
CD63+ and MHC Class I+ Subsets of Extracellular Vesicles Produced by Wild-Type and CD47-Deficient Jurkat T Cells Have Divergent Functional Effects on Endothelial Cell Gene Expression

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. EV Extraction
2.3. Immunocapture of EVs
2.4. EV Size Characterization
2.5. Small RNA Sequencing Analysis
2.6. Microarray Expression and Gene Set Enrichment Analysis (GSEA)
2.7. EV Uptake
2.8. Incucyte EV Uptake Assay
2.9. Cytokine Analysis
2.10. Real-Time PCR
2.11. Flow Cytometry Analysis
2.12. Negative Stain Electron Microscopy and Immunogold Labeling
2.13. NanoSight Analysis
2.14. Supplemental RNASEQ and Microarray Data
3. Results
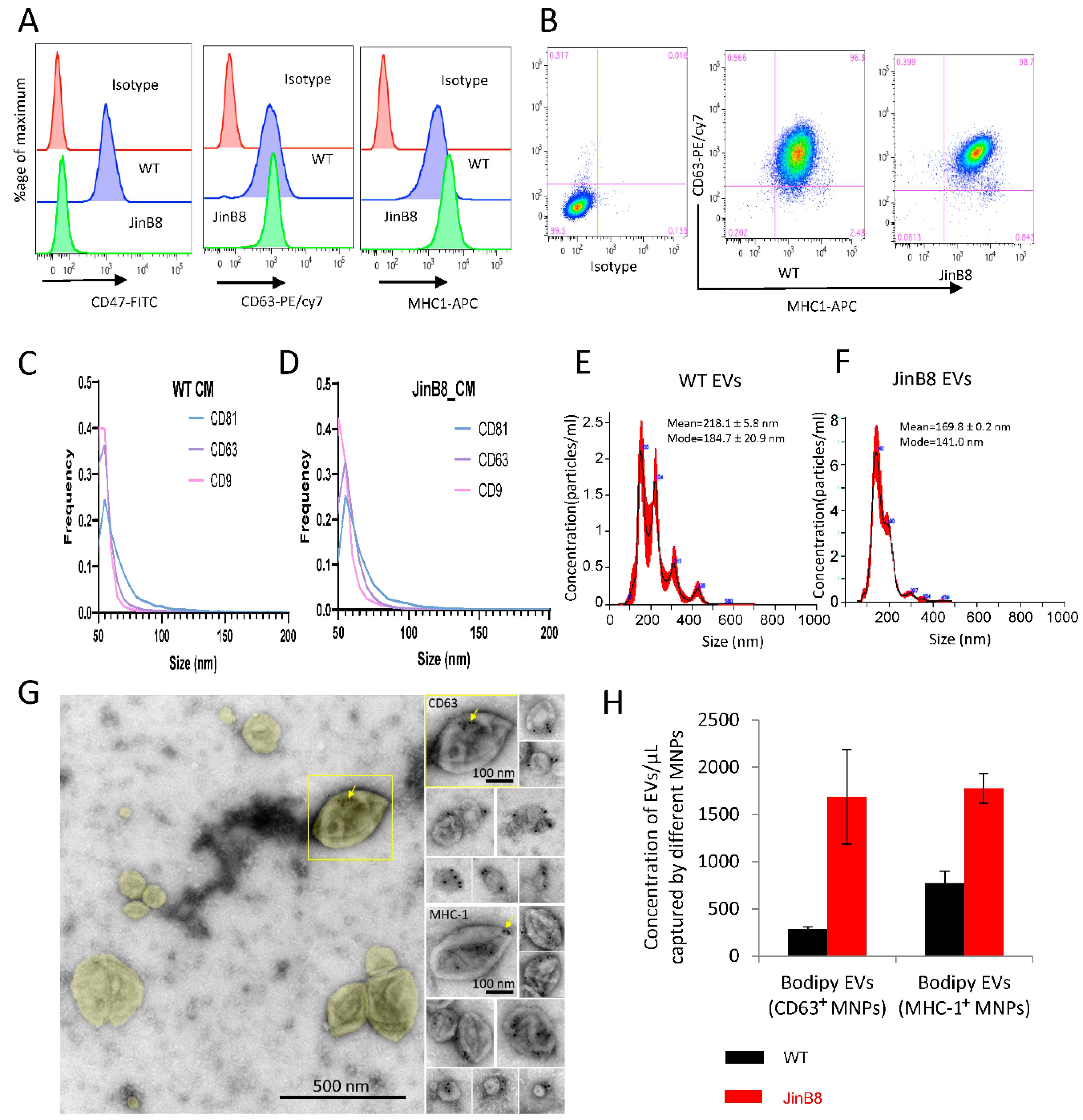
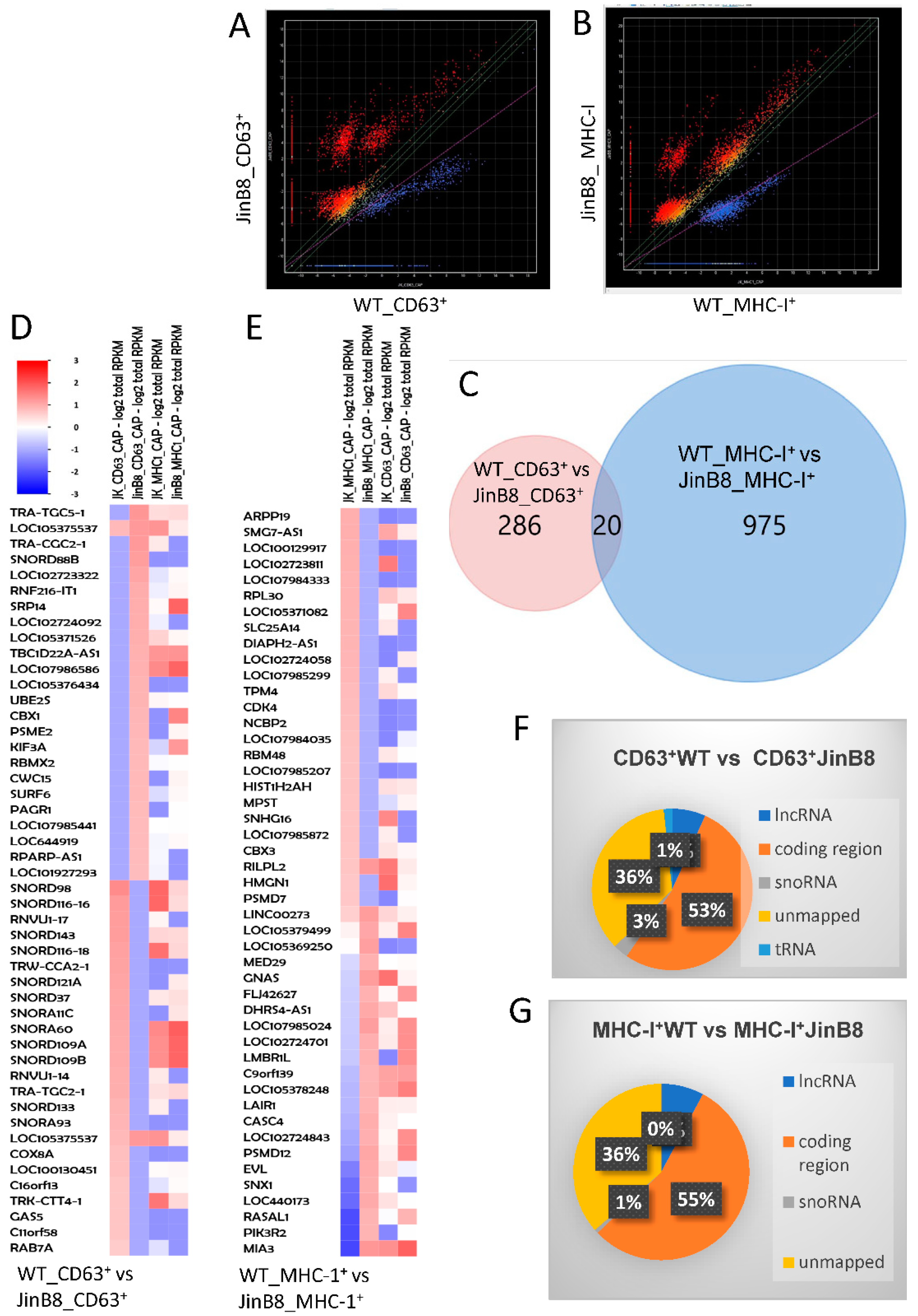
3.1. CD47 Regulates the Release of CD63+ and MHC-I+ EVs from Jurkat T Cells and Their RNA Contents
3.2. Selective Uptake of MHC-I+ and CD63+ EVs by Target Endothelial Cells
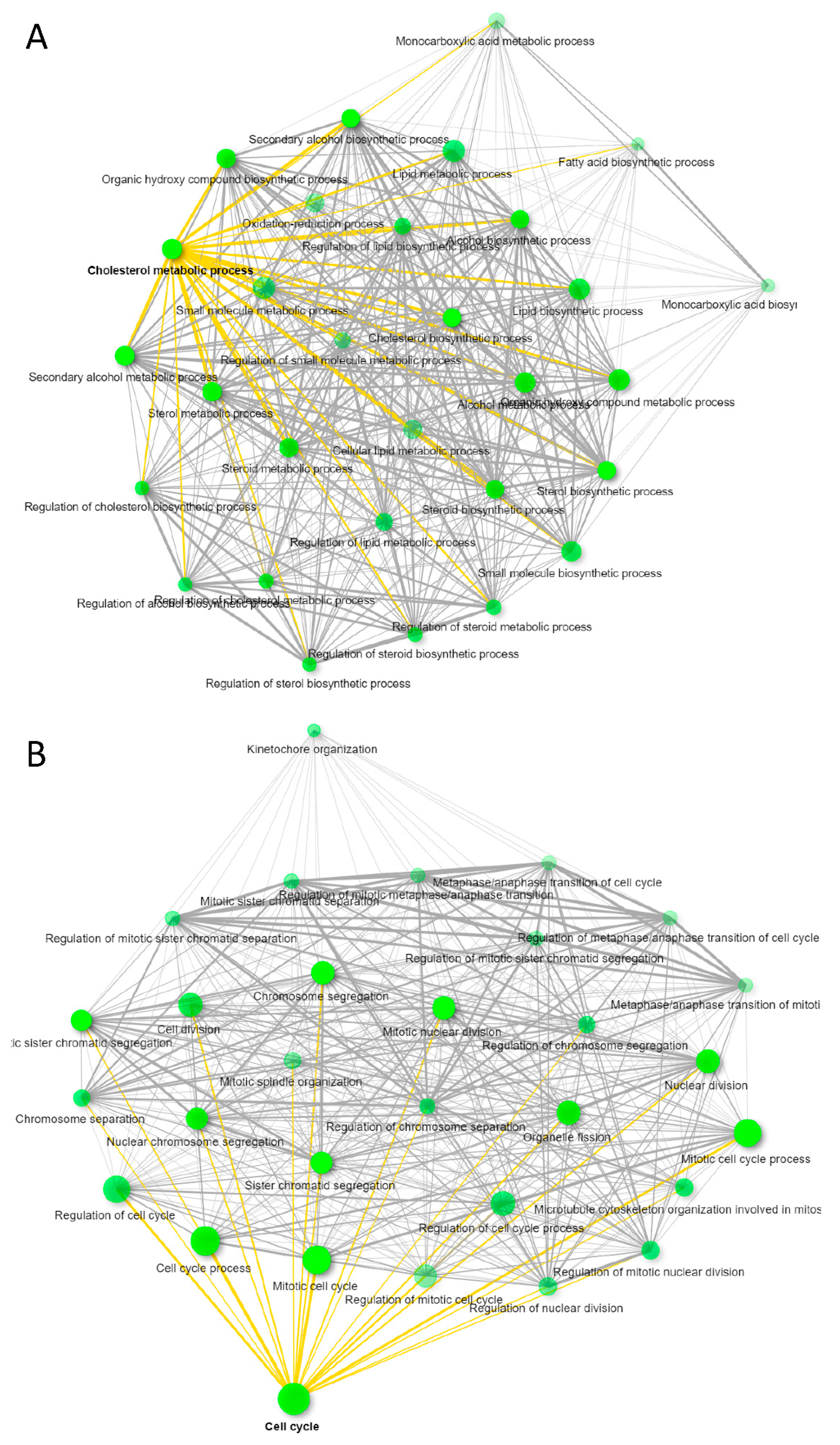
3.3. CD63+ and MHC-I+ EVs from WT Jurkat T Cells Regulate Different Molecular and Functional Programs in Recipient Endothelial Cells
3.4. MHC-I+ EVs Upregulate More CD47-Dependent Transcripts in HUVEC Than CD63+ EVs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- McKelvey, K.J.; Powell, K.L.; Ashton, A.W.; Morris, J.M.; McCracken, S.A. Exosomes: Mechanisms of Uptake. J. Circ. Biomark. 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morelli, A.E.; Larregina, A.T.; Shufesky, W.J.; Sullivan, M.L.; Stolz, D.B.; Papworth, G.D.; Zahorchak, A.F.; Logar, A.J.; Wang, Z.; Watkins, S.C.; et al. Endocytosis, intracellular sorting, and processing of exosomes by dendritic cells. Blood 2004, 104, 3257–3266. [Google Scholar] [CrossRef] [Green Version]
- Gong, M.; Yu, B.; Wang, J.; Wang, Y.; Liu, M.; Paul, C.; Millard, R.W.; Xiao, D.S.; Ashraf, M.; Xu, M. Mesenchymal stem cells release exosomes that transfer miRNAs to endothelial cells and promote angiogenesis. Oncotarget 2017, 8, 45200–45212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, A.; Yadav, J.; Thakur, K.; Aggarwal, N.; Tripathi, T.; Chhokar, A.; Singh, T.; Jadli, M.; Bharti, A.C. Exosomes from cervical cancer cells facilitate pro-angiogenic endothelial reconditioning through transfer of Hedgehog-GLI signaling components. Cancer Cell Int. 2021, 21, 319. [Google Scholar] [CrossRef]
- Saez, T.; de Vos, P.; Kuipers, J.; Sobrevia, L.; Faas, M.M. Exosomes derived from monocytes and from endothelial cells mediate monocyte and endothelial cell activation under high d-glucose conditions. Immunobiology 2019, 224, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, A.; Sawada, K.; Kobayashi, M.; Yamamoto, M.; Yagi, T.; Kinose, Y.; Kodama, M.; Hashimoto, K.; Kimura, T. Exosomal CD47 Plays an Essential Role in Immune Evasion in Ovarian Cancer. Mol. Cancer Res. 2021, 19, 1583–1595. [Google Scholar] [CrossRef] [PubMed]
- Kugeratski, F.G.; Hodge, K.; Lilla, S.; McAndrews, K.M.; Zhou, X.; Hwang, R.F.; Zanivan, S.; Kalluri, R. Quantitative proteomics identifies the core proteome of exosomes with syntenin-1 as the highest abundant protein and a putative universal biomarker. Nat. Cell Biol. 2021, 23, 631–641. [Google Scholar] [CrossRef]
- Fenselau, C.; Ostrand-Rosenberg, S. Molecular cargo in myeloid-derived suppressor cells and their exosomes. Cell Immunol. 2021, 359, 104258. [Google Scholar] [CrossRef]
- Kibria, G.; Ramos, E.K.; Lee, K.E.; Bedoyan, S.; Huang, S.; Samaeekia, R.; Athman, J.J.; Harding, C.V.; Lotvall, J.; Harris, L.; et al. A rapid, automated surface protein profiling of single circulating exosomes in human blood. Sci. Rep. 2016, 6, 36502. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, S.P.; Elkahloun, A.G.; Wu, W.; Abu-Asab, M.S.; Roberts, D.D. CD47-dependent immunomodulatory and angiogenic activities of extracellular vesicles produced by T cells. Matrix Biol. 2014, 37, 49–59. [Google Scholar] [CrossRef]
- Kaur, S.; Elkahloun, A.G.; Singh, S.P.; Arakelyan, A.; Roberts, D.D. A function-blocking CD47 antibody modulates extracellular vesicle-mediated intercellular signaling between breast carcinoma cells and endothelial cells. J. Cell Commun. Signal. 2018, 12, 157–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Elkahloun, A.G.; Arakelyan, A.; Young, L.; Myers, T.G.; Otaizo-Carrasquero, F.; Wu, W.; Margolis, L.; Roberts, D.D. CD63, MHC class 1, and CD47 identify subsets of extracellular vesicles containing distinct populations of noncoding RNAs. Sci. Rep. 2018, 8, 2577. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, M.I.; Green, J.M.; Lindberg, F.P.; Ticchioni, M.; Brown, E.J. Cell spreading distinguishes the mechanism of augmentation of T cell activation by integrin-associated protein/CD47 and CD28. Int. Immunol. 1999, 11, 707–718. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Kuznetsova, S.A.; Pendrak, M.L.; Sipes, J.M.; Romeo, M.J.; Li, Z.; Zhang, L.; Roberts, D.D. Heparan sulfate modification of the transmembrane receptor CD47 is necessary for inhibition of T cell receptor signaling by thrombospondin-1. J. Biol. Chem. 2011, 286, 14991–15002. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Saldana, C.A.; Elkahloun, A.G.; Petersen, J.D.; Arakelyan, A.; Singh, P.S.; Jenkins, L.M.; Kuo, B.; Reginauld, B.; Jordan, J.D.; et al. CD47 interactions with exportin-1 regulate targeting of m7G-modified RNAs to extracellular vesicles. J. Cell Commun. Signal. 2021, in press. [Google Scholar]
- Arakelyan, A.; Petersen, J.D.; Blazkova, J.; Margolis, L. Macrophage-derived HIV-1 carries bioactive TGF-beta. Sci. Rep. 2019, 9, 19100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Chang, T.; Singh, S.P.; Lim, L.; Mannan, P.; Garfield, S.H.; Pendrak, M.L.; Soto-Pantoja, D.R.; Rosenberg, A.Z.; Jin, S.; et al. CD47 signaling regulates the immunosuppressive activity of VEGF in T cells. J. Immunol. 2014, 193, 3914–3924. [Google Scholar] [PubMed] [Green Version]
- Citrin, K.M.; Fernandez-Hernando, C.; Suarez, Y. MicroRNA regulation of cholesterol metabolism. Ann. N. Y. Acad. Sci. 2021, 1495, 55–77. [Google Scholar] [CrossRef]
- Fang, L.; Liu, C.; Miller, Y.I. Zebrafish models of dyslipidemia: Relevance to atherosclerosis and angiogenesis. Transl. Res. 2014, 163, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Lyu, J.; Yang, E.J.; Shim, J.S. Cholesterol Trafficking: An Emerging Therapeutic Target for Angiogenesis and Cancer. Cells 2019, 8, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lightbody, R.J.; Taylor, J.M.W.; Dempsie, Y.; Graham, A. Induction of microRNA hsa-let-7d-5p, and repression of HMGA2, contribute protection against lipid accumulation in macrophage ‘foam’ cells. Biochim. Biophys. Acta Mol. Cell Biol Lipids 2021, 1866, 159005. [Google Scholar] [CrossRef] [PubMed]
- Lidington, E.A.; Moyes, D.L.; McCormack, A.M.; Rose, M.L. A comparison of primary endothelial cells and endothelial cell lines for studies of immune interactions. Transpl. Immunol. 1999, 7, 239–246. [Google Scholar] [CrossRef]
- Barkal, A.A.; Weiskopf, K.; Kao, K.S.; Gordon, S.R.; Rosental, B.; Yiu, Y.Y.; George, B.M.; Markovic, M.; Ring, N.G.; Tsai, J.M.; et al. Engagement of MHC class I by the inhibitory receptor LILRB1 suppresses macrophages and is a target of cancer immunotherapy. Nat. Immunol. 2018, 19, 76–84. [Google Scholar] [CrossRef]
- Lv, N.; Liu, F.; Cheng, L.; Liu, F.; Kuang, J. The Expression of Transcription Factors is Different in Papillary Thyroid Cancer Cells during TNF-alpha induced EMT. J. Cancer 2021, 12, 2777–2786. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shi, L.; Wang, Y.; Ye, D.; Ju, H.; Ma, H.; Yang, W.; Wang, Y.; Hu, J.; Deng, J.; et al. Stabilization of Slug by NF-kappaB is Essential for TNF-alpha -Induced Migration and Epithelial-Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma Cells. Cell Physiol. Biochem. 2018, 47, 567–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Function 1 | CD63+ EV vs. UT (Fold) | p Value | MHC-I+ EV vs. UT (Fold) | p Value | miRNAs Differing in CD63+ vs. MHC-I+ EVs 2 |
|---|---|---|---|---|---|---|
| LDLR | cholesterol uptake | 2.0 | 2.8 × 10−6 | 1.93 | 5.2 × 10−6 | miR-140, miR-185, miR-130b |
| SREBF2 | cholesterol biosynthesis | 1.5 | 0.00032 | NS | miR-185 | |
| INSIG1 | cholesterol biosynthesis | 2.81 | 2.6 × 10−5 | 1.97 | 0.00124 | miR-130b |
| HMGCR | cholesterol biosynthesis | 2.1 | 0.032 | NS | ||
| HMGCS1 | cholesterol biosynthesis | 2.99 | 0.0052 | NS | miR-223 | |
| ACAT2 | lipid/cholesterol metabolism | 2.3 | 0.014 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, S.; Elkahloun, A.G.; Petersen, J.D.; Arakelyan, A.; Livak, F.; Singh, S.P.; Margolis, L.; Zimmerberg, J.; Roberts, D.D. CD63+ and MHC Class I+ Subsets of Extracellular Vesicles Produced by Wild-Type and CD47-Deficient Jurkat T Cells Have Divergent Functional Effects on Endothelial Cell Gene Expression. Biomedicines 2021, 9, 1705. https://doi.org/10.3390/biomedicines9111705
Kaur S, Elkahloun AG, Petersen JD, Arakelyan A, Livak F, Singh SP, Margolis L, Zimmerberg J, Roberts DD. CD63+ and MHC Class I+ Subsets of Extracellular Vesicles Produced by Wild-Type and CD47-Deficient Jurkat T Cells Have Divergent Functional Effects on Endothelial Cell Gene Expression. Biomedicines. 2021; 9(11):1705. https://doi.org/10.3390/biomedicines9111705
Chicago/Turabian StyleKaur, Sukhbir, Abdel G. Elkahloun, Jennifer D. Petersen, Anush Arakelyan, Ferenc Livak, Satya P. Singh, Leonid Margolis, Joshua Zimmerberg, and David D. Roberts. 2021. "CD63+ and MHC Class I+ Subsets of Extracellular Vesicles Produced by Wild-Type and CD47-Deficient Jurkat T Cells Have Divergent Functional Effects on Endothelial Cell Gene Expression" Biomedicines 9, no. 11: 1705. https://doi.org/10.3390/biomedicines9111705
APA StyleKaur, S., Elkahloun, A. G., Petersen, J. D., Arakelyan, A., Livak, F., Singh, S. P., Margolis, L., Zimmerberg, J., & Roberts, D. D. (2021). CD63+ and MHC Class I+ Subsets of Extracellular Vesicles Produced by Wild-Type and CD47-Deficient Jurkat T Cells Have Divergent Functional Effects on Endothelial Cell Gene Expression. Biomedicines, 9(11), 1705. https://doi.org/10.3390/biomedicines9111705