
Antitumoral Activities of Curcumin and Recent Advances to ImProve Its Oral Bioavailability


 ,
, 
Abstract
1. Introduction
2. Antitumoral Activities of Curcumin
2.1. Antiproliferative Effects of Curcumin
2.2. Pro-Apoptotic Effects of Curcumin
2.3. Antimetastatic Effects of Curcumin
3. Bioavailability of Curcumin and Therapeutic Promises
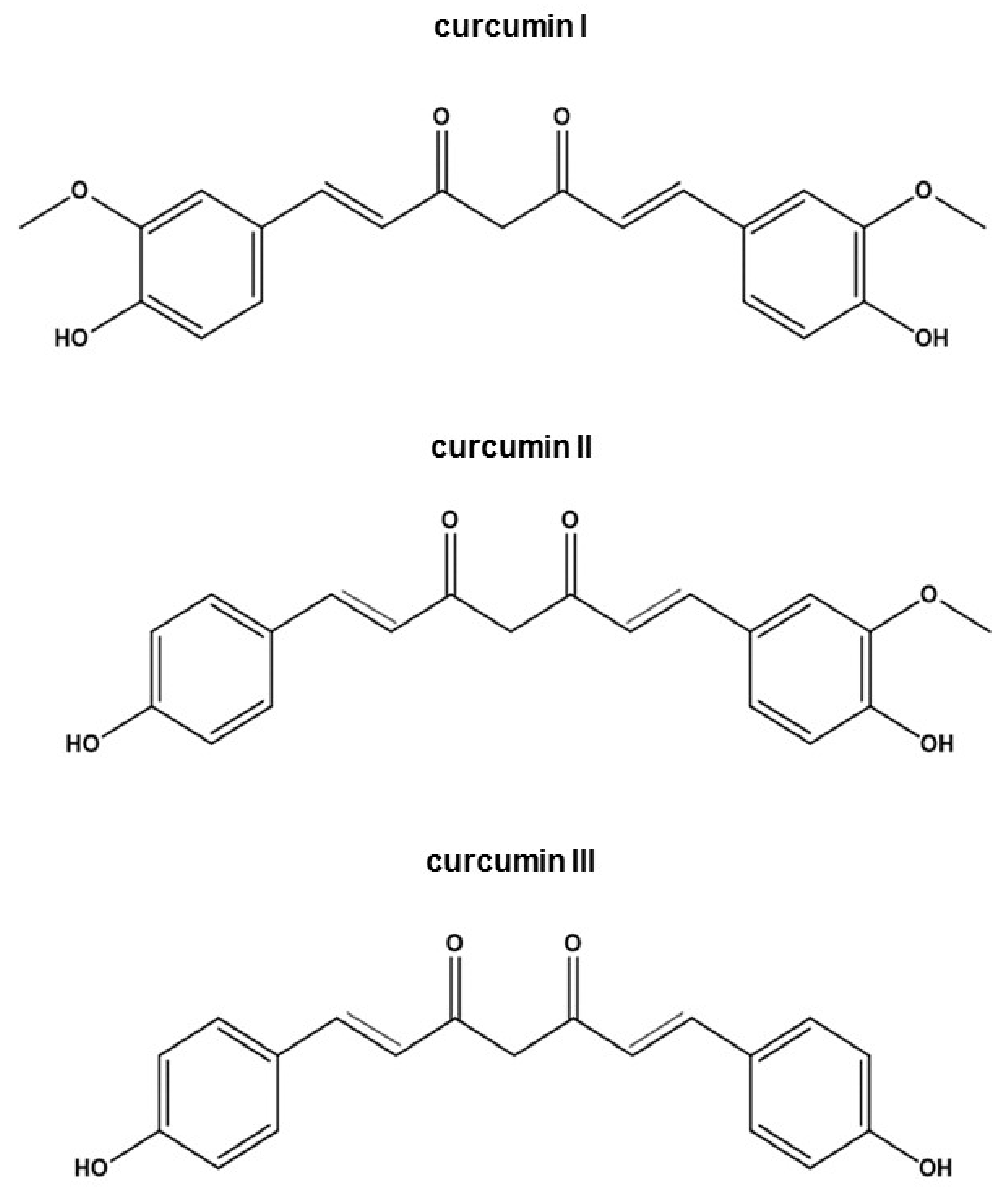
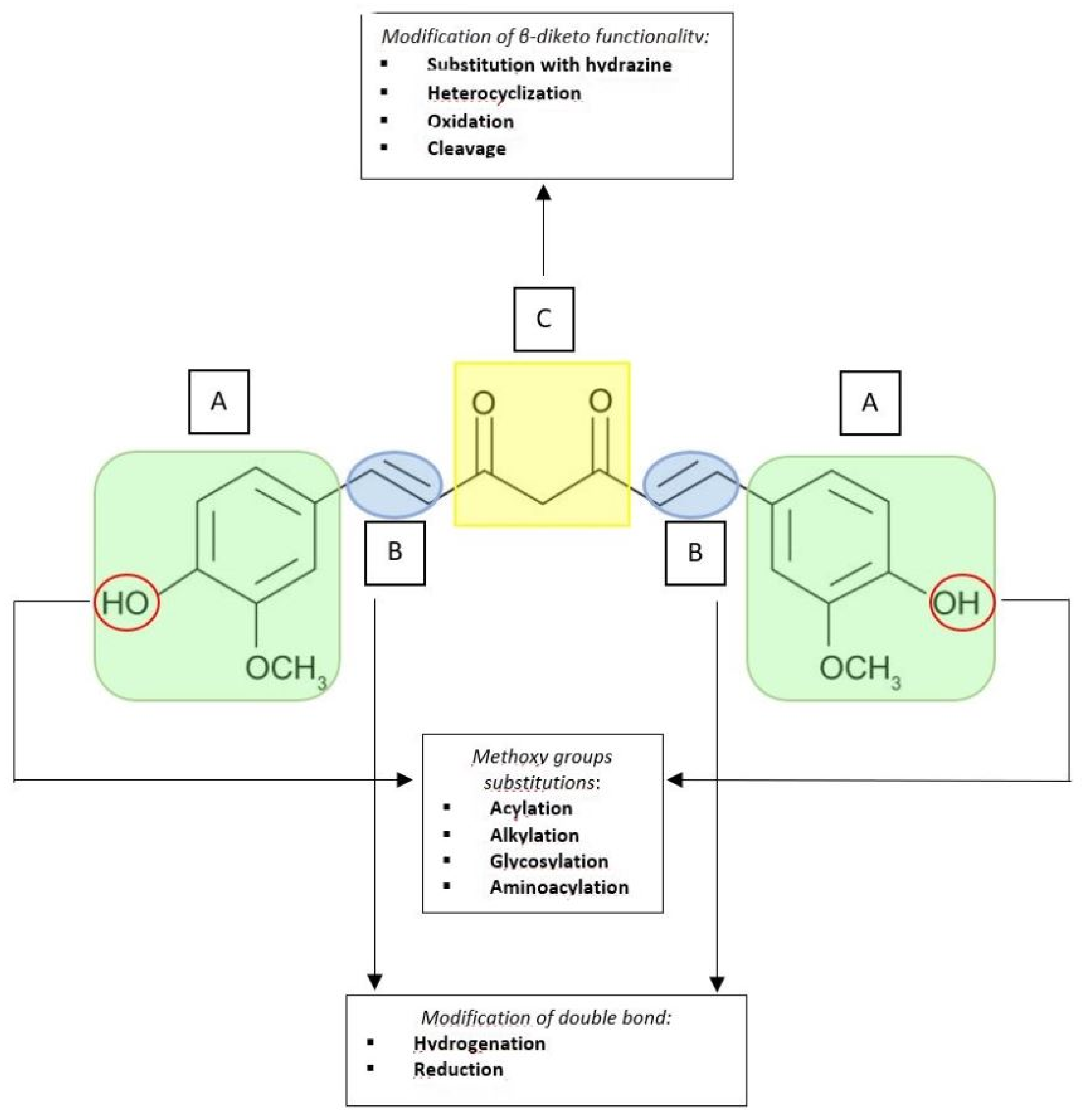
3.1. Curcumin Structural Derivatives and Analogues
3.2. Curcumin Delivery Systems
- (a).
- Delaying metabolism through its entrapment within the hydrophobic phases that isolate it from aqueous phase or cell membranes enzymes;
- (b).
- Improving its bioaccessibility through an increase in the quantity that is solubilized inside the mixed micelles present in the small intestine; this can be achieved by inserting surfactants, phospholipids, fatty acids or monoglycerides into the curcumin-loaded carrier particles;
- (c).
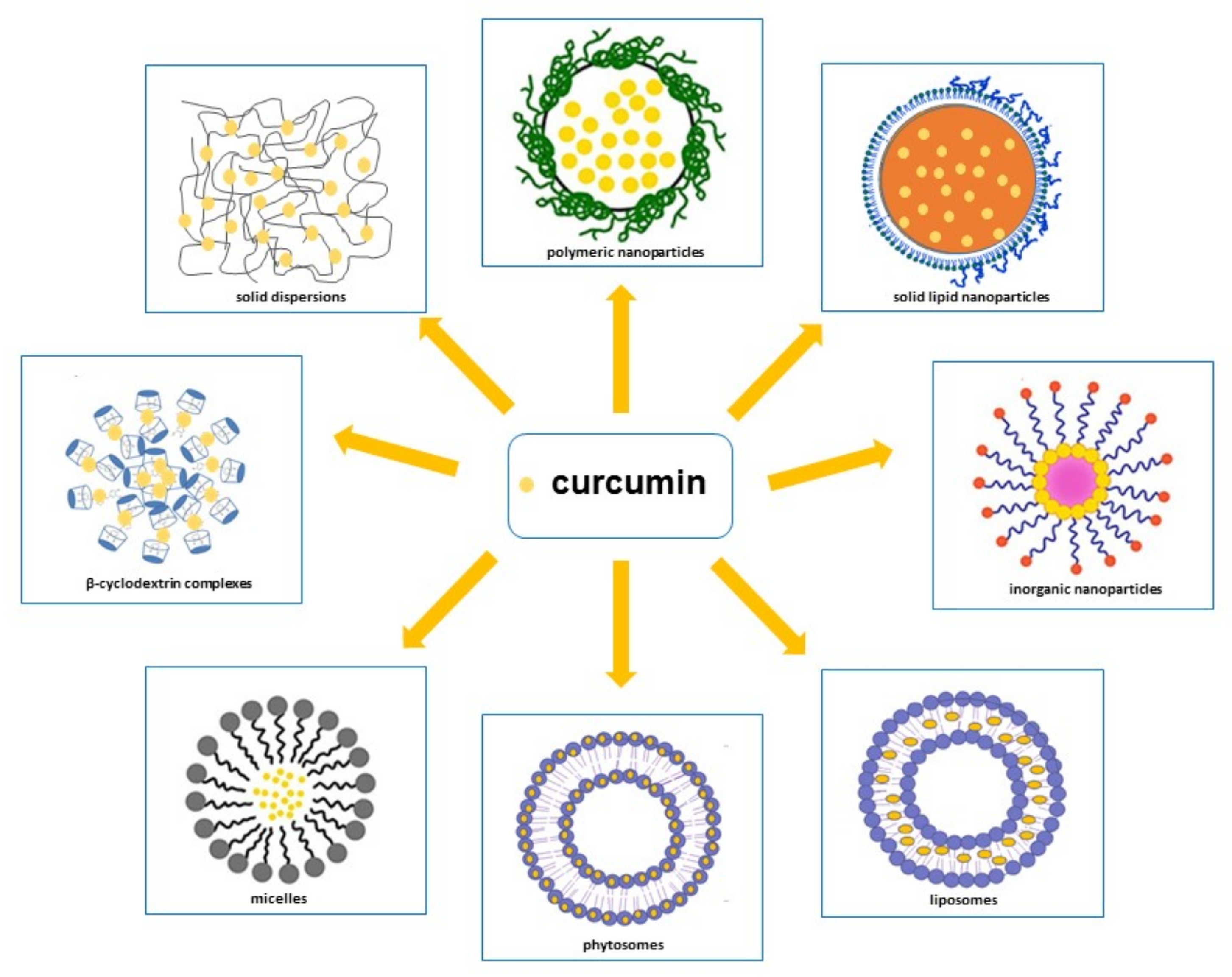
- Promoting its absorption by loading curcumin into particles carrier that contain substances able to increase epithelium cell membranes permeability or block efflux transporters [147]. Therefore, in order to ameliorate curcumin’s pharmacokinetic characteristics, various methodological approaches have been attempted, such as polymeric approaches, magnetic approaches, solid lipid nanoparticles, liposomes, phytosomes, micelles, β-cyclodextrins and solid dispersions [21,25,26,27,28,29,30,148,149,150] (Figure 3) [151,152,153,154]. In addition to these approaches, curcumin conjugation with substances, such as piperine, which is able to inhibit its metabolism [27,155], has emerged as a prominent solution to increase curcumin serum concentration.
3.2.1. Nanoparticles
Polymeric Nanoparticles
Solid Lipid Nanoparticles
Inorganic Nanoparticles
3.2.2. Liposomes
3.2.3. Phytosomes
3.2.4. Micelles
3.2.5. Curcumin/β-Cyclodextrin and Solid Dispersions Formulations
3.2.6. Curcumin Conjugates Formulations
4. In Situ Implant Systems
5. Clinical Trials with Curcumin
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AKT | protein kinase B |
| AMPK | adenosine monophosphate (AMP)-activated protein kinase |
| AP-1 | activator protein1 |
| APC | adenomatous polyposis coli |
| ARE | antioxidant response element |
| AUC | area under curve |
| AuNPs | gold nanoparticles |
| Bcl-2 | B-cell lymphoma 2 |
| Bcl-xL | B-cell lymphoma-extra large |
| cAMP | cyclic adenosine monophosphate |
| CDK2 | cyclin-dependent kinase 2 |
| CDK4 | cyclin-dependent kinase 4 |
| CD | cyclodextrin |
| cGMP | cyclic guanosin monophosphate |
| CNrasGEFs | cyclic nucleotide-ras guanine nucleotide exchange factors |
| COX2 | cyclooxygenase-2 |
| CPC | curcumin phospholipid complex |
| CUR | curcumin |
| CUR-CD-CS | curcumin-loaded cyclodextrin chitosan |
| CUR-MMs | curcumin mixed micelles |
| CUR-MPP-TPGS-MMs | curcumin-loaded methoxy poly(ethylene glycol)-poly(lactide) (mPEG-PLA)/D-α-tocopherol polyethylene glycol 1000 succinate (TPGS) mixed micelles |
| CUR-PSMNPs | curcumin pluronic stabilized Fe3O4 magnetic nanoparticles |
| CUR-SF NPs | curcumin silk fibroin nanoparticles |
| DAPs | diarylpentanoids |
| DDAB | dimethyl dioctadecyl ammonium bromide |
| DFMO | difluoro-methylornithine |
| DHC | dihydrocurcumin |
| DMPC | dimyristoyl phosphatidylcholine |
| DNMT1 | DNA methyltransferase 1 |
| E2F4 | E2F trascription factor 4 |
| EGFR | epidermal growth factor receptor |
| Egr-1 | early growth response 1 |
| EMT | epithelial mesenchymal transition |
| ERK | extracellulare signal-regulated kinase |
| EZH2 | enhancer of zeste homolog 2 |
| FA | folic acid |
| FA-CUR-NPs | folate receptor-targeted β-cyclodextrin nanoparticles |
| Fas | fas-associated protein with death domain |
| FAS | fatty acid synthase |
| FRs | folic receptors |
| GSH | glutathione |
| GSK3β | glycogen synthase kinase 3 beta |
| HA-CUR@AuNPs | hyaluronic acid curcumin gold nanoparticles |
| HDGF | hepatoma-derived growth factor |
| HER2 | human epidermal growth factor receptor 2 |
| HHC | hexahydrocurcumin |
| HIF-1α | hypoxia-inducible factor 1-alpha |
| HPH | high pressure homogenization |
| IAP-2 | inhibitor of apoptosis protein-2 |
| ICAM1 | intercellular adhesion molecule 1 |
| IC50 | half-maximal inhibitory concentration |
| IFN-g | interferon gamma |
| IkB | inhibitor of KB |
| IKK | IκB kinase |
| IL-6 | interleukin-6 |
| JAK2 | janus kinase 2 |
| JNK | c-jun-N-terminal kinase |
| LPA | lysophosphatidic acid |
| LRP6 | LDL (low density lipoprotein) receptor related protein 6 |
| MAPK | mitogen-activated protein kinase |
| Mcl-1 | myeloid cell leukemia 1 |
| MDM2 | mouse double minute 2 homolog |
| MePEG-b-PCL | methoxy poly (ethylene glycol) -block-polycaprolactone |
| miRNA | microRNA |
| MMPs | matrix metalloproteinases |
| MNPs | magnetic nanoparticles |
| MPEG-P [CL-co-PDO] | methoxy poly (ethylene glycol) -b-poly (ε-caprolactone-co-dioxanone) |
| mPEG–PCL | methoxy poly(ethylene glycol)–polycaprolactone |
| mTOR | mechanistic target of rapamycin |
| MUC1 | mucin 1 |
| NaCas | sodium caseinate |
| NaCas-Lac | sodium caseinate-lactose |
| NF-κB | nuclear factor-κB |
| NIPAAM | N-isopropylacrylamide |
| NPs | nanoparticles |
| NSCLC | non-small cell lung cancer |
| NVP | N-vinyl-2-pyrrolidone |
| ODC | ornithine decarboxylase |
| OHC | octahydrocurcumin |
| OPN | osteopontin |
| PAK1 | p21 (RAC1) activated kinase 1 |
| PARP-1 | poly (ADP-ribose) polymerase 1 |
| PCL | poly-(ε-caprolactone) |
| PCNA | proliferating cell nuclear antigen |
| PDE | cyclic nucleotide phosphodiesterase |
| PDE1A | phosphodiesterase 1A |
| PEG | polyethylene glycol |
| PEG-A | poly(ethylene glycol) monoacrylate |
| PGA | polyglycolide acid |
| P-gp | p-glycoprotein 1 |
| PI3K | phosphoinositide 3-kinase |
| PLA | polylactide |
| PLGA | polylactic-co-glycolic acid |
| PSMNPs | pluronic stabilized Fe3O4 magnetic nanoparticles |
| PSBMA | poly (sulphobetaine methacrylate) |
| RhoA | ras homolog family member A |
| ROCK1 | rho associated coiled-coil containing protein kinase 1 |
| ROS | reactive oxygen species |
| SCLC | small cell lung cancer |
| SDs | solid dispersions |
| SIP1 | smad interacting protein 1 |
| SLNs | solid lipid nanoparticles |
| SMOX | spermine oxidase |
| STAT3 | signal transducer and activator of transcription 3 |
| SULT | sulfotransferase |
| THC | tetrahydrocurcumin |
| TNF-α | tumor necrosis factor-a |
| TPGS | d-α-Tocopheryl polyethylene glycol 1000 succinate |
| UGT | UDP-glucurosyltransferase |
| UHRF1 | ubiquitin like with PHD and ring finger domain 1 |
| u-PA | urokinase-type plasminogen activator |
| VEGF | vascular endothelial growth factor |
| VHL | Von Hippel-Lindau tumor suppressor |
| VP | N-vinyl-2-pyrrolidone |
| XIAP | X-linked inhibitor of apoptosis protein |
| β-CD | β- cyclodextrins |
| εCL | ε-caprolactone |
References
- Prasad, S.; Aggarwal, B.B. Turmeric, the golden spice: From traditional medicine to modern medicine. In Herbal Medicine: Biomolecular and Clinical Aspects; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Sandur, S.K.; Pandey, M.K.; Sung, B.; Ahn, K.S.; Murakami, A.; Sethi, G