
Red Blood Cell-Derived Iron Alters Macrophage Function in COPD

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Lung Tissue Preparation
2.3. Lung Macrophage Culture
2.4. Monocyte-Derived Macrophage Culture
2.5. Statistics
3. Results
3.1. Study Subjects
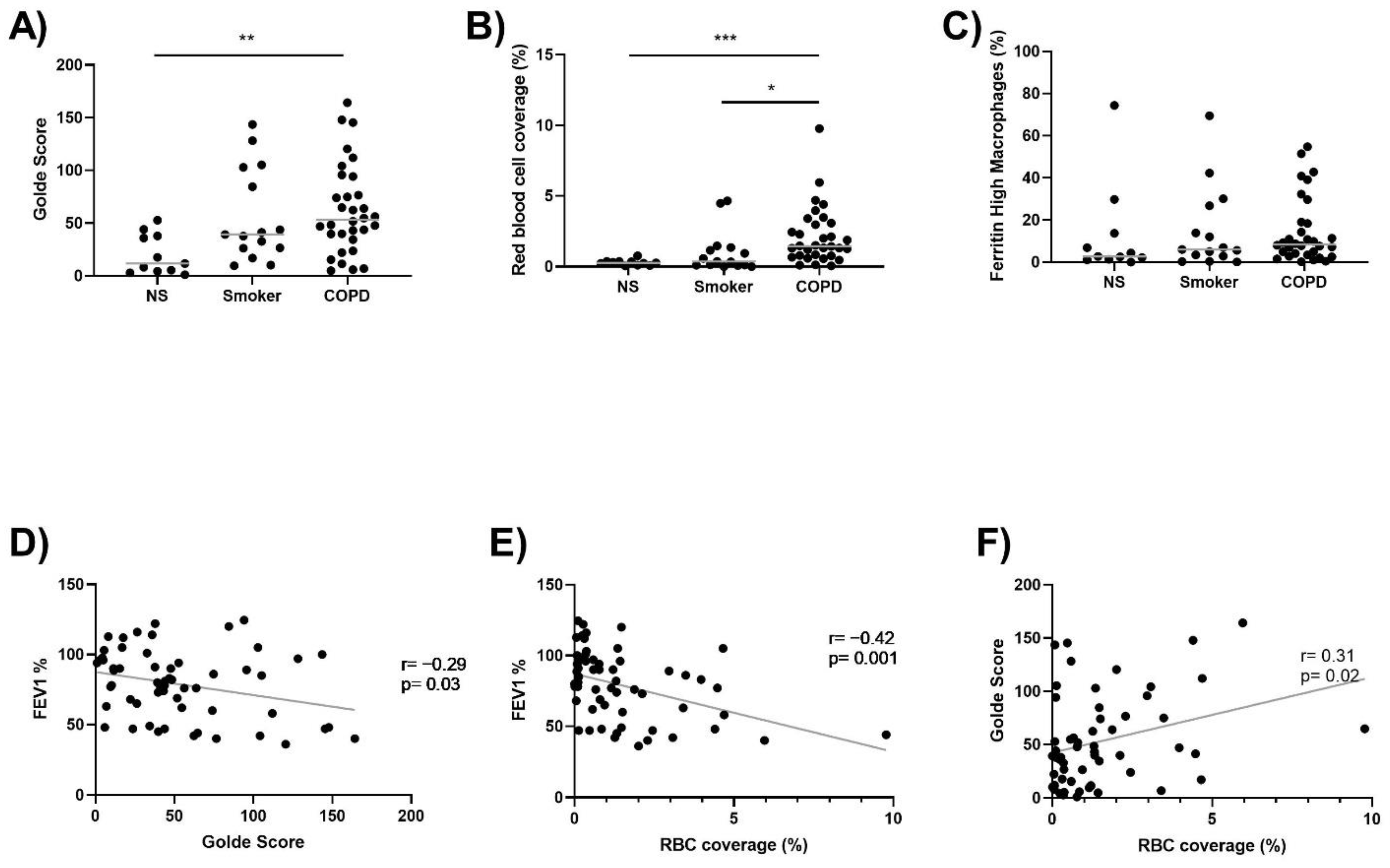
3.2. Lung Macrophage Iron and Ferritin Staining, and Alveolar RBC Coverage
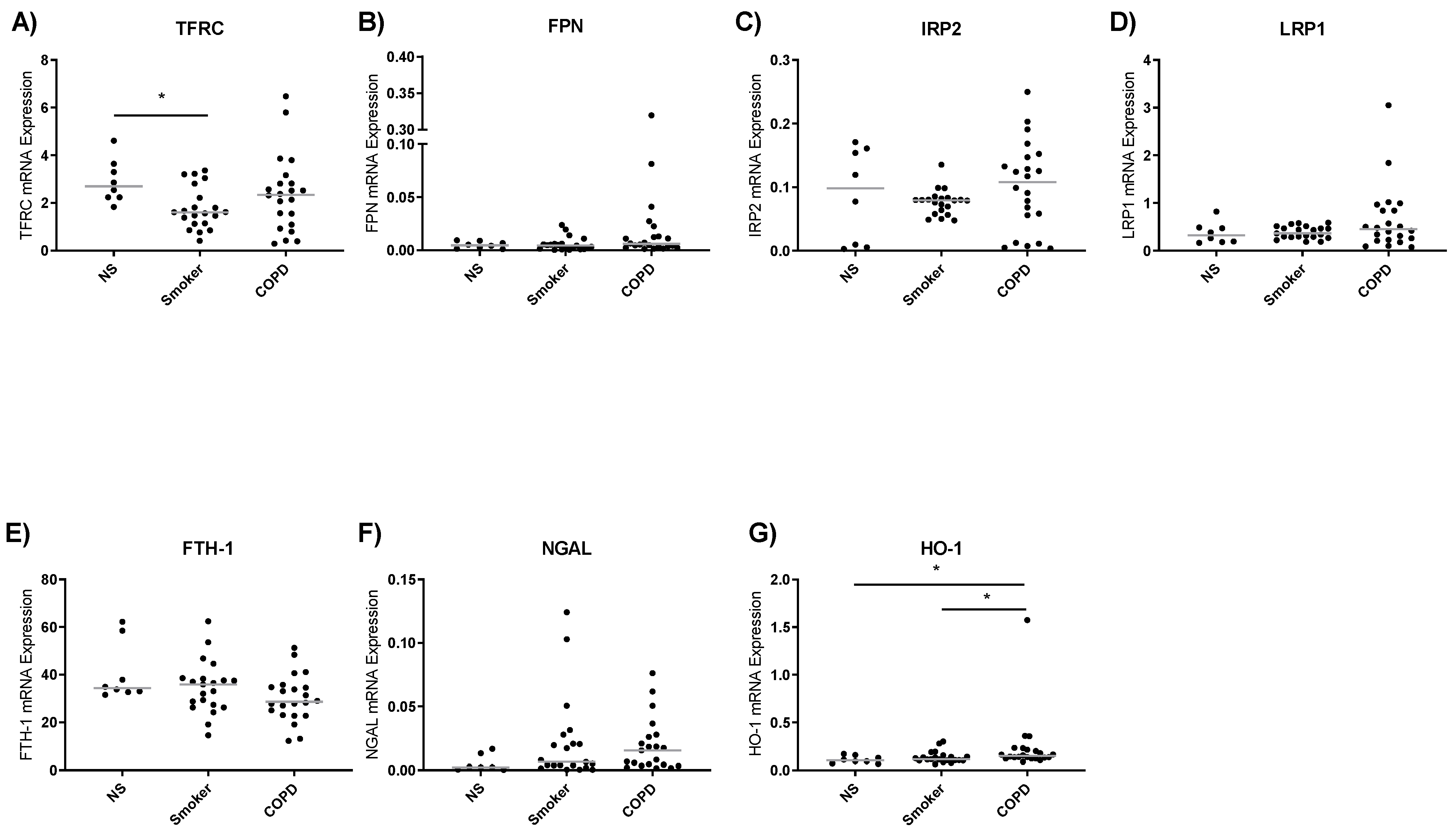
3.3. Iron Metabolism Gene Expression in Lung Macrophages
3.4. Effect of Iron Treatment on Lung Macrophage Gene Expression
3.4.1. Iron Metabolism Gene Expression
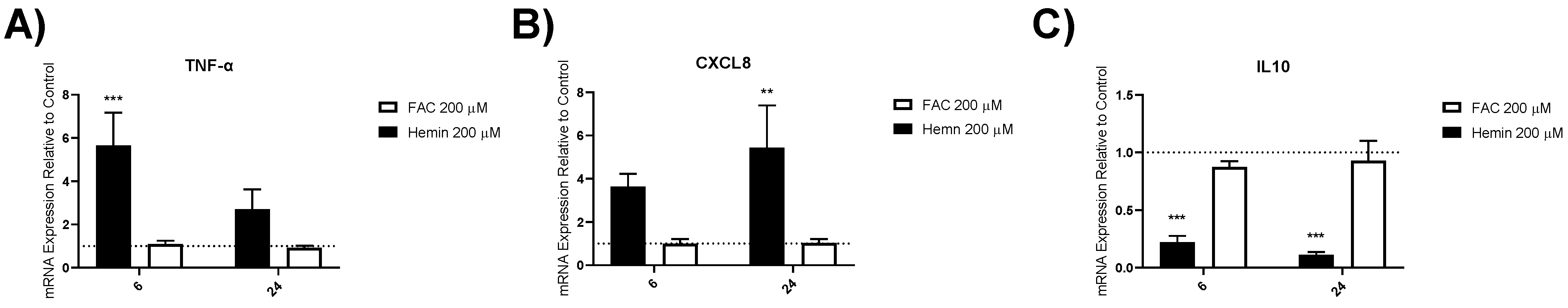
3.4.2. Inflammatory Gene Expression
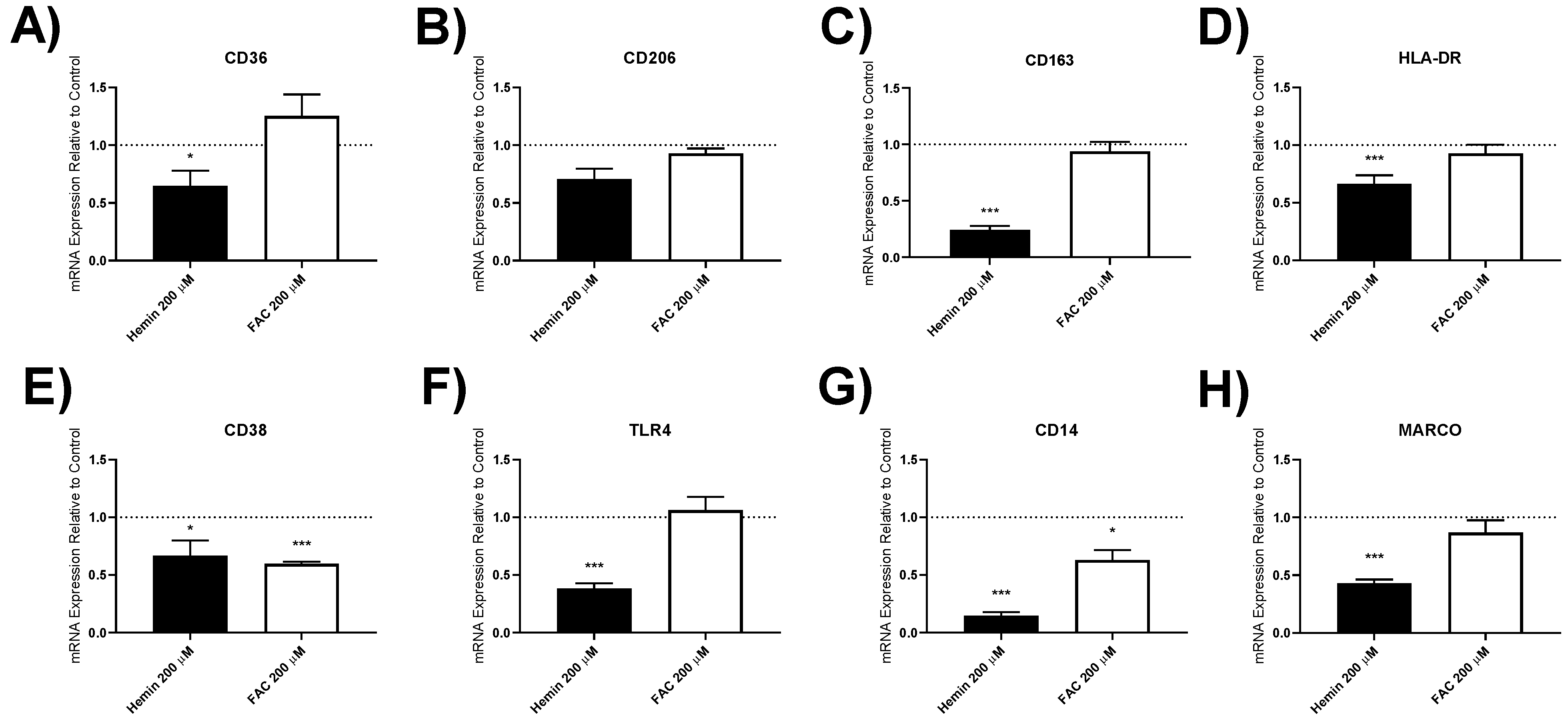
3.4.3. Macrophage Polarisation Marker Gene Expression
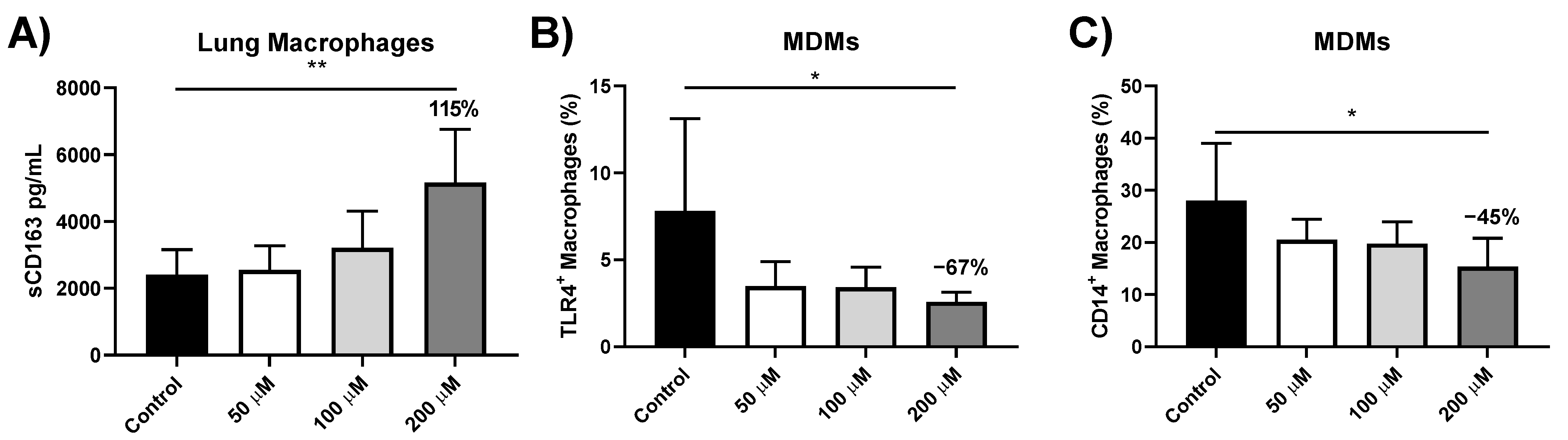
3.5. sCD163 Release from Iron-Treated Lung Macrophages
3.6. Effect of Iron Treatment on TLR4 and CD14 Protein Expression in MDMs
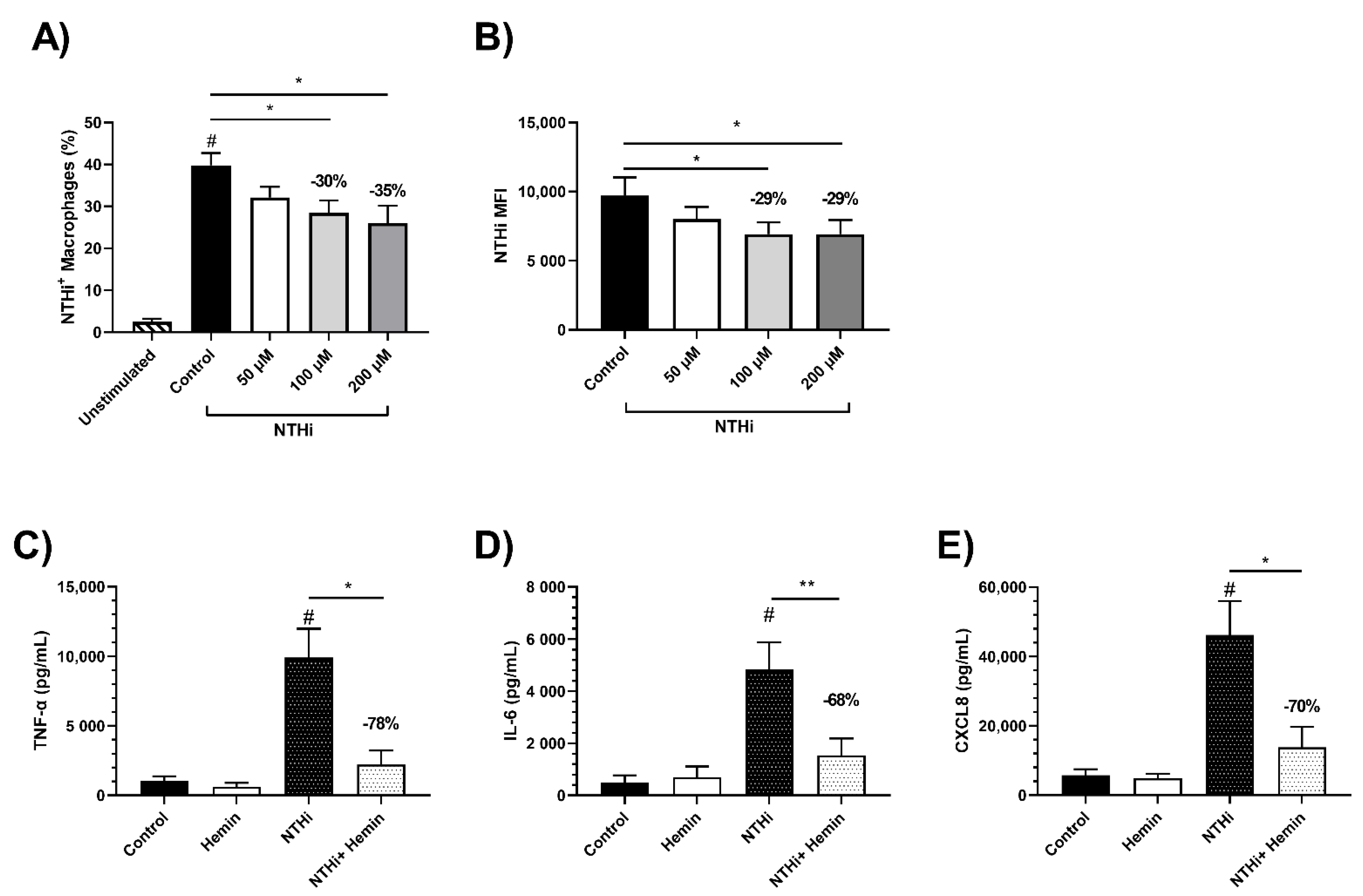
3.7. Effects of Iron Treatment on MDM Phagocytosis of NTHi
3.8. Effect of Iron Treatment on MDM Cytokine Response to NTHi
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vogelmeier, C.F.; Criner, G.J.; Martinez, F.J.; Anzueto, A.; Barnes, P.J.; Bourbeau, J.; Celli, B.R.; Chen, R.; Decramer, M.; Fabbri, L.M.; et al. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease 2017 Report. GOLD Executive Summary. Am. J. Respir. Crit. Care Med. 2017, 195, 557–582. [Google Scholar] [CrossRef]
- Corhay, J.L.; Weber, G.; Bury, T.; Mariz, S.; Roelandts, I.; Radermecker, M.F. Iron content in human alveolar macrophages. Eu-Ropean Respir. J. 1992, 5, 804–809. [Google Scholar]
