Smits, S.M.; Bouwmeester, H.; Hamers, F.P.T.; van der Linden, A.J.A.; Hellemons, A.J.C.G.M.; Graw, J.; Burbach, J.P.H. Early developmental failure of substantia nigra dopamine neurons in mice lacking the homeodomain gene Pitx3. Dev. Camb. Engl. 2004, 131, 1145–1155. [Google Scholar] [CrossRef]
- Fenstermaker, A.G.; Prasad, A.A.; Bechara, A.; Adolfs, Y.; Tissir, F.; Goffinet, A.; Zou, Y.; Pasterkamp, R.J. Wnt/planar cell polarity signaling controls the anterior-posterior organization of monoaminergic axons in the brainstem. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 16053–16064. [Google Scholar] [CrossRef]
- Kolk, S.M.; Gunput, R.-A.F.; Tran, T.S.; van den Heuvel, D.M.A.; Prasad, A.A.; Hellemons, A.J.C.G.M.; Adolfs, Y.; Ginty, D.D.; Kolodkin, A.L.; Burbach, J.P.H.; et al. Semaphorin 3F is a bifunctional guidance cue for dopaminergic axons and controls their fasciculation, channeling, rostral growth, and intracortical targeting. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 12542–12557. [Google Scholar] [CrossRef] [PubMed]
- Smidt, M.P.; Asbreuk, C.H.J.; Cox, J.J.; Chen, H.; Johnson, R.L.; Burbach, J.P.H. A second independent pathway for development of mesencephalic dopaminergic neurons requires Lmx1b. Nat. Neurosci. 2000, 3, 337–341. [Google Scholar] [CrossRef]
- Bye, C.R.; Thompson, L.H.; Parish, C.L. Birth dating of midbrain dopamine neurons identifies A9 enriched tissue for transplantation into Parkinsonian mice. Exp. Neurol. 2012, 236, 58–68. [Google Scholar] [CrossRef]
- Zhuang, Y.; Cheng, P.; Weintraub, H. B-lymphocyte development is regulated by the combined dosage of three basic helix-loop-helix genes, E2A, E2-2, and HEB. Mol. Cell. Biol. 1996, 16, 2898–2905. [Google Scholar] [CrossRef] [PubMed]
- Tiklová, K.; Björklund, Å.K.; Lahti, L.; Fiorenzano, A.; Nolbrant, S.; Gillberg, L.; Volakakis, N.; Yokota, C.; Hilscher, M.M.; Hauling, T.; et al. Single-Cell RNA Sequencing Reveals Midbrain Dopamine Neuron Diversity Emerging during Mouse Brain Development. Nat. Commun. 2019, 10, 581. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, E.J.; von Oerthel, L.; van der Heide, L.P.; Kouwenhoven, W.M.; Veenvliet, J.V.; Wever, I.; Jin, Y.-R.; Yoon, J.K.; van der Linden, A.J.A.; Holstege, F.C.P.; et al. Lmx1a Encodes a Rostral Set of Mesodiencephalic Dopaminergic Neurons Marked by the Wnt/B-Catenin Signaling Activator R-spondin 2. PLoS ONE 2013, 8, e74049. [Google Scholar] [CrossRef]
- Nishimura, K.; Murayama, S.; Takahashi, J. Identification of Neurexophilin 3 as a Novel Supportive Factor for Survival of Induced Pluripotent Stem Cell-Derived Dopaminergic Progenitors. Stem Cells Transl. Med. 2015, 4, 932–944. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesman, S.; Wever, I.; Smidt, M.P. Tcf4 Is Involved in Subset Specification of Mesodiencephalic Dopaminergic Neurons. Biomedicines 2021, 9, 317. https://doi.org/10.3390/biomedicines9030317
Mesman S, Wever I, Smidt MP. Tcf4 Is Involved in Subset Specification of Mesodiencephalic Dopaminergic Neurons. Biomedicines. 2021; 9(3):317. https://doi.org/10.3390/biomedicines9030317
Chicago/Turabian StyleMesman, Simone, Iris Wever, and Marten P. Smidt. 2021. "Tcf4 Is Involved in Subset Specification of Mesodiencephalic Dopaminergic Neurons" Biomedicines 9, no. 3: 317. https://doi.org/10.3390/biomedicines9030317
APA StyleMesman, S., Wever, I., & Smidt, M. P. (2021). Tcf4 Is Involved in Subset Specification of Mesodiencephalic Dopaminergic Neurons. Biomedicines, 9(3), 317. https://doi.org/10.3390/biomedicines9030317