
The Role of Omics Approaches to Characterize Molecular Mechanisms of Rare Ovarian Cancers: Recent Advances and Future Perspectives

 ,
,  ,
, 
Abstract
1. Introduction
2. Ovarian Cancer
2.1. Epithelial Ovarian Cancers
2.1.1. Serous Carcinomas
2.1.2. Ovarian Clear Cell Cancer
2.1.3. Mucinous Epithelial Ovarian Cancer
2.2. Non-Epithelial Ovarian Cancers
2.2.1. Malignant Germ Cell Tumors
2.2.2. Granulosa Cell Tumors
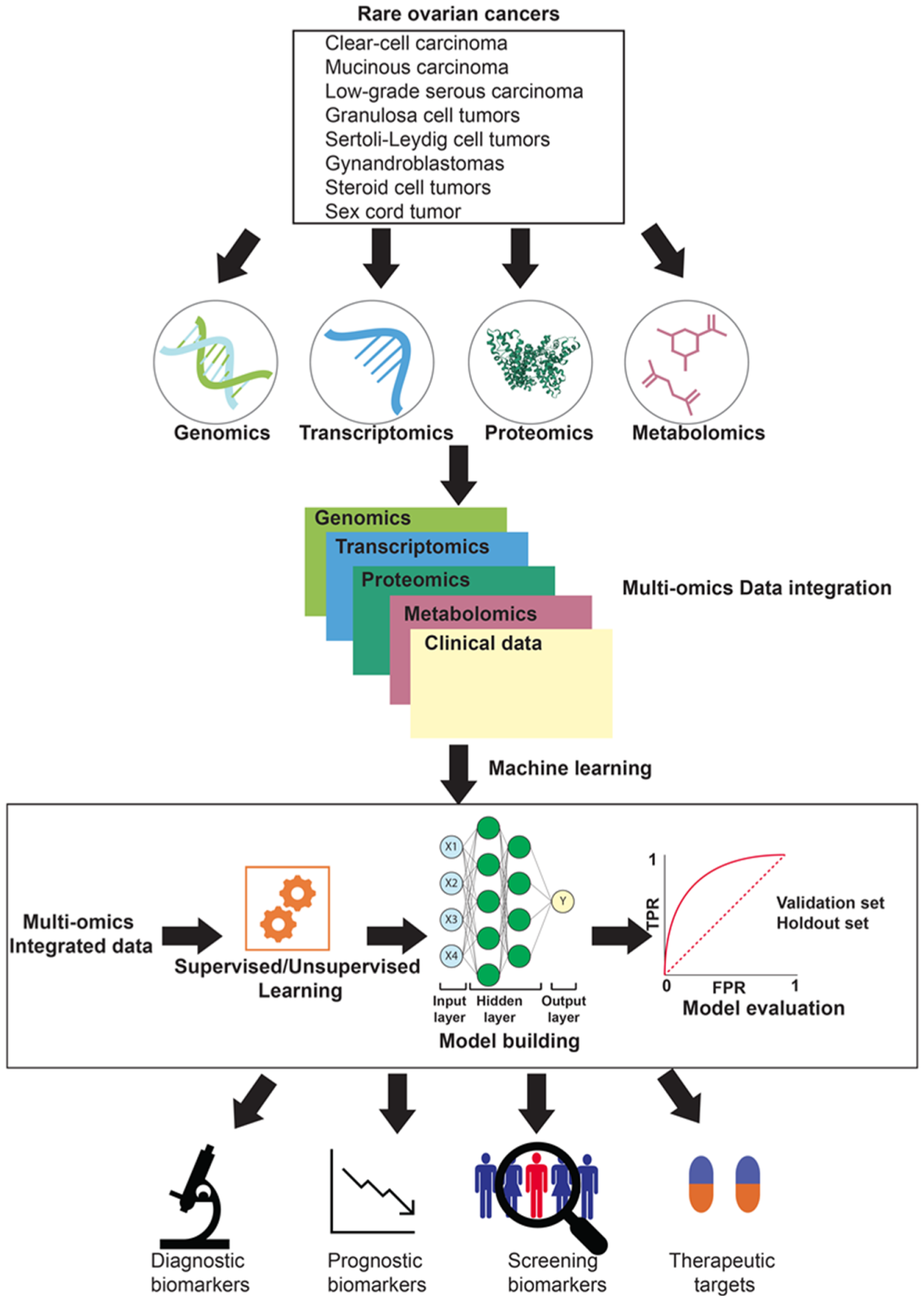
3. “Omics” Approaches and Integrative Biology
3.1. Genomics Approaches for the Identification of Alterations in the Rare Ovarian Cancer Genome
3.2. Exploration of the Rare Ovarian Cancer Transcriptome
3.3. Interrogation of the Proteomic Landscape of Rare Ovarian Cancers Using Prote-Omics
3.4. Exploration of the Rare Ovarian Cancer Metabolome
4. Multi-Omics Dataset Integration towards a Systems Biology View of Rare Ovarian Cancers
5. Potential Applications of Omics Approaches to Study Cancer Recurrence and Drug Resistance
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EOCs | Epithelial ovarian cancers |
| GCT | Granulosa cell tumors |
| GWAS | Genome-wide association study |
| HGSOC | High-grade serous ovarian carcinoma |
| LC-MS/MS | Liquid chromatography tandem mass spectrometry |
| LGSOC | Low-grade serous ovarian carcinoma |
| LncRNA | Long non-coding RNA |
| mEOC | Mucinous epithelial ovarian cancer |
| miRNA/miR | Micro-RNA |
| ncRNA | Non-coding RNA |
| NGS | Next-generation sequencing |
| OC | Ovarian cancer |
| OCCC | Ovarian clear cell cancer/carcinoma |
| OS | Overall survival |
| PCR | Polymerase chain reaction |
| PFS | Progression-free survival |
| RGCs | Rare gynecological cancers |
| RNA | Ribonucleic Acid |
| RNA-Seq | RNA sequencing |
| TKIs | Tyrosine kinase inhibitors |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2017. CA A Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef]
- Li, J.-Y.; Li, C.-J.; Lin, L.-T.; Tsui, K.-H. Multi-Omics Analysis Identifying Key Biomarkers in Ovarian Cancer. Cancer Control. 2020, 27, 1073274820976671. [Google Scholar] [CrossRef]
- Ray-Coquard, I.; Trama, A.; Seckl, M.; Fotopoulou, C.; Pautier, P.; Pignata, S.; Kristensen, G.; Mangili, G.; Falconer, H.; Massuger, L.; et al. Rare ovarian tumours: Epidemiology, treatment challenges in and outside a network setting. Eur. J. Surg. Oncol. 2019, 45, 67–74. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization; International Agency for Research on Cancer (IARC); WHO Classification of Tumours Editorial Board. WHO Classification of Tumours: Female Genital Tumours, 5th ed.; IARC: Lyon, France, 2020; 632p. [Google Scholar]
- Di Fiore, R.; Suleiman, S.; Ellul, B.; O’Toole, S.; Savona-Ventura, C.; Felix, A.; Napolioni, V.; Conlon, N.; Erson-Bensan, A.; Kahramanoglu, I.; et al. GYNOCARE Update: Modern Strategies to Improve Diagnosis and Treatment of Rare Gynecologic Tumors—Current Challenges and Future Directions. Cancers 2021, 13, 493. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Asmis, R.; Hawkins, G.A.; Howard, T.D.; Cox, L.A. The Need for Multi-Omics Biomarker Signatures in Precision Medicine. Int. J. Mol. Sci. 2019, 20, 4781. [Google Scholar] [CrossRef]
- Lu, M.; Zhan, X. The crucial role of multiomic approach in cancer research and clinically relevant outcomes. EPMA J. 2018, 9, 77–102. [Google Scholar] [CrossRef] [PubMed]
- Menyhárt, O.; Győrffy, B. Multi-omics approaches in cancer research with applications in tumor subtyping, prognosis, and diagnosis. Comput. Struct. Biotechnol. J. 2021, 19, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Hosen, I.; Ahmed, M.; Shekhar, H.U. Onco-Multi-OMICS Approach: A New Frontier in Cancer Research. BioMed Res. Int. 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Nicora, G.; Vitali, F.; Dagliati, A.; Geifman, N.; Bellazzi, R. Integrated Multi-Omics Analyses in Oncology: A Review of Machine Learning Methods and Tools. Front. Oncol. 2020, 10, 1030. [Google Scholar] [CrossRef]