- E Nelson, M.; O’Brien-Ladner, A.R.; Wesselius, L.J. Regional variation in iron and iron-binding proteins within the lungs of smokers. Am. J. Respir. Crit. Care Med. 1996, 153, 1353–1358. [Google Scholar] [CrossRef]
- Ghio, A.J.; Hilborn, E.D.; Stonehuerner, J.G.; Dailey, L.A.; Carter, J.D.; Richards, J.H.; Crissman, K.M.; Foronjy, R.F.; Uyeminami, D.L.; Pinkerton, K.E. Particulate Matter in Cigarette Smoke Alters Iron Homeostasis to Produce a Biological Effect. Am. J. Respir. Crit. Care Med. 2008, 178, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Philippot, Q.; Deslee, G.; Adair-Kirk, T.L.; Woods, J.C.; Byers, D.; Conradi, S.; Dury, S.; Perotin, J.M.; Lebargy, F.; Cassanet, C.; et al. Increased iron sequestration in alveolar macrophages in chronic obstructive pulmonary disease. PLoS ONE 2014, 9, e96285. [Google Scholar] [CrossRef]
- Cloonan, S.M.; Mumby, S.; Adcock, I.M.; Choi, A.M.K.; Chung, K.F.; Quinlan, G.J. The “Iron”-y of Iron Overload and Iron Deficiency in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2017, 196, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, P.A.; Barnes, P.J. Oxidative Stress in COPD. Chest 2013, 144, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F.; Adcock, I. Multifaceted mechanisms in COPD: Inflammation, immunity, and tissue repair and destruction. Eur. Respir. J. 2008, 31, 1334–1356. [Google Scholar] [CrossRef]
- Bewley, M.A.; Budd, R.C.; Ryan, E.; Cole, J.; Collini, P.; Marshall, J.; Kolsum, U.; Beech, G.; Emes, R.D.; Tcherniaeva, I.; et al. Opsonic Phagocytosis in Chronic Obstructive Pulmonary Disease Is Enhanced by Nrf2 Agonists. Am. J. Respir. Crit. Care Med. 2018, 198, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Hodge, S.; Hodge, G.; Scicchitano, R.; Reynolds, P.N.; Holmes, M. Alveolar macrophages from subjects with chronic obstructive pulmonary disease are deficient in their ability to phagocytose apoptotic airway epithelial cells. Immunol. Cell Biol. 2003, 81, 289–296. [Google Scholar] [CrossRef]
