
Neuromodulation Induced by Sitagliptin: A New Strategy for Treating Diabetic Retinopathy

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
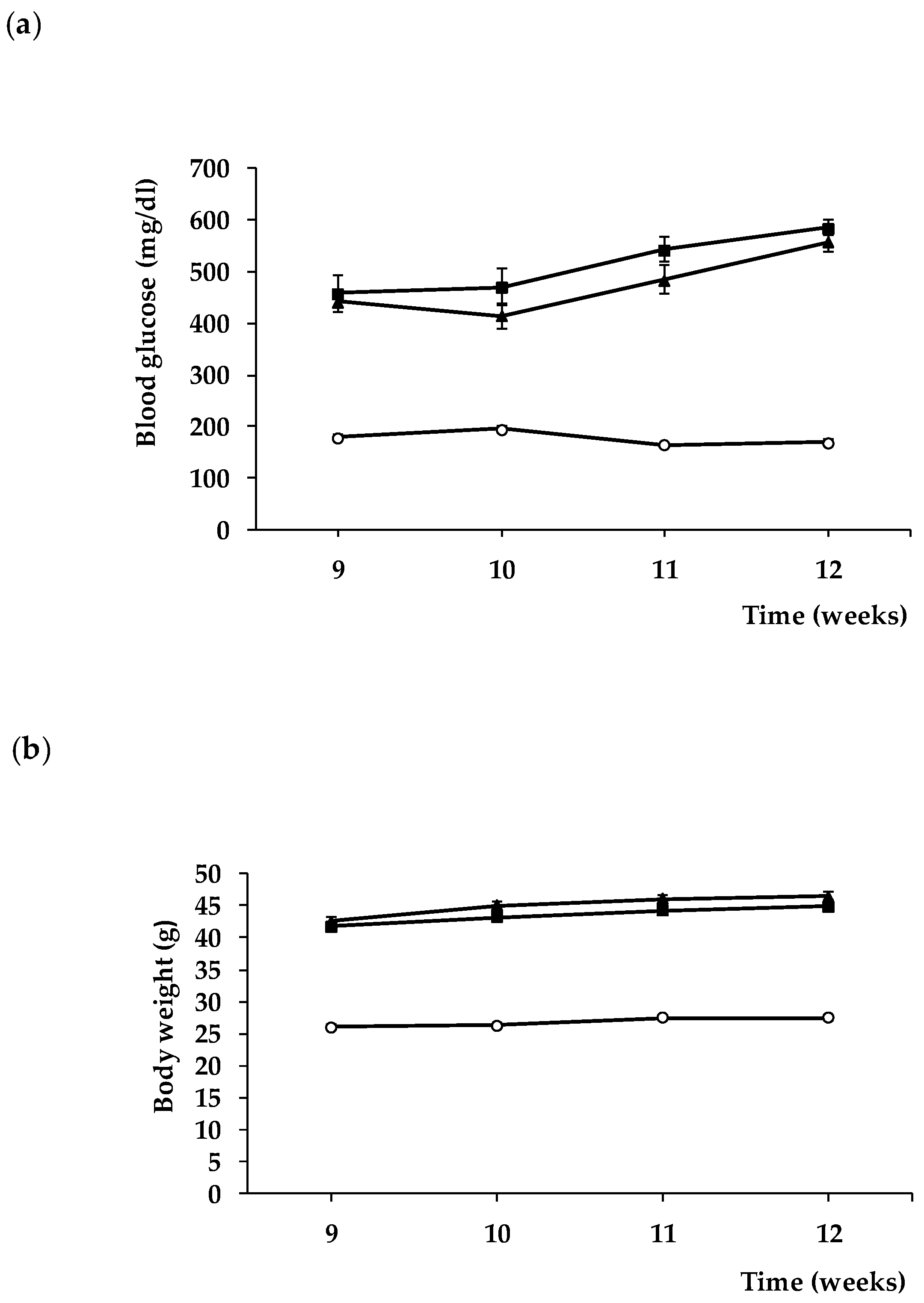
2.1. Animals
2.2. Topical Ocular Treatment
2.3. Electroretinogram (ERG)
2.4. Retinal Tissue Processing
2.4.1. RNA Extraction and Quantitative Reverse Transcription Polymerase Chain Reaction (RT-PCR) Assay
2.4.2. Western Blotting
2.4.3. Immunofluorescence Analysis
2.4.4. Statistical Analysis
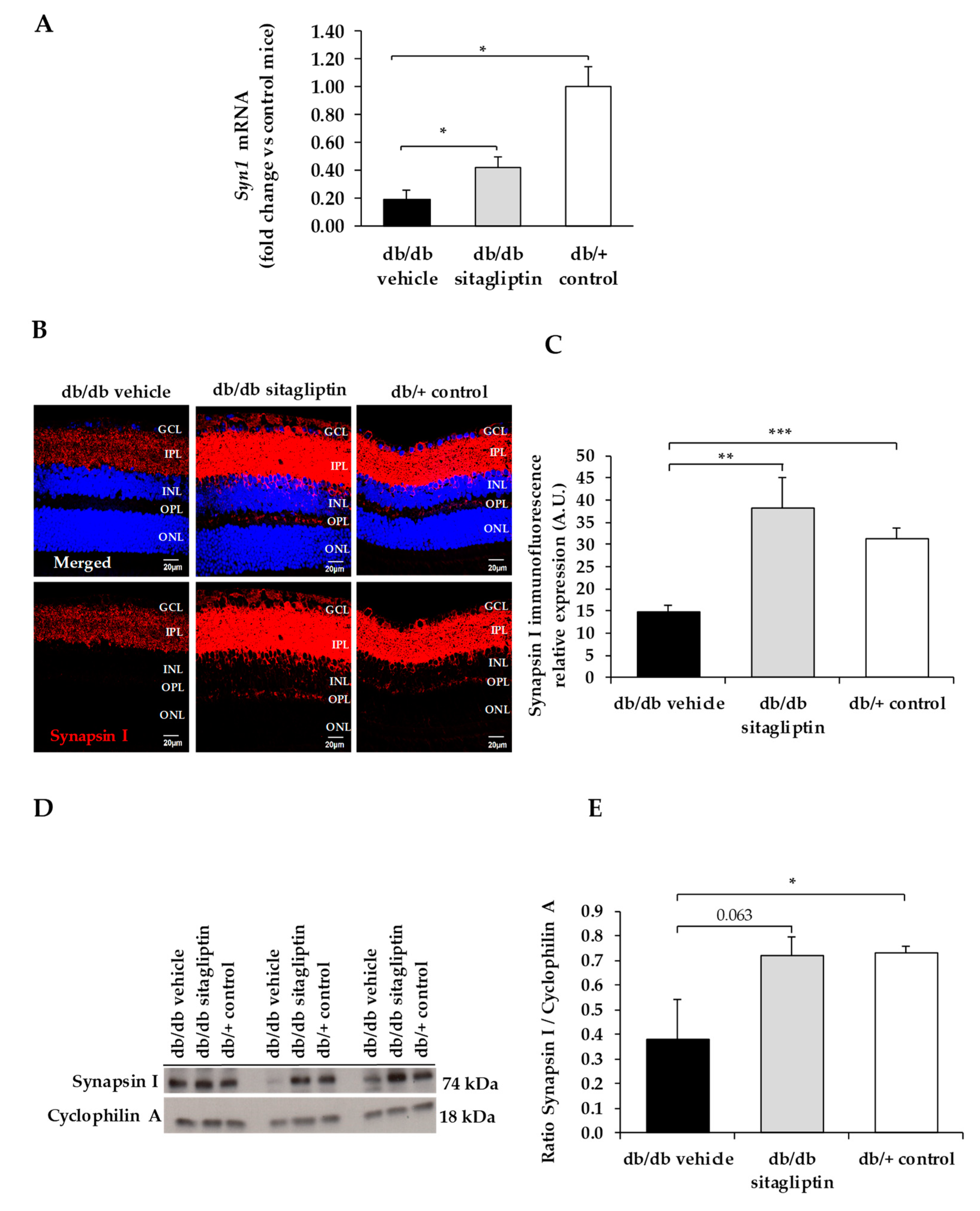
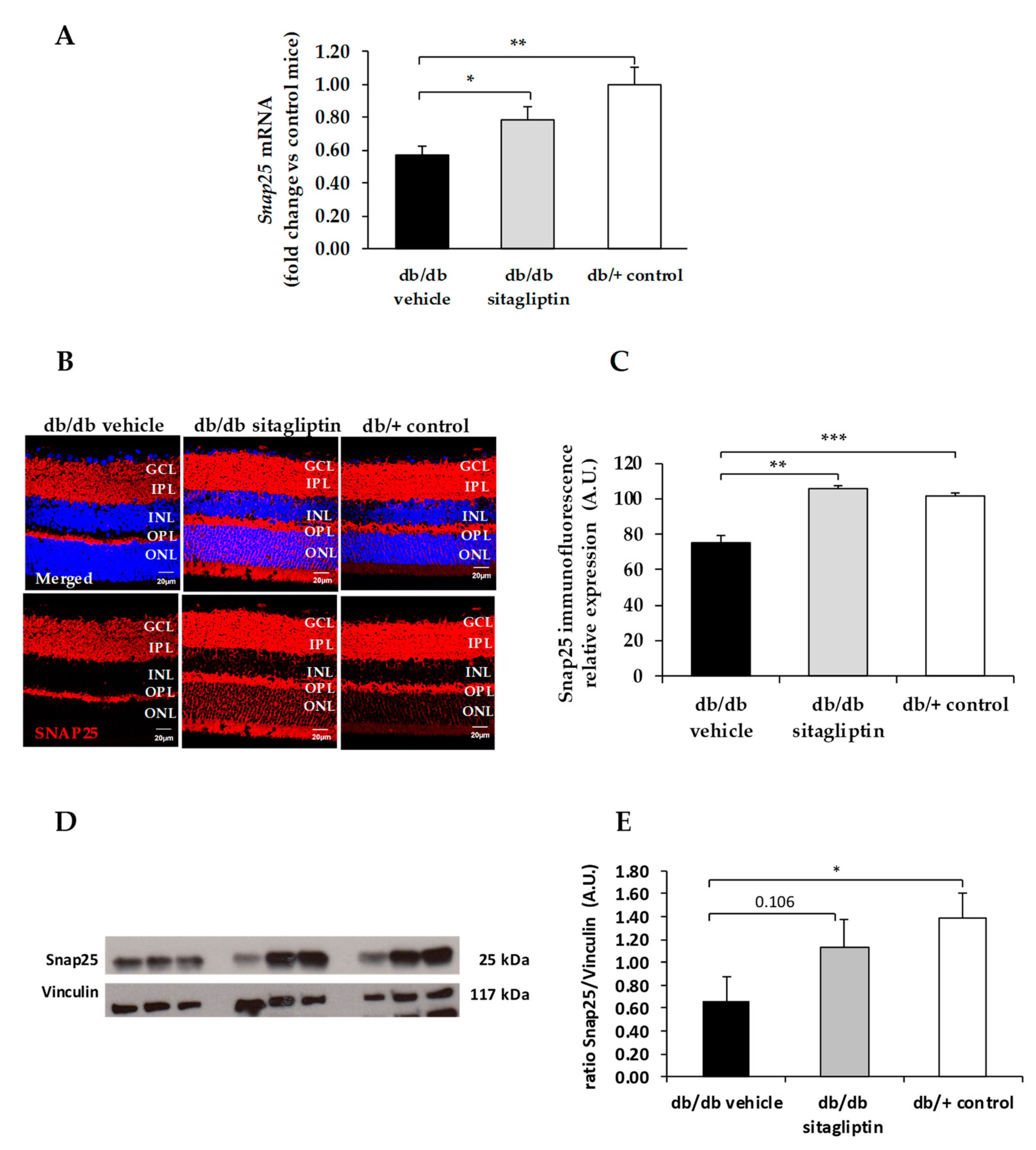
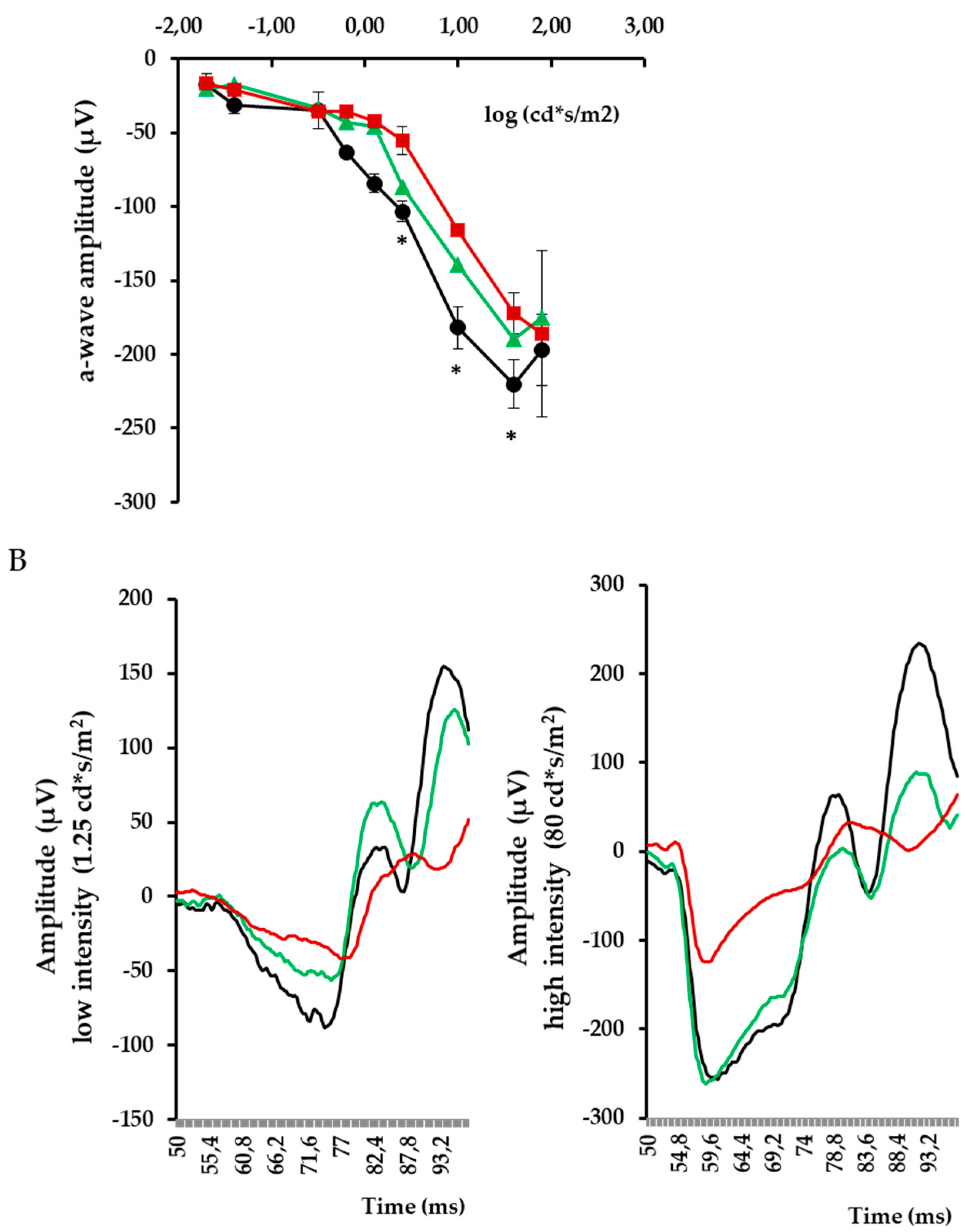
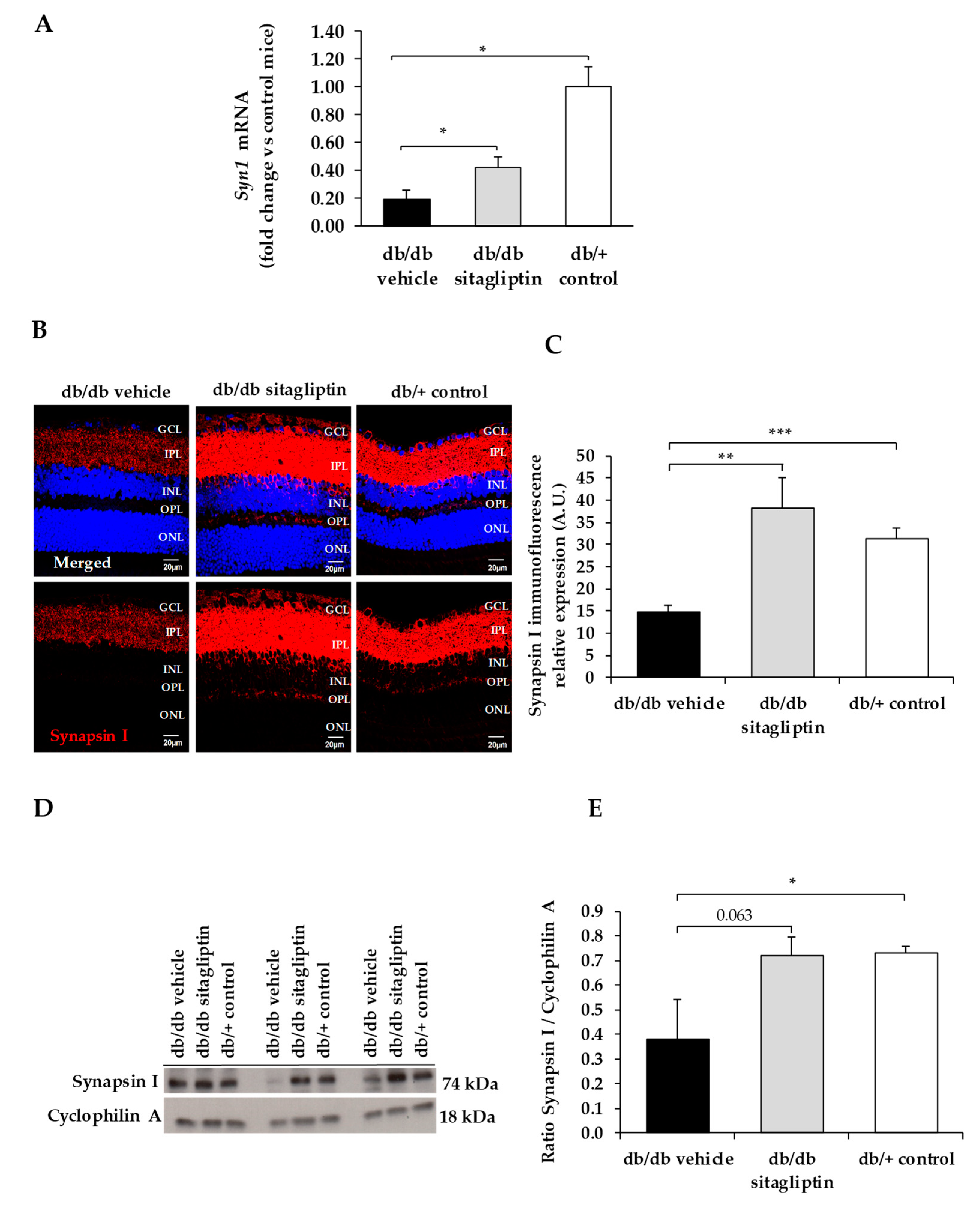
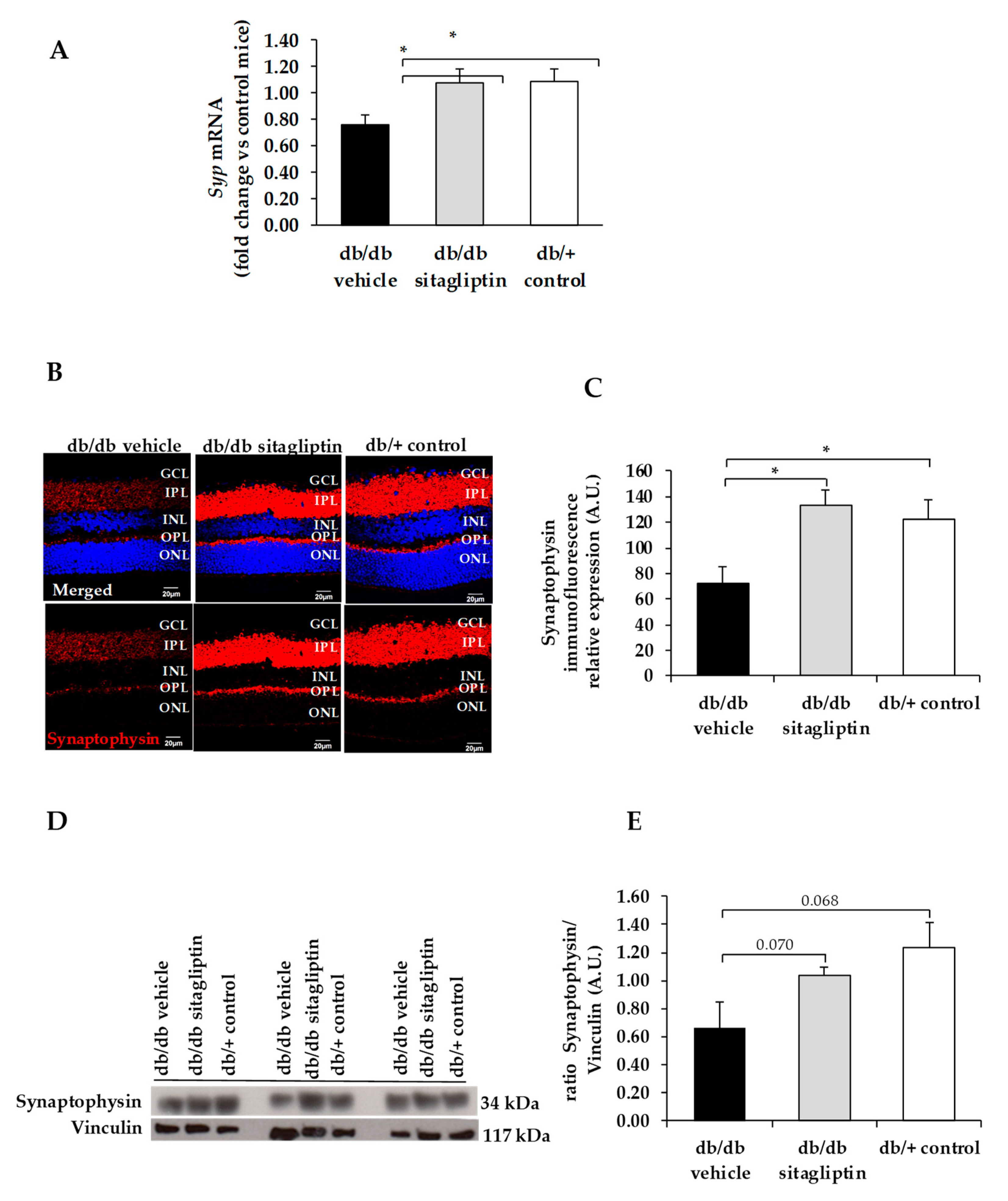
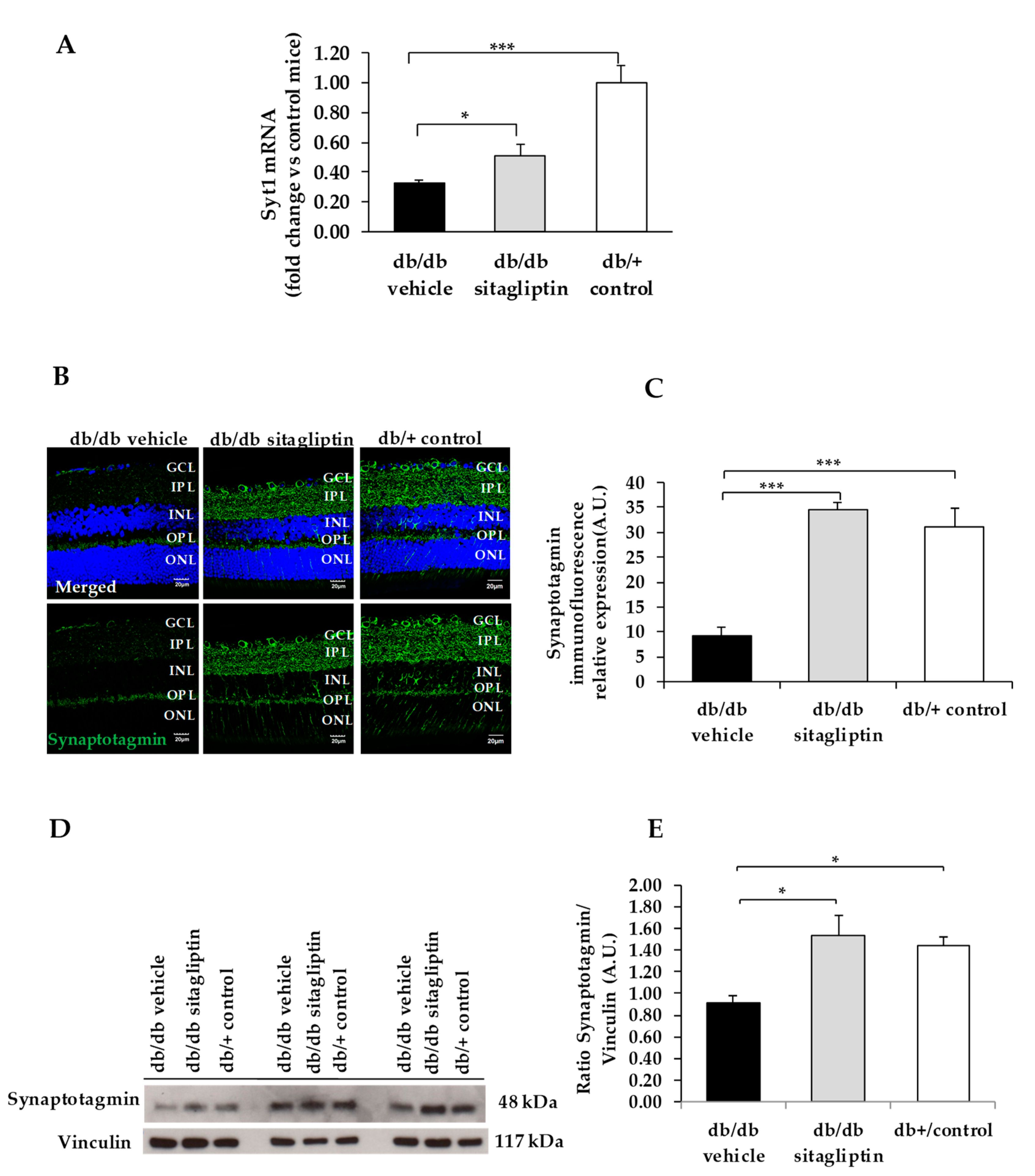
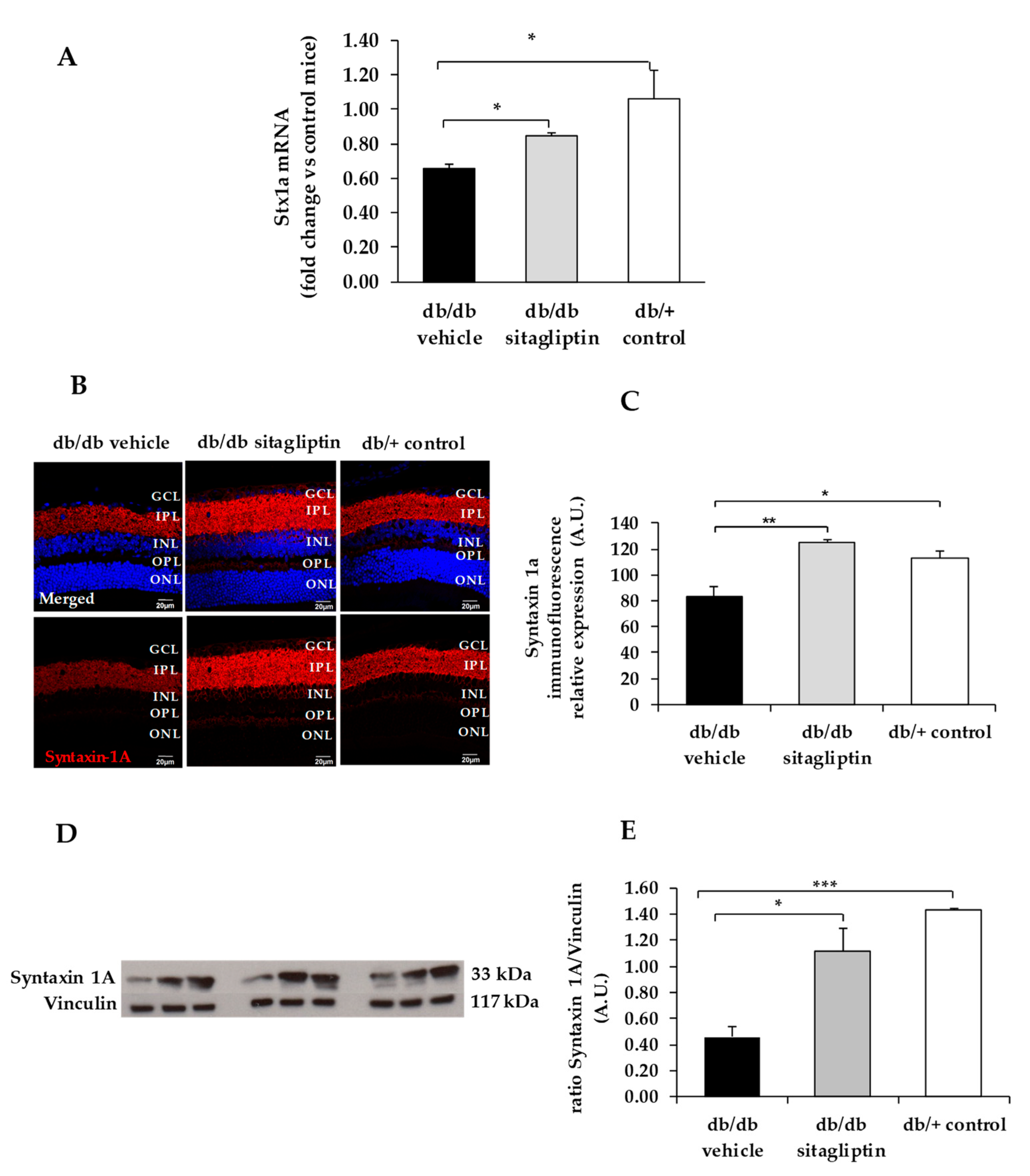
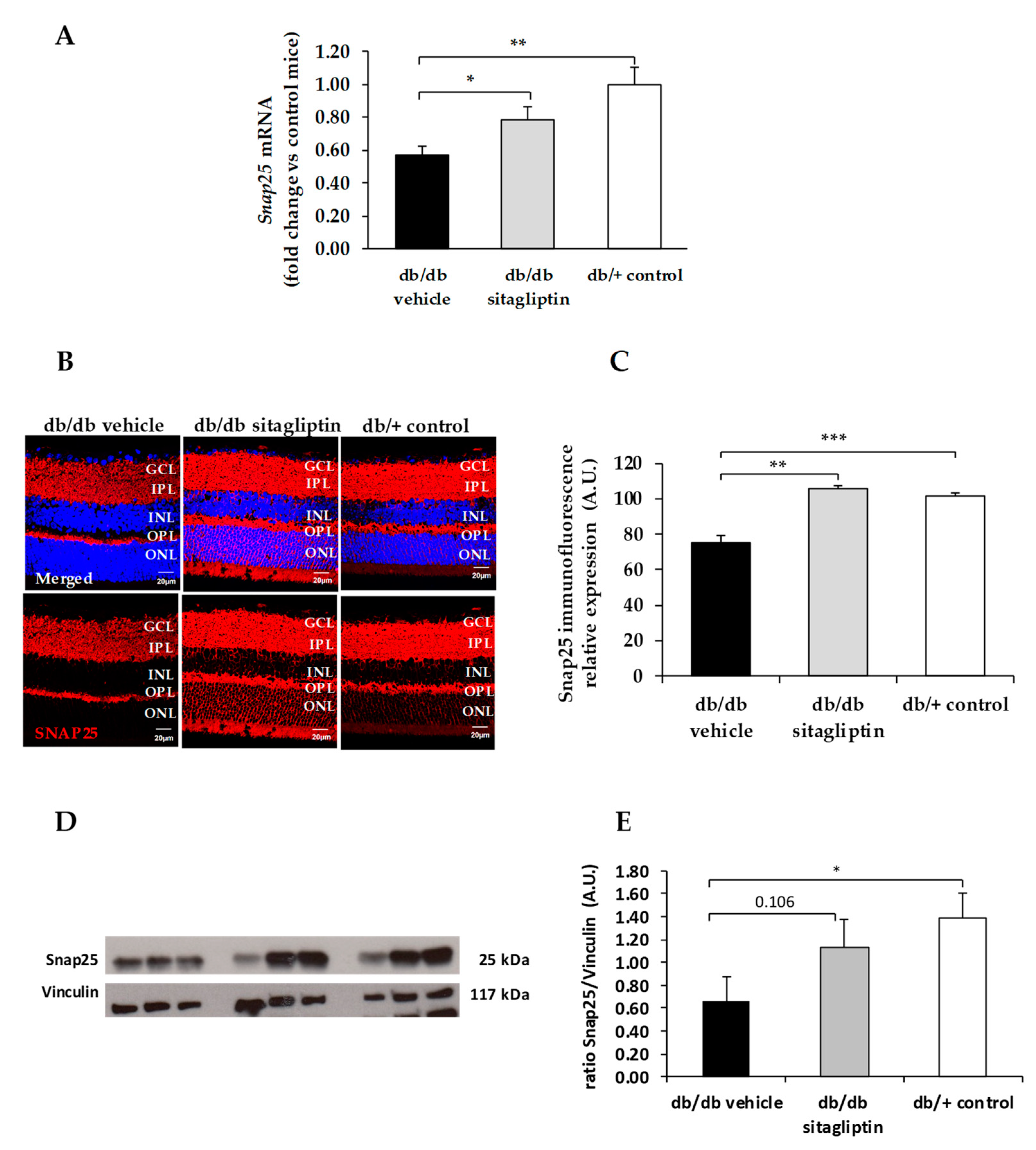
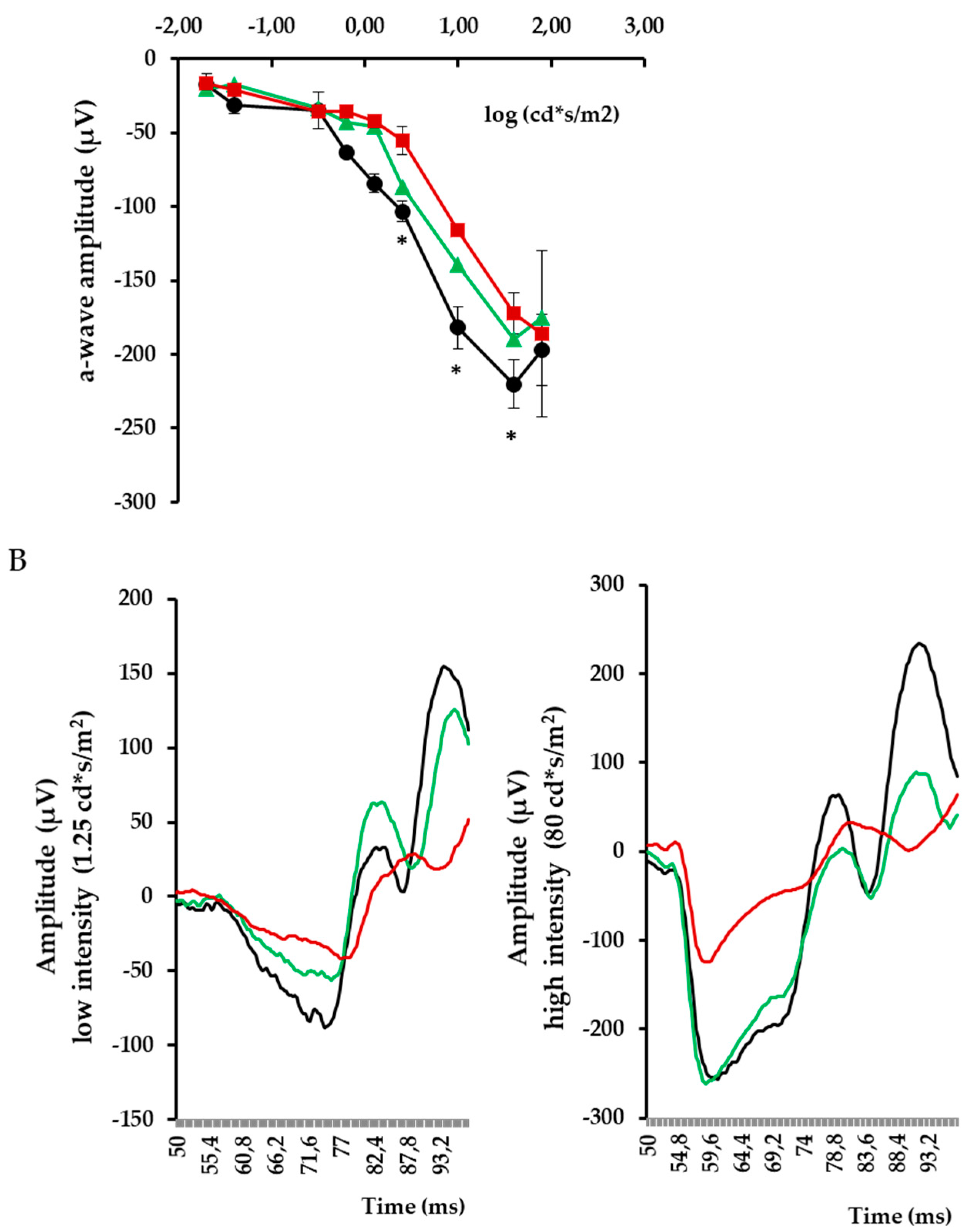
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simó, R.; Hernández, C. European Consortium for the Early Treatment of Diabetic Retinopathy (EUROCONDOR). Neurodegeneration in the diabetic eye: New insights and therapeutic perspectives. Trends Endocrinol. Metab. 2014, 25, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Simó, R.; Stitt, A.W.; Gardner, T.W. Neurodegeneration in diabetic retinopathy: Does it really matter? Diabetologia 2018, 61, 1902–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, S.D.; Chew, E.; Duh, E.J.; Sobrin, L.; Sun, J.K.; VanderBeek, B.L.; Wykoff, C.C.; Gardner, T.W. Diabetic retinopathy: A position statement by the American Diabetes Association. Diabetes Care 2017, 40, 412–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, A.J.; Lieth, E.; Khin, S.A.; Antonetti, D.A.; Buchanan, A.G.; Gardner, T.W. Neural apoptosis in the retina during experimental and human diabetes. Early onset and effect of insulin. J. Clin. Investig. 