- Yanai, H. Pathology of Epithelial Ovarian Tumors. In Frontiers in Ovarian Cancer Science; Katabuchi, H., Ed.; Springer: Singapore, 2017; pp. 83–113. [Google Scholar]
- Kurman, R.J.; Shih, I.-M. Molecular pathogenesis and extraovarian origin of epithelial ovarian cancer—Shifting the paradigm. Hum. Pathol. 2011, 42, 918–931. [Google Scholar] [CrossRef]
- Fung-Kee-Fung, M.; Oliver, T.; Elit, L.; Oza, A.; Hirte, H.W.; Bryson, P.; on behalf of the Gynecology Cancer Disease Site Group of Cancer Care Ontario’s Program in Evidence-Based Care. Optimal Chemotherapy Treatment for Women with Recurrent Ovarian Cancer. Curr. Oncol. 2007, 14, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Amante, S.; Santos, F.; Cunha, T.M. Low-grade serous epithelial ovarian cancer: A comprehensive review and update for radiologists. Insights Imaging 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mandilaras, V.; Karakasis, K.; Clarke, B.; Oza, A.; Lheureux, S. Rare tumors in gynaecological cancers and the lack of therapeutic options and clinical trials. Expert Opin. Orphan Drugs 2016, 5, 71–83. [Google Scholar] [CrossRef]
- Zorn, K.; Bonome, T.; Gangi, L.; Chandramouli, G.V.; Awtrey, C.S.; Gardner, G.J.; Barrett, J.C.; Boyd, J.; Birrer, M.J. Gene Expression Profiles of Serous, Endometrioid, and Clear Cell Subtypes of Ovarian and Endometrial Cancer. Clin. Cancer Res. 2005, 11, 6422–6430. [Google Scholar] [CrossRef]
- Yoshida, S.; Furukawa, N.; Haruta, S.; Tanase, Y.; Kanayama, S.; Noguchi, T.; Sakata, M.; Yamada, Y.; Oi, H.; Kobayashi, H. Theoretical model of treatment strategies for clear cell carcinoma of the ovary: Focus on perspectives. Cancer Treat. Rev. 2009, 35, 608–615. [Google Scholar] [CrossRef]
- Shibuya, Y.; Tokunaga, H.; Saito, S.; Shimokawa, K.; Katsuoka, F.; Bin, L.; Kojima, K.; Nagasaki, M.; Yamamoto, M.; Yaegashi, N.; et al. Identification of somatic genetic alterations in ovarian clear cell carcinoma with next generation sequencing. Genes Chromosom. Cancer 2018, 57, 51–60. [Google Scholar] [CrossRef]
- Bitler, B.; Aird, K.M.; Garipov, A.; Li, H.; Amatangelo, M.; Kossenkov, A.V.; Schultz, D.C.; Liu, Q.; Shih, I.-M.; Conejo-Garcia, J.; et al. Synthetic lethality by targeting EZH2 methyltransferase activity in ARID1A-mutated cancers. Nat. Med. 2015, 21, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Bitler, B.; Wu, S.; Park, P.H.; Hai, Y.; Aird, K.M.; Wang, Y.; Zhai, Y.; Kossenkov, A.V.; Vara-Ailor, A.; Iii, F.J.R.; et al. ARID1A-mutated ovarian cancers depend on HDAC6 activity. Nat. Cell Biol. 2017, 19, 962–973. [Google Scholar] [CrossRef]
- Miller, R.E.; Brough, R.; Bajrami, I.; Williamson, C.T.; McDade, S.; Campbell, J.; Kigozi, A.; Rafiq, R.; Pemberton, H.; Natrajan, R.; et al. Synthetic Lethal Targeting of ARID1A-Mutant Ovarian Clear Cell Tumors with Dasatinib. Mol. Cancer Ther. 2016, 15, 1472–1484. [Google Scholar] [CrossRef]
- Berns, K.; Caumanns, J.J.; Hijmans, E.M.; Gennissen, A.M.C.; Severson, T.M.; Evers, B.; Wisman, G.B.A.; Meersma, G.J.; Lieftink, C.; Beijersbergen, R.; et al. ARID1A mutation sensitizes most ovarian clear cell carcinomas to BET inhibitors. Oncogene 2018, 37, 4611–4625. [Google Scholar] [CrossRef]
- Fujimura, M.; Hidaka, T.; Saito, S. Selective inhibition of the epidermal growth factor receptor by ZD1839 decreases the growth and invasion of ovarian clear cell adenocarcinoma cells. Clin. Cancer Res. 2002, 8, 2448–2454. [Google Scholar] [PubMed]
- Mabuchi, S.; Kawase, C.; Altomare, D.A.; Morishige, K.; Sawada, K.; Hayashi, M.; Tsujimoto, M.; Yamoto, M.; Klein-Szanto, A.J.; Schilder, R.J.; et al. mTOR Is a Promising Therapeutic Target Both in Cisplatin-Sensitive and Cisplatin-Resistant Clear Cell Carcinoma of the Ovary. Clin. Cancer Res. 2009, 15, 5404–5413. [Google Scholar] [CrossRef]
- Anglesio, M.S.; Kommoss, S.; Tolcher, M.C.; Clarke, B.; Galletta, L.; Porter, H.; Damaraju, S.; Fereday, S.; Winterhoff, B.J.; Kalloger, S.E.; et al. Molecular characterization of mucinous ovarian tumours supports a stratified treatment approach with HER2 targeting in 19% of carcinomas. J. Pathol. 2012, 229, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, M. Pathology of Non-epithelial Ovarian Tumors. In Frontiers in Ovarian Cancer Science; Katabuchi, H., Ed.; Springer: Singapore, 2017; pp. 115–141. [Google Scholar]