- Lea, S.R.; Reynolds, S.L.; Kaur, M.; Simpson, K.D.; Hall, S.R.; Hessel, E.M.; Singh, D. The effects of repeated Toll-like receptors 2 and 4 stim-ulation in COPD alveolar macrophages. Int. J. Chronic Obstr. Pulm. Disease 2018, 13, 771–780. [Google Scholar] [CrossRef]
- Mesquita, G.; Silva, T.; Gomes, A.C.; Oliveira, P.F.; Alves, M.G.; Fernandes, R.; Almeida, A.A.; Moreira, A.C.; Gomes, M.S. H-Ferritin is essential for macrophages’ capacity to store or detoxify exogenously added iron. Sci. Rep. 2020, 10, 3061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Z.; Oromendia, C.; Kikkers, S.A.; Butler, J.J.; O’Beirne, S.; Kim, K.; O’Neal, W.K.; Freeman, C.M.; Christenson, S.A.; Peters, S.P.; et al. Increased airway iron parameters and risk for exac-erbation in COPD: An analysis from SPIROMICS. Sci. Rep. 2020, 10, 10562. [Google Scholar] [CrossRef] [PubMed]
- Cloonan, S.M.; Glass, K.; Laucho-Contreras, M.E.; Bhashyam, A.R.; Cervo, M.; Pabon, M.A.; Csaba, K.; Polverino, F.; Siempos, I.I.; Perezet, E.; et al. Mitochondrial iron chelation amelio-rates cigarette smoke-induced bronchitis and emphysema in mice. Nat. Med. 2016, 22, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Minagawa, S.; Araya, J.; Sakamoto, T.; Hara, H.; Tsubouchi, K.; Hosaka, Y.; Ichikawa, A.; Saito, N.; Kadota, T.; et al. Involvement of cigarette smoke-induced epithelial cell ferroptosis in COPD pathogenesis. Nat. Commun. 2019, 10, 3145. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.; Baker, J.R.; Di Giandomenico, S.; Kermani, P.; Parker, J.; Kim, K.; Yang, J.; Barnes, P.J.; Vaulont, S.; Scanduraet, J.M.; et al. Hepcidin Is Essential for Alveolar Macrophage Function and Is Disrupted by Smoke in a Murine Chronic Obstructive Pulmonary Disease Model. J. Immunol. 2020, 205, 2489–2498. [Google Scholar] [CrossRef]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef]
- Sakao, S.; Voelkel, N.F.; Tatsumi, K. The vascular bed in COPD: Pulmonary hypertension and pulmonary vascular alterations. Eur. Respir. Rev. 2014, 23, 350–355. [Google Scholar] [CrossRef]
- Müllebner, A.; Dorighello, G.G.; Kozlov, A.V.; Duvigneau, J.C. Interaction between Mitochondrial Reactive Oxygen Species, Heme Oxygenase, and Nitric Oxide Synthase Stimulates Phagocytosis in Macrophages. Front. Med. 2018, 4, 252. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Liu, Y.-K.; Hu, W.-T.; Tang, L.-L.; Sheng, Y.-R.; Wei, C.-Y.; Li, M.-Q.; Zhu, X.Y. Elevated heme impairs macrophage phagocytosis in endome-triosis. Reproduction 2019, 158, 257–266. [Google Scholar] [CrossRef]
- Martins, R.; Maier, J.; Gorki, A.-D.; Huber, K.V.M.; Sharif, O.; Starkl, P.; Saluzzo, S.; Quattrone, F.; Gawish, R.; Lakovits, K.; et al. Heme drives hemolysis-induced susceptibility to infection via disruption of phagocyte functions. Nat. Immunol. 2016, 17, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.; Sargent, C.; Singh, D. Glucocorticoid sensitivity of lipopolysaccharide-stimulated chronic obstructive pulmonary disease alveolar macrophages. Clin. Exp. Immunol. 2009, 158, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Golde, D.W.; Drew, W.L.; Klein, H.Z.; Finley, T.N.; Cline, M.J. Occult pulmonary haemorrhage in leukaemia. BMJ 1975, 2, 166–168. [Google Scholar] [CrossRef]