1998, 102, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrasco, E.; Hernández, C.; Miralles, A.; Huguet, P.; Farrés, J.; Simó, R. Lower somatostatin expression is an early event in diabetic retinopathy and is associated with retinal neurodegeneration. Diabetes Care 2007, 30, 2902–2908. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ramírez, M.; Hernández, C.; Villarroel, M.; Canals, F.; Alonso, M.A.; Fortuny, R.; Masmiquel, L.; Navarro, A.; García-Arumí, J.; Simó, R. Interphotoreceptor retinoid-binding protein (IRBP) is downregulated at early stages of diabetic retinopathy. Diabetologia 2009, 52, 2633–2641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, A.J.; Gardner, T.W.; Abcouwer, S.F. The significance of vascular and neural apoptosis to the pathology of diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1156–1163. [Google Scholar] [CrossRef]
- VanGuilder, H.D.; Brucklacher, R.M.; Patel, K.; Ellis, R.W.; Freeman, W.M.; Barber, A.J. Diabetes downregulates presynaptic proteins and reduces basal synapsin I phosphorylation in rat retina. Eur. J. Neurosci. 2008, 28, 1–11. [Google Scholar] [CrossRef]
- Gaspar, J.M.; Baptista, F.I.; Galvão, J.; Castilho, A.F.; Cunha, R.A.; Ambrósio, A.F. Diabetes differentially affects the content of exocytotic proteins in hippocampal and retinal nerve terminals. Neuroscience 2010, 169, 1589–1600. [Google Scholar] [CrossRef]
- Ly, A.; Scheerer, M.F.; Zukunft, S.; Muschet, C.; Merl, J.; Adamski, J.; de Angelis, M.H.; Neschen, S.; Hauck, S.M.; Ueffing, M. Retinal proteome alterations in a mouse model of type 2 diabetes. Diabetologia 2014, 57, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Robinson, W.F.; Van Guilder, H.D.; D’Cruz, T.S.; El-Remessy, A.B.; Barber, A.J. Synapsin 1 Protein Expression and Phosphorylation Are Compromised by Diabetes in Rodent and Human Retinas. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4920. [Google Scholar]
- Hernández, C.; Bogdanov, P.; Solà-Adell, C.; Sampedro, J.; Valeri, M.; Genís, X.; Simó-Servat, O.; García-Ramírez, M.; Simó, R. Topical administration of DPP-IV inhibitors prevents retinal neurodegeneration in experimental diabetes. Diabetologia 2017, 60, 2285–2298. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, P.; Corraliza, L.; Villena, J.A.; Carvalho, A.R.; García-Arumí, J.; Ramos, D.; Ruberte, J.; Simó, R.; Hernández, C. The db/db mouse: A useful model for the study of diabetic retinal neurodegeneration. PLoS ONE 2014, 9, e97302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marmor, M.F.; Holder, G.E.; Seeliger, M.W.; Yamamoto, S.; International Society for Clinical Electrophysiology of Vision. Standard for clinical electroretinography (2004 update). Doc. Ophthalmol. 2004, 108, 107–114. [Google Scholar] [CrossRef]
- Simó, R.; Simó-Servat, O.; Bogdanov, P.; Hernández, C. Neurovascular Unit: A New Target for Treating Early Stages of Diabetic Retinopathy. Pharmaceutics 2021, 13, 1320. [Google Scholar] [CrossRef] [PubMed]
- Micheva, K.D.; Busse, B.; Weiler, N.C.; O’Rourke, N.; Smith, S.J. Single-synapse analysis of a diverse synapse population: Proteomic imaging methods and markers. Neuron 2010, 68, 639–653. [Google Scholar] [CrossRef] [Green Version]
- Huttner, W.B.; Schiebler, W.; Greengard, P.; De Camilli, P. Synapsin I (protein I), a nerve terminal-specific phosphoprotein. III. Its association with synaptic vesicles studied in a highly purified synaptic vesicle preparation. J. Cell Biol. 1983, 96, 1374–1388. [Google Scholar] [CrossRef]
- Bähler, M.; Benfenati, F.; Valtorta, F.; Greengard, P. The synapsins and the regulation of synaptic function. BioEssays News Rev. Mol. Cell. Dev. Biol. 1990, 12, 259–263. [Google Scholar] [CrossRef]
- Bykhovskaia, M. Synapsin regulation of vesicle organization and functional pools. Semin. Cell Dev. Biol. 2011, 22, 387–392. [Google Scholar] [CrossRef]
- Wiedenmann, B.; Franke, W.W. Identification and localization of synaptophysin, an integral membrane glycoprotein of Mr 38,000 characteristic of presynaptic vesicles. Cell 1985, 41, 1017–1028. [Google Scholar] [CrossRef]
- Jahn, R.; Schiebler, W.; Ouimet, C.; Greengard, P.A. 38,000-dalton membrane protein (p38) present in synaptic vesicles. Proc. Natl. Acad. Sci. USA 1985, 82, 4137–4141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takamori, S.; Holt, M.; Stenius, K.; Lemke, E.A.; Gronborg, M.; Riedel, D.; Urlaub, H.; Schenck, S.; Brugger, B.; Ringler, P.; et al. Molecular anatomy of a trafficking organelle. Cell 2006, 127, 831–846. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.E.; Chapman, E.R. Synaptophysin regulates the kinetics of synaptic vesicle endocytosis in central neurons. Neuron 2011, 70, 847–854. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.L.; Leube, R.E.; Cousin, M.A. Synaptophysin is required for synaptobrevin retrieval during synaptic vesicle endocytosis. J. Neurosci. 2011, 31, 14032–14036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishnan, N.A.; Drescher, M.J.; Drescher, D.G. The SNARE complex in neuronal and sensory cells. Mol. Cell Neurosci. 2012, 50, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Masser, D.R.; VanGuilder, H.D.; Bixler, G.V.; Dunton, W.; Bronson, S.K.; Freeman, W.M. Insulin treatment normalizes retinal neuroinflammation but not markers of synapse loss in diabetic rats. Exp. Eye Res. 2014, 125, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, C.; Bogdanov, P.; Corraliza, L.; García-Ramírez, M.; Solà-Adell, C.; Arranz, J.A.; Arroba, A.I.; Valverde, A.M.; Simó, R. Topical administration of GLP-1 receptor agonists prevents retinal neurodegeneration in experimental diabetes. Diabetes 2016, 65, 172–187. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Xu, J.; Sun, Q.; Yu, F.; Cheng, J.; Peng, B.; Liu, W.; Xiao, Z.; Yin, J.; et al. Inhibition of DPP4 enhances inhibitory synaptic transmission through activating the GLP-1/GLP-1R signaling pathway in a rat model of febrile seizures. Biochem. Pharmacol. 2018, 156, 78–85. [Google Scholar] [CrossRef]
- Dietrich, N.; Kolibabka, M.; Busch, S.; Bugert, P.; Kaiser, U.; Lin, J.; Fleming, T.; Morcos, M.; Klein, T.; Schlotterer, A.; et al. The DPP4 inhibitor linagliptin protects from experimental diabetic retinopathy. PLoS ONE 2016, 11, e0167853. [Google Scholar] [CrossRef]
- Király, K.; Kozsurek, M.; Lukácsi, E.; Barta, B.; Alpár, A.; Balázsa, T.; Fekete, C.; Szabon, J.; Helyes, Z.; Bölcskei, K.; et al. Glial cell type-specific changes in spinal dipeptidyl peptidase 4 expression and effects of its inhibitors in inflammatory and neuropatic pain. Sci. Rep. 2018, 8, 3490. [Google Scholar] [CrossRef]
- Yau, J.W.; Rogers, S.L.; Kawasaki, R.; Lamoureux, E.L.; Kowalski, J.W.; Bek, T.; Chen, S.J.; Dekker, J.M.; Fletcher, A.; Grauslund, J.; et al. Global prevalence and major risk factors of diabetic retinopathy. Diabetes Care 2012, 35, 556–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leasher, J.L.; Bourne, R.R.; Flaxman, S.R.; Jonas, J.B.; Keeffe, J.; Naidoo, K.; Pesudovs, K.; Price, H.; White, R.A.; Wong, T.Y.; et al. Global Estimates on the Number of People Blind or Visually Impaired by Diabetic Retinopathy: A Meta-analysis from 1990 to 2010. Diabetes Care 2016, 39, 1643–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Nucleotide Sequence | |
|---|---|---|
| B2m | Forward (5′-3′) | 5′-GTATGCTATCCAGAAAACCC-3′ |
| Reverse (5′-3′) | 5′-CTGAAGGACATATCTGACATC-3′ | |
| Actb | Forward (5′-3′) | 5′-CTAAGGCCAACCGTGAAAG -3′ |
| Reverse (5′-3′) | 5′-CAGTATGTTCGGCTTCCCATTC-3′ | |
| Syn1 | Forward (5′-3′) | 5′-AATCACAAAGAGATGCTCAG-3′ |
| Reverse (5′-3′) | 5′-GGACACGCACATCATATTTAG-3′ | |
| Syp | Forward (5′-3′) | 5′-TGCCAACAAGACGGAGAGTG-3′ |
| Reverse (5′-3′) | 5′-TAGTGCCCCCTTTAACGCAG-3′ | |
| Syt1 | Forward (5′-3′) | 5′-ACCCTGGGCTCTGTATCCC-3′ |
| Reverse (5′-3′) | 5′-CCCTGACCACTGAGTGCAAA-3′ | |
| Stx1a | Forward (5′-3′) | 5′-CGCTGTCCCGAAAGTTTGTG-3′ |
| Reverse (5′-3′) | 5′-GTGTCTGGTCTCGATCTCACT-3′ | |
| Vamp2 | Forward (5′-3′) | 5′-ATCATCGTTTACTTCAGCAC-3′ |
| Reverse (5′-3′) | 5′-TGAAAGATATGGCTGAGAGG-3′ | |
| Snap25 | Forward (5′-3′) | 5′-CAACTGGAACGCATTGAGGAA-3′ |
| Reverse (5′-3′) | 5′-GGCCACTACTCCATCCTGATTAT-3′ |
| Primary Antibodies | Description |
|---|---|
| Synapsin I | Rabbit polyclonal; 1:2000; ab64581; Abcam, Cambridge, UK |
| Synaptophysin | Rabbit monoclonal; 1:200,000; ab32127; Abcam, Cambridge, UK |
| Synaptotagmin | Mouse monoclonal; 1:1,000; ab13259; Abcam, Cambridge, UK |
| Syntaxin 1A | Rabbit polyclonal; 1:100,000; ab41453; Abcam, Cambridge, UK |
| SNAP-25 | Rabbit polyclonal; 1:1000; 14903-1-AP; Proteintech, Rosemont, IL, USA |
| Vamp2 | Rabbit polyclonal; 1:1000; 10135-1-AP; Proteintech, Rosemont, IL, USA |
| Cyclophilin A | 1:10,000; BML-SA296; Enzo, NY, USA |
| Vinculin | Mouse monoclonal; 1:7000; sc-73614; Santa Cruz, Dallas, TX, USA |
| Primary Antibodies | Description |
| Synapsin I | Rabbit polyclonal; 1:100; ab64581; Abcam, Cambridge, UK |
| Synaptophysin | Rabbit monoclonal; 1:100; ab32127; Abcam, Cambridge, UK |
| Synaptotagmin | Mouse monoclonal; 1:200; ab13259; Abcam, Cambridge, UK |
| Syntaxin 1A | Rabbit polyclonal; 1:200; ab41453; Abcam, Cambridge, UK |
| Vamp2 | Rabbit polyclonal; 1:100; 10135-1-AP; Proteintech, Rosemont, IL, USA |
| SNAP-25 | Rabbit polyclonal; 1:100; 14903-1-AP; Proteintech, Rosemont, IL, USA |
| Secondary Antibodies | Description |
| Alexa Fluor 488 Goat anti-mouse | Goat polyclonal; 1:600; ab150113; Abcam, Cambridge, UK |
| Alexa Fluor 488 Goat anti-rabbit | Goat polyclonal; 1:600; ab150081; Abcam, Cambridge, UK |
| Alexa Fluor 594 Goat anti-mouse | Goat polyclonal; 1:600; A-11032; ThermoFisher Scientific, Waltham, MA, USA |
| Alexa Fluor 594 Goat anti-rabbit | Goat polyclonal; 1:600; A-11012; ThermoFisher Scientific, Waltham, MA, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, H.; Bogdanov, P.; Sabater, D.; Huerta, J.; Valeri, M.; Hernández, C.; Simó, R. Neuromodulation Induced by Sitagliptin: A New Strategy for Treating Diabetic Retinopathy. Biomedicines 2021, 9, 1772. https://doi.org/10.3390/biomedicines9121772
Ramos H, Bogdanov P, Sabater D, Huerta J, Valeri M, Hernández C, Simó R. Neuromodulation Induced by Sitagliptin: A New Strategy for Treating Diabetic Retinopathy. Biomedicines. 2021; 9(12):1772. https://doi.org/10.3390/biomedicines9121772
Chicago/Turabian StyleRamos, Hugo, Patricia Bogdanov, David Sabater, Jordi Huerta, Marta Valeri, Cristina Hernández, and Rafael Simó. 2021. "Neuromodulation Induced by Sitagliptin: A New Strategy for Treating Diabetic Retinopathy" Biomedicines 9, no. 12: 1772. https://doi.org/10.3390/biomedicines9121772
APA StyleRamos, H., Bogdanov, P., Sabater, D., Huerta, J., Valeri, M., Hernández, C., & Simó, R. (2021). Neuromodulation Induced by Sitagliptin: A New Strategy for Treating Diabetic Retinopathy. Biomedicines, 9(12), 1772. https://doi.org/10.3390/biomedicines9121772