- Bower, M.; Fife, K.; Holden, L.; Paradinas, F.; Rustin, G.; Newlands, E. Chemotherapy for ovarian germ cell tumours. Eur. J. Cancer 1996, 32, 593–597. [Google Scholar] [CrossRef]
- Simone, C.G.; Markham, M.J.; Dizon, D.S. Chemotherapy in ovarian germ cell tumors: A systematic review. Gynecol. Oncol. 2016, 141, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Goulvent, T.; Ray-Coquard, I.; Borel, S.; Haddad, V.; Devouassoux-Shisheboran, M.; Vacher-Lavenu, M.-C.; Pujade-Laurraine, E.; Savina, A.; Maillet, D.; Gillet, G.; et al. DICER1andFOXL2mutations in ovarian sex cord-stromal tumours: A GINECO Group study. Histopathology 2016, 68, 279–285. [Google Scholar] [CrossRef]
- Ray-Coquard, I.; Weber, B.; Lotz, J.P.; Tournigand, C.; Provençal, J.; Mayeur, D.; Treilleux, I.; Paraiso, D.; Duvillard, P.; Pujade-Lauraine, É. Management of rare ovarian cancers: The experience of the French website «Observatory for rare malignant tumours of the ovaries» by the GINECO group: Interim analysis of the first 100 patients. Gynecol. Oncol. 2010, 119, 53–59. [Google Scholar] [CrossRef]
- Manchana, T.; Ittiwut, C.; Mutirangura, A.; Kavanagh, J.J. Targeted therapies for rare gynaecological cancers. Lancet Oncol. 2010, 11, 685–693. [Google Scholar] [CrossRef]
- Kommoss, S.; Anglesio, M.S.; Mackenzie, R.; Yang, W.; Senz, J.; Ho, J.; Bell, L.; Lee, S.; Lorette, J.; Huntsman, D.G.; et al. FOXL2 molecular testing in ovarian neoplasms: Diagnostic approach and procedural guidelines. Mod. Pathol. 2013, 26, 860–867. [Google Scholar] [CrossRef]
- Jamieson, S.; Fuller, P. Molecular Pathogenesis of Granulosa Cell Tumors of the Ovary. Endocr. Rev. 2012, 33, 109–144. [Google Scholar] [CrossRef]
- Itamochi, H.; Oishi, T.; Oumi, N.; Takeuchi, S.; Yoshihara, K.; Mikami, M.; Yaegashi, N.; Terao, Y.; Takehara, K.; Ushijima, K.; et al. Whole-genome sequencing revealed novel prognostic biomarkers and promising targets for therapy of ovarian clear cell carcinoma. Br. J. Cancer 2017, 117, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Ryland, G.L.; Australian Ovarian Cancer Study Group; Hunter, S.M.; Doyle, M.A.; Caramia, F.; Li, J.; Rowley, S.; Christie, M.; Allan, P.E.; Stephens, A.N.; et al. Mutational landscape of mucinous ovarian carcinoma and its neoplastic precursors. Genome Med. 2015, 7, 1–12. [Google Scholar] [CrossRef]
- Cheasley, D.; Nigam, A.; Zethoven, M.; Hunter, S.; Etemadmoghadam, D.; Semple, T.; Allan, P.; Carey, M.S.; Fernandez, M.L.; Dawson, A.; et al. Genomic analysis of low-grade serous ovarian carcinoma to identify key drivers and therapeutic vulnerabilities. J. Pathol. 2021, 253, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, S.; Ikeda, K.; Horie-Inoue, K.; Sato, S.; Itakura, A.; Takeda, S.; Hasegawa, K.; Inoue, S. Systematic Identification of Characteristic Genes of Ovarian Clear Cell Carcinoma Compared with High-Grade Serous Carcinoma Based on RNA-Sequencing. Int. J. Mol. Sci. 2019, 20, 4330. [Google Scholar] [CrossRef]
- Alexiadis, M.; Chu, S.; Leung, D.; Gould, J.A.; Jobling, T.; Fuller, P.J. Transcriptomic analysis of stage 1 versus advanced adult granulosa cell tumors. Oncotarget 2016, 7, 14207–14219. [Google Scholar] [CrossRef]
- Shih, A.J.; Menzin, A.; Whyte, J.; Lovecchio, J.; Liew, A.; Khalili, H.; Bhuiya, T.; Gregersen, P.K.; Lee, A.T. Identification of grade and origin specific cell populations in serous epithelial ovarian cancer by single cell RNA-seq. PLoS ONE 2018, 13, e0206785. [Google Scholar] [CrossRef]
- Morita, A.; Miyagi, E.; Yasumitsu, H.; Kawasaki, H.; Hirano, H.; Hirahara, F. Proteomic search for potential diagnostic markers and therapeutic targets for ovarian clear cell adenocarcinoma. Proteomics 2006, 6, 5880–5890. [Google Scholar] [CrossRef]
- Fata, C.R.; Seeley, E.; Desouki, M.M.; Du, L.; Gwin, K.; Hanley, K.Z.; Hecht, J.L.; Jarboe, E.A.; Liang, S.X.; Parkash, V.; et al. Are clear cell carcinomas of the ovary and endometrium phenotypically identical? A proteomic analysis. Hum. Pathol. 2015, 46, 1427–1436. [Google Scholar] [CrossRef]
- Faratian, D.; Um, I.; Wilson, D.S.; Mullen, P.; Langdon, S.P.; Harrison, D.J. Phosphoprotein pathway profiling of ovarian carcinoma for the identification of potential new targets for therapy. Eur. J. Cancer 2011, 47, 1420–1431. [Google Scholar] [CrossRef]
- Garg, G.; Yilmaz, A.; Kumar, P.; Turkoglu, O.; Mutch, D.G.; Powell, M.A.; Rosen, B.; Bahado-Singh, R.O.; Graham, S.F. Targeted metabolomic profiling of low and high grade serous epithelial ovarian cancer tissues: A pilot study. Metabolomics 2018, 14, 154. [Google Scholar] [CrossRef] [PubMed]
- Buas, M.F.; Gu, H.; Djukovic, D.; Zhu, J.; Drescher, C.W.; Urban, N.; Raftery, D.; Li, C.I. Identification of novel candidate plasma metabolite biomarkers for distinguishing serous ovarian carcinoma and benign serous ovarian tumors. Gynecol. Oncol. 2016, 140, 138–144. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef]
- Bitler, B.G.; Watson, Z.; Wheeler, L.J.; Behbakht, K. PARP inhibitors: Clinical utility and possibilities of overcoming resistance. Gynecol. Oncol. 2017, 147, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Wang, T.-L.; Shih, I.-M.; Mao, T.-L.; Nakayama, K.; Roden, R.; Glas, R.; Slamon, D.; Diaz, L.A., Jr.; Vogelstein, B.; et al. Frequent Mutations of Chromatin Remodeling Gene ARID1A in Ovarian Clear Cell Carcinoma. Science 2010, 330, 228–231. [Google Scholar] [CrossRef]
- Mackenzie, R.; Kommoss, S.; Winterhoff, B.J.; Kipp, B.R.; Garcia, J.J.; Voss, J.; Halling, K.; Karnezis, A.; Senz, J.; Yang, W.; et al. Targeted deep sequencing of mucinous ovarian tumors reveals multiple overlapping RAS-pathway activating mutations in borderline and cancerous neoplasms. BMC Cancer 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cheasley, D.; Wakefield, M.J.; Ryland, G.L.; Allan, P.E.; Alsop, K.; Amarasinghe, K.C.; Ananda, S.; Anglesio, M.S.; Au-Yeung, G.; Böhm, M.; et al. The molecular origin and taxonomy of mucinous ovarian carcinoma. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Hunter, S.M.; Anglesio, M.S.; Ryland, G.L.; Sharma, R.; Chiew, Y.-E.; Rowley, S.M.; Doyle, M.A.; Li, J.; Gilks, C.B.; Moss, P.; et al. Molecular profiling of low grade serous ovarian tumours identifies novel candidate driver genes. Oncotarget 2015, 6, 37663–37677. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Winterhoff, B.J.; Kalli, K.R.; Block, M.S.; Armasu, S.M.; Larson, M.C.; Chen, H.-W.; Keeney, G.L.; Hartmann, L.C.; Shridhar, V.; et al. Expression signature distinguishing two tumour transcriptome classes associated with progression-free survival among rare histological types of epithelial ovarian cancer. Br. J. Cancer 2016, 114, 1412–1420. [Google Scholar] [CrossRef] [PubMed]
- Alldredge, J.; Randall, L.; De Robles, G.; Agrawal, A.; Mercola, D.; Liu, M.; Randhawa, P.; Edwards, R.; McClelland, M.; Rahmatpanah, F. Transcriptome Analysis of Ovarian and Uterine Clear Cell Malignancies. Front. Oncol. 2020, 10, 598579. [Google Scholar] [CrossRef]
- Sato, M.; Kawana, K.; Adachi, K.; Fujimoto, A.; Yoshida, M.; Nakamura, H.; Nishida, H.; Inoue, T.; Taguchi, A.; Ogishima, J.; et al. Targeting glutamine metabolism and the focal adhesion kinase additively inhibits the mammalian target of the rapamycin pathway in spheroid cancer stem-like properties of ovarian clear cell carcinoma in vitro. Int. J. Oncol. 2017, 50, 1431–1438. [Google Scholar] [CrossRef]
- Earp, M.A.; Raghavan, R.; Li, Q.; Dai, J.; Winham, S.J.; Cunningham, J.M.; Natanzon, Y.; Kalli, K.R.; Hou, X.; Weroha, S.J.; et al. Characterization of fusion genes in common and rare epithelial ovarian cancer histologic subtypes. Oncotarget 2017, 8, 46891–46899. [Google Scholar] [CrossRef]
- Benayoun, B.; Anttonen, M.; L’Hôte, D.; Bailly-Bechet, M.; Andersson, N.; Heikinheimo, M.; Veitia, R.A. Adult ovarian granulosa cell tumor transcriptomics: Prevalence of FOXL2 target genes misregulation gives insights into the pathogenic mechanism of the p.Cys134Trp somatic mutation. Oncogene 2012, 32, 2739–2746. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Auguste, A.; Bessière, L.; Todeschini, A.-L.; Caburet, S.; Sarnacki, S.; Prat, J.; D’Angelo, E.; De La Grange, P.; Ariste, O.; Lemoine, F.; et al. Molecular analyses of juvenile granulosa cell tumors bearingAKT1mutations provide insights into tumor biology and therapeutic leads. Hum. Mol. Genet. 2015, 24, 6687–6698. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Cheung, H.H.; Lu, G.; Chen, Z.; Chan, W.-Y. MicroRNA-10a promotes granulosa cells tumor development via PTEN-AKT/Wnt regulatory axis. Cell Death Dis. 2018, 9, 1076. [Google Scholar] [CrossRef]
- Fan, J.; Slowikowski, K.; Zhang, F. Single-cell transcriptomics in cancer: Computational challenges and opportunities. Exp. Mol. Med. 2020, 52, 1452–1465. [Google Scholar] [CrossRef]
- Pandey, A.; Mann, M. Proteomics to study genes and genomes. Nat. Cell Biol. 2000, 405, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, P.R.; Srivastava, S.; Hanash, S.; Wright, G.L. Proteomics in Early Detection of Cancer. Clin. Chem. 2001, 47, 1901–1911. [Google Scholar] [CrossRef]
- Subbannayya, Y.; Syed, N.; Barbhuiya, M.; Raja, R.; Marimuthu, A.; Sahasrabuddhe, N.; Pinto, S.; Manda, S.S.; Renuse, S.; Manju, H.; et al. Calcium calmodulin dependent kinase kinase 2—A novel therapeutic target for gastric adenocarcinoma. Cancer Biol. Ther. 2015, 16, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Kreunin, P.; Zhao, J.; Rosser, C.; Urquidi, V.; Lubman, D.M.; Goodison, S. Bladder Cancer Associated Glycoprotein Signatures Revealed by Urinary Proteomic Profiling. J. Proteome Res. 2007, 6, 2631–2639. [Google Scholar] [CrossRef]
- Subbannayya, Y.; Mir, S.A.; Renuse, S.; Manda, S.S.; Pinto, S.M.; Puttamallesh, V.N.; Solanki, H.S.; Manju, H.; Syed, N.; Sharma, R.; et al. Identification of differentially expressed serum proteins in gastric adenocarcinoma. J. Proteom. 2015, 127, 80–88. [Google Scholar] [CrossRef]
- Chen, H.; Pimienta, G.; Gu, Y.; Sun, X.; Hu, J.; Kim, M.-S.; Chaerkady, R.; Gucek, M.; Cole, R.N.; Sukumar, S.; et al. Proteomic characterization of Her2/neu-overexpressing breast cancer cells. Proteomics 2010, 10, 3800–3810. [Google Scholar] [CrossRef]
- Marimuthu, A.; Subbannayya, Y.; Sahasrabuddhe, N.A.; Balakrishnan, L.; Syed, N.; Sekhar, N.R.; Katte, T.V.; Pinto, S.; Srikanth, S.M.; Kumar, P.; et al. SILAC-based quantitative proteomic analysis of gastric cancer secretome. Proteom. Clin. Appl. 2013, 7, 355–366. [Google Scholar] [CrossRef]
- Hebert, J.D.; Myers, S.A.; Naba, A.; Abbruzzese, G.; Lamar, J.M.; Carr, S.A.; Hynes, R.O. Proteomic Profiling of the ECM of Xenograft Breast Cancer Metastases in Different Organs Reveals Distinct Metastatic Niches. Cancer Res. 2020, 80, 1475–1485. [Google Scholar] [CrossRef] [PubMed]
- Toyama, A.; Suzuki, A.; Shimada, T.; Aoki, C.; Aoki, Y.; Umino, Y.; Nakamura, Y.; Aoki, D.; Sato, T.-A. Proteomic characterization of ovarian cancers identifying annexin-A4, phosphoserine aminotransferase, cellular retinoic acid-binding protein 2, and serpin B5 as histology-specific biomarkers. Cancer Sci. 2012, 103, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Grunt, T.W.; Slany, A.; Semkova, M.; Colomer, R.; López-Rodríguez, M.L.; Wuczkowski, M.; Wagner, R.; Gerner, C.; Stübiger, G. Membrane disruption, but not metabolic rewiring, is the key mechanism of anticancer-action of FASN-inhibitors: A multi-omics analysis in ovarian cancer. Sci. Rep. 2020, 10, 1–28. [Google Scholar] [CrossRef]
- Arakawa, N.; Miyagi, E.; Nomura, A.; Morita, E.; Ino, Y.; Ohtake, N.; Miyagi, Y.; Hirahara, F.; Hirano, H. Secretome-Based Identification of TFPI2, A Novel Serum Biomarker for Detection of Ovarian Clear Cell Adenocarcinoma. J. Proteome Res. 2013, 12, 4340–4350. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, N.; Kobayashi, H.; Yonemoto, N.; Masuishi, Y.; Ino, Y.; Shigetomi, H.; Furukawa, N.; Ohtake, N.; Miyagi, Y.; Hirahara, F.; et al. Clinical Significance of Tissue Factor Pathway Inhibitor 2, a Serum Biomarker Candidate for Ovarian Clear Cell Carcinoma. PLoS ONE 2016, 11, e0165609. [Google Scholar] [CrossRef] [PubMed]
- Pinto, S.M.; Subbannayya, Y.; Prasad, T.S.K. Functional Proteomic Analysis to Characterize Signaling Crosstalk. Methods Mol. Biol. 2019, 1871, 197–224. [Google Scholar] [CrossRef]
- Kimura, A.; Arakawa, N.; Hirano, H. Mass Spectrometric Analysis of the Phosphorylation Levels of the SWI/SNF Chromatin Remodeling/Tumor Suppressor Proteins ARID1A and Brg1 in Ovarian Clear Cell Adenocarcinoma Cell Lines. J. Proteome Res. 2014, 13, 4959–4969. [Google Scholar] [CrossRef]
- Suh, D.-S.; Oh, H.K.; Kim, J.; Park, S.; Shin, E.; Lee, K.; Kim, Y.-H.; Bae, J. Identification and Validation of Differential Phosphorylation Sites of the Nuclear FOXL2 Protein as Potential Novel Biomarkers for Adult-Type Granulosa Cell Tumors. J. Proteome Res. 2015, 14, 2446–2456. [Google Scholar] [CrossRef]
- Sogabe, M.; Nozaki, H.; Tanaka, N.; Kubota, T.; Kaji, H.; Kuno, A.; Togayachi, A.; Gotoh, M.; Nakanishi, H.; Nakanishi, T.; et al. Novel Glycobiomarker for Ovarian Cancer That Detects Clear Cell Carcinoma. J. Proteome Res. 2014, 13, 1624–1635. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics--the link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Markley, J.L.; Brüschweiler, R.; Edison, A.; Eghbalnia, H.R.; Powers, R.; Raftery, D.; Wishart, D.S. The future of NMR-based metabolomics. Curr. Opin. Biotechnol. 2017, 43, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Zhang, W.; Yin, M.; Zhang, T.; Wu, X.; Zhang, H.; Sun, M.; Li, Z.; Hou, Y.; Zhou, X.; et al. Identification of metabolic biomarkers to diagnose epithelial ovarian cancer using a UPLC/QTOF/MS platform. Acta Oncol. 2012, 51, 473–479. [Google Scholar] [CrossRef]
- Wu, W.; Wang, Q.; Yin, F.; Yang, Z.; Zhang, W.; Gabra, H.; Li, L. Identification of proteomic and metabolic signatures associated with chemoresistance of human epithelial ovarian cancer. Int. J. Oncol. 2016, 49, 1651–1665. [Google Scholar] [CrossRef]
- Gottschalk, R.A.; Martins, A.J.; Sjoelund, V.H.; Angermann, B.R.; Lin, B.; Germain, R.N. Recent progress using systems biology approaches to better understand molecular mechanisms of immunity. Semin. Immunol. 2013, 25, 201–208. [Google Scholar] [CrossRef]
- Picard, M.; Scott-Boyer, M.-P.; Bodein, A.; Périn, O.; Droit, A. Integration strategies of multi-omics data for machine learning analysis. Comput. Struct. Biotechnol. J. 2021, 19, 3735–3746. [Google Scholar] [CrossRef]
- Duan, R.; Gao, L.; Gao, Y.; Hu, Y.; Xu, H.; Huang, M.; Song, K.; Wang, H.; Dong, Y.; Jiang, C.; et al. Evaluation and comparison of multi-omics data integration methods for cancer subtyping. PLoS Comput. Biol. 2021, 17, e1009224. [Google Scholar] [CrossRef]
- Subramanian, I.; Verma, S.; Kumar, S.; Jere, A.; Anamika, K. Multi-omics Data Integration, Interpretation, and Its Application. Bioinform. Biol. Insights 2020, 14, 1177932219899051. [Google Scholar] [CrossRef] [PubMed]
- Subbannayya, Y.; Pinto, S.M.; Gowda, H.; Prasad, T.S.K. Proteogenomics for understanding oncology: Recent advances and future prospects. Expert Rev. Proteom. 2016, 13, 297–308. [Google Scholar] [CrossRef]
- Johansson, H.J.; Consortia Oslo Breast Cancer Research Consortium (OSBREAC); Socciarelli, F.; Vacanti, N.M.; Haugen, M.H.; Zhu, Y.; Siavelis, I.; Fernandez, A.; Aure, M.R.; Sennblad, B.; et al. Breast cancer quantitative proteome and proteogenomic landscape. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Mertins, P.; Cptac, N.; Mani, D.R.; Ruggles, K.V.; Gillette, M.A.; Clauser, K.R.; Wang, P.; Wang, X.; Qiao, J.W.; Cao, S.; et al. Proteogenomics connects somatic mutations to signalling in breast cancer. Nat. Cell Biol. 2016, 534, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-B.; Karpova, A.; Gritsenko, M.A.; Kyle, J.E.; Cao, S.; Li, Y.; Rykunov, D.; Colaprico, A.; Rothstein, J.H.; Hong, R.; et al. Proteogenomic and metabolomic characterization of human glioblastoma. Cancer Cell 2021, 39, 509–528.e20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, T.; Zhang, Z.; Payne, S.; Zhang, B.; McDermott, J.; Zhou, J.-Y.; Petyuk, V.A.; Chen, L.; Ray, D.; et al. Integrated Proteogenomic Characterization of Human High-Grade Serous Ovarian Cancer. Cell 2016, 166, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, J.; Wang, X.; Zhu, J.; Liu, Q.; Shi, Z.; Chambers, M.C.; Zimmerman, L.J.; Shaddox, K.F.; Kim, S.; et al. Proteogenomic characterization of human colon and rectal cancer. Nature 2014, 513, 382–387. [Google Scholar] [CrossRef]
- Wiegand, K.C.; Hennessy, B.T.; Leung, S.; Wang, Y.; Ju, Z.; McGahren, M.; Kalloger, S.E.; Finlayson, S.; Stemke-Hale, K.; Lu, Y.; et al. A functional proteogenomic analysis of endometrioid and clear cell carcinomas using reverse phase protein array and mutation analysis: Protein expression is histotype-specific and loss of ARID1A/BAF250a is associated with AKT phosphorylation. BMC Cancer 2014, 14, 120. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, E.; Tabata, J.; Yanaihara, N.; Ishikawa, T.; Koseki, K.; Iida, Y.; Saito, M.; Komazaki, H.; Shapiro, J.S.; Goto, C.; et al. Application of Artificial Intelligence for Preoperative Diagnostic and Prognostic Prediction in Epithelial Ovarian Cancer Based on Blood Biomarkers. Clin. Cancer Res. 2019, 25, 3006–3015. [Google Scholar] [CrossRef]
- Elias, K.M.; Fendler, W.; Stawiski, K.; Fiascone, S.J.; Vitonis, A.F.; Berkowitz, R.S.; Frendl, G.; Konstantinopoulos, P.; Crum, C.P.; Kedzierska, M.; et al. Diagnostic potential for a serum miRNA neural network for detection of ovarian cancer. eLife 2017, 6, e28932. [Google Scholar] [CrossRef]
- Guo, L.-Y.; Wu, A.-H.; Wang, Y.-X.; Zhang, L.-P.; Chai, H.; Liang, X.-F. Deep learning-based ovarian cancer subtypes identification using multi-omics data. BioData Min. 2020, 13, 1–12. [Google Scholar] [CrossRef]
- Hay, C.; Courtney-Brooks, M.; Lefkowits, C.; Hagan, T.L.; Edwards, R.P.; Donovan, H.S. Symptom management in women with recurrent ovarian cancer: Do patients and clinicians agree on what symptoms are most important? Gynecol. Oncol. 2016, 143, 367–370. [Google Scholar] [CrossRef]
- Kwek, J.W.; Iyer, R.B. Recurrent Ovarian Cancer: Spectrum of Imaging Findings. Am. J. Roentgenol. 2006, 187, 99–104. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist. 2019, 2, 141–160. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Lord, C.J.; Ashworth, A. PARP inhibitors: Synthetic lethality in the clinic. Science 2017, 355, 1152–1158. [Google Scholar] [CrossRef]
- Kurman, R.J.; Shih, I.-M. The Dualistic Model of Ovarian Carcinogenesis: Revisited, revised, and expanded. Am. J. Pathol. 2016, 186, 733–747. [Google Scholar] [CrossRef]
- Patch, A.-M.; The Australian Ovarian Cancer Study Group; Christie, E.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; et al. Whole–genome characterization of chemoresistant ovarian cancer. Nature 2015, 521, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Barry, W.T.; Birrer, M.; Lee, J.-M.; Buckanovich, R.J.; Fleming, G.F.; Rimel, B.; Buss, M.K.; Nattam, S.; Hurteau, J.; et al. Combination cediranib and olaparib versus olaparib alone for women with recurrent platinum-sensitive ovarian cancer: A randomised phase 2 study. Lancet Oncol. 2014, 15, 1207–1214. [Google Scholar] [CrossRef]
- Pignata, S.; Cecere, S.C.; Du Bois, A.; Harter, P.; Heitz, F. Treatment of recurrent ovarian cancer. Ann. Oncol. 2017, 28, viii51–viii56. [Google Scholar] [CrossRef] [PubMed]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug Resistance in Cancer: An Overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef]
- Sokolenko, A.P.; Imyanitov, E.N. Molecular Diagnostics in Clinical Oncology. Front. Mol. Biosci. 2018, 5, 76. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Omics | Study Type | Ovarian Cancer Subtype | Findings | Ref |
|---|---|---|---|---|
| Genomics | Whole-genome sequencing | Ovarian clear cell carcinoma | prevalent mutations in ARID1A (42%), PIK3CA (35%), HLA-DRB1 (25%), MUC4 (22%), ZNF717 (22%), and ARID1B (18%) | [34] |
| Exome sequencing | Mucinous ovarian cancers | mutations in KRAS, BRAF, CDKN2A, TP53, RNF43, ELF3, GNAS, ERBB3, and KLF5 genes | [35] | |
| Targeted sequencing | Low-grade serous ovarian cancers | prevalent mutations in KRAS, BRAF, and NRAS | [36] | |
| Transcriptomics | RNA-Seq | Ovarian clear cell carcinoma | CPNE8 is a differential marker to distinguish between OCCC and HGSOC | [37] |
| Gene expression | Ovarian granulosa cell tumors | 24 genes differentially expressed between early and advanced granulosa cell tumors | [38] | |
| Single-cell RNA-seq | Serous epithelial ovarian tumors | sixteen distinct cell populations with specific cells correlated to high-grade, low-grade, and benign tumors | [39] | |
| Proteomics | 2D-gel electrophoresis and mass spectrometry | Clear cell adenocarcinoma and mucinous adenocarcinoma | distinguishing markers between clear cell adenocarcinoma and mucinous adenocarcinoma | [40] |
| matrix-assisted laser desorption/ionization (MALDI) mass spectrometry | Ovarian clear cell carcinoma | Differential proteins between ovarian clear cell carcinoma and endometrial clear cell carcinoma | [41] | |
| reverse-phase protein array (RPPA) | 4 histological subtypes of ovarian cancer | subtype-specific activation states of druggable oncogenic pathways | [42] | |
| Metabolomics | Nuclearmagnetic resonance (NMR) and mass spectrometry | EOCs | Differential markers between high-grade and low-grade ovarian cancers | [43] |
| Mass spectrometry | Serous ovarian cancers | Differential markers between serous ovarian cancers and benign tumors | [44] |
| Rare Ovarian Cancer Types | Rare Ovarian Cancer Subtypes | Characteristic Genomic Alterations |
|---|---|---|
| Epithelial cell tumors | Low-grade serous carcinoma | K/N-RAS (75%), BRAF (20%) |
| Mucinous carcinoma | KRAS (50%), TP53 (26%), MSI (20%), HER2-amp (20%) | |
| Clear cell carcinoma | ARI1D1A (55%), PI3K pathway (PTEN loss PI3KCAmut AKT2ampl), HER2 amp (14%), MSI (10%) | |
| Carcinosarcoma | TP53 (80%), PI3K pathway (PTEN loss, PI3K mutation) | |
| Germ-cell tumors | Dysgerminoma | KIT mutation (25%), High CNV 11p gain (60%) |
| Yolk sac tumor | DICER 1 (10%), High CNV | |
| Immature teratoma | Few CNV | |
| Other: choriocarcinoma, embryonal and mixed carcinoma | Few mutations | |
| Sex cord stromal tumors | Adult granulosa | FOXL-2 (missense mutt 402C ˃ G C, >95%, pathognomonic) ER+, PR+, AR+, AKT1/2, Pi3Kca, TGFb |
| Juvenile granulosa | GSP mutation (30%), AKT1 | |
| Sertoli-Leydig | DICER-1 (90%, including germline mutations), FGFR2 | |
| Small cell carcinoma hypercalcemic | SMARCA4 deficiency part of the SWI/SNF complex (90%), PDGFR1, FGFR1 | |
| Other sex cord stroma tumors | Poorly known |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subbannayya, Y.; Di Fiore, R.; Urru, S.A.M.; Calleja-Agius, J. The Role of Omics Approaches to Characterize Molecular Mechanisms of Rare Ovarian Cancers: Recent Advances and Future Perspectives. Biomedicines 2021, 9, 1481. https://doi.org/10.3390/biomedicines9101481
Subbannayya Y, Di Fiore R, Urru SAM, Calleja-Agius J. The Role of Omics Approaches to Characterize Molecular Mechanisms of Rare Ovarian Cancers: Recent Advances and Future Perspectives. Biomedicines. 2021; 9(10):1481. https://doi.org/10.3390/biomedicines9101481
Chicago/Turabian StyleSubbannayya, Yashwanth, Riccardo Di Fiore, Silvana Anna Maria Urru, and Jean Calleja-Agius. 2021. "The Role of Omics Approaches to Characterize Molecular Mechanisms of Rare Ovarian Cancers: Recent Advances and Future Perspectives" Biomedicines 9, no. 10: 1481. https://doi.org/10.3390/biomedicines9101481
APA StyleSubbannayya, Y., Di Fiore, R., Urru, S. A. M., & Calleja-Agius, J. (2021). The Role of Omics Approaches to Characterize Molecular Mechanisms of Rare Ovarian Cancers: Recent Advances and Future Perspectives. Biomedicines, 9(10), 1481. https://doi.org/10.3390/biomedicines9101481