- Harvey, C.J.; Thimmulappa, R.K.; Sethi, S.; Kong, X.; Yarmus, L.; Brown, R.H.; Feller-Kopman, D.; Wise, R.; Biswal, S. Targeting Nrf2 Signaling Improves Bacterial Clearance by Alveolar Macrophages in Patients with COPD and in a Mouse Model. Sci. Transl. Med. 2011, 3, 32. [Google Scholar] [CrossRef]
- Wang, X.; Moser, C.; Louboutin, J.-P.; Lysenko, E.S.; Weiner, D.J.; Weiser, J.N.; Wilson, J.M. Toll-Like Receptor 4 Mediates Innate Immune Responses to Haemophilus influenzae Infection in Mouse Lung. J. Immunol. 2002, 168, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, R.; Fernandez, P.L.; Mourao-Sa, D.S.; Porto, B.N.; Dutra, F.F.; Alves, L.S.; Oliveira, M.F.; Oliveira, P.; Graça-Souza, A.V.; Bozza, M. Characterization of Heme as Activator of Toll-like Receptor 4. J. Biol. Chem. 2007, 282, 20221–20229. [Google Scholar] [CrossRef] [PubMed]
- Barker, B.L.; Haldar, K.; Patel, H.; Pavord, I.D.; Barer, M.R.; Brightling, C.E.; Bafathel, M. Association between pathogens detected using quan-titative polymerase chain reaction with airway inflammation in COPD at stable state and exacerbations. Chest 2015, 147, 46–55. [Google Scholar] [CrossRef]
- Dewhurst, J.A.; Lea, S.; Hardaker, E.; Dungwa, J.V.; Ravi, A.K.; Singh, D. Characterisation of lung macrophage subpopulations in COPD patients and controls. Sci. Rep. 2017, 7, 7143. [Google Scholar] [CrossRef]
- Mohan, S.; Ho, T.; Kjarsgaard, M.; Radford, K.; Borhan, A.S.M.; Thabane, L.; Nair, P. Hemosiderin in sputum macrophages may predict infective exacerbations of chronic obstructive pulmonary disease: A retrospective observational study. BMC Pulm. Med. 2017, 17, 60. [Google Scholar] [CrossRef]
- Chana, K.K.; Fenwick, P.S.; Nicholson, A.G.; Barnes, P.J.; Donnelly, L.E. Identification of a distinct glucocorticosteroid-insensitive pulmonary macrophage phenotype in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2014, 133, 207–216.e11. [Google Scholar] [CrossRef]
- Maras, J.S.; Das, S.; Sharma, S.; Sukriti, S.; Kumar, J.; Vyas, A.K.; Kumar, D.; Bhat, A.; Yadav, G.; Choudhary, M.C.; et al. Iron-Overload triggers ADAM-17 mediated inflammation in Severe Alcoholic Hepatitis. Sci. Rep. 2018, 8, 10264. [Google Scholar] [CrossRef] [PubMed]
- Higham, A.; Scott, T.; Li, J.; Gaskell, R.; Dikwa, A.B.; Shah, R.; Montero-Fernandez, M.; Lea, S.; Singh, D. Effects of corticosteroids on COPD lung macrophage phenotype and function. Clin. Sci. 2020, 134, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Kao, J.-K.; Wang, S.-C.; Ho, L.-W.; Huang, S.-W.; Chang, S.-H.; Yang, R.-C.; Ke, Y.-Y.; Wu, C.-Y.; Wang, J.-Y.; Shieh, J.-J. Chronic Iron Overload Results in Impaired Bacterial Killing of THP-1 Derived Macrophage through the Inhibition of Lysosomal Acidification. PLoS ONE 2016, 11, e0156713. [Google Scholar] [CrossRef] [PubMed]
- Rőszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [PubMed]
- Berenson, C.S.; Kruzel, R.L.; Eberhardt, E.; Dolnick, R.; Minderman, H.; Wallace, P.K.; Sethi, S. Impaired innate immune alveolar macro-phage response and the predilection for COPD exacerbations. Thorax 2014, 69, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Puxeddu, E.; Comandini, A.; Cavalli, F.; Pezzuto, G.; D’Ambrosio, C.; Senis, L.; Paci, M.; Curradi, G.; Sergiacomi, G.L.; Saltini, C. Iron laden macrophages in idiopathic pulmonary fibrosis: The telltale of occult alveolar hemorrhage? Pulm. Pharmacol. Ther. 2014, 28, 35–40. [Google Scholar] [CrossRef]
- Fukihara, J.; Taniguchi, H.; Ando, M.; Kondoh, Y.; Kimura, T.; Kataoka, K.; Furukawa, T.; Johkoh, T.; Fukuoka, J.; Sakamoto, K.; et al. Hemosiderin-laden macrophages are an independent factor correlated with pulmonary vascular resistance in idiopathic pulmonary fibrosis: A case control study. BMC Pulm. Med. 2017, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Maldonado, F.; Ryu, J.H.; Eiken, P.W.; E Hartman, T.; Bartholmai, B.J.; A Decker, P.; Yi, E.S. Iron deposition and increased alveolar septal capillary density in nonfibrotic lung tissue are associated with pulmonary hypertension in idiopathic pulmonary fibrosis. Respir. Res. 2010, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Arisi, I.; Puxeddu, E.; Mramba, L.K.; Amicosante, M.; Swaisgood, C.M.; Pallante, M.; Brantly, M.L.; Sköld, C.M.; Saltini, C. Bronchoalveolar lavage (BAL) cells in idiopathic pulmonary fibrosis express a complex pro-inflammatory, pro-repair, angiogenic activation pattern, likely associated with macrophage iron accumulation. PLoS ONE 2018, 13, e0194803. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.; Agusti, A.; Barberà, J.A.; Celli, B.; Criner, G.; Humbert, M.; Sin, D.D.; Voelkel, N.; Olschewski, H. Pulmonary Vascular Involvement in Chronic Obstructive Pulmonary Disease. Is There a Pulmonary Vascular Phenotype? Am. J. Respir. Crit. Care Med. 2018, 198, 1000–1011. [Google Scholar] [CrossRef]
- Belchamber, K.B.; Singh, R.; Batista, C.M.; Whyte, M.K.; Dockrell, D.H.; Kilty, I.; Robinson, M.J.; Wedzicha, J.; Barnes, P.J.; Donnelly, L.E. Defective bacterial phagocytosis is associated with dysfunctional mitochondria in COPD macrophages. Eur. Respir. J. 2019, 54, 1802244. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Smoker | Smoker | COPD | p Value | |
|---|---|---|---|---|
| n | 26 | 42 | 74 | N/A |
| Age (Years) | 62 (19) | 67 (10) | 69 (7) | >0.05 |
| Gender: Male (%) | 31 | 29 | 53 | <0.05 |
| FEV1 (L) | 2.7 (0.9) | 2.5 (1.0) | 1.7 (0.6) | <0.001 |
| FEV1% predicted | 108 (21) | 99 (18) | 66 (19) | <0.001 |
| FVC (L) | 3.3 (1.2) | 3.0 (0.8) | 3.1 (0.8) | >0.05 |
| FEV1/FVC ratio (%) | 81 (7) | 80 (10) | 54 (12) | <0.001 |
| Current smokers (%) | N/A | 50 | 47 | >0.05 |
| Pack years | N/A | 33 (18) | 50 (39) | <0.01 |
| ICS usage (%) | N/A | N/A | 25 | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baker, J.M.; Hammond, M.; Dungwa, J.; Shah, R.; Montero-Fernandez, A.; Higham, A.; Lea, S.; Singh, D. Red Blood Cell-Derived Iron Alters Macrophage Function in COPD. Biomedicines 2021, 9, 1939. https://doi.org/10.3390/biomedicines9121939
Baker JM, Hammond M, Dungwa J, Shah R, Montero-Fernandez A, Higham A, Lea S, Singh D. Red Blood Cell-Derived Iron Alters Macrophage Function in COPD. Biomedicines. 2021; 9(12):1939. https://doi.org/10.3390/biomedicines9121939
Chicago/Turabian StyleBaker, James M., Molly Hammond, Josiah Dungwa, Rajesh Shah, Angeles Montero-Fernandez, Andrew Higham, Simon Lea, and Dave Singh. 2021. "Red Blood Cell-Derived Iron Alters Macrophage Function in COPD" Biomedicines 9, no. 12: 1939. https://doi.org/10.3390/biomedicines9121939
APA StyleBaker, J. M., Hammond, M., Dungwa, J., Shah, R., Montero-Fernandez, A., Higham, A., Lea, S., & Singh, D. (2021). Red Blood Cell-Derived Iron Alters Macrophage Function in COPD. Biomedicines, 9(12), 1939. https://doi.org/10.3390/biomedicines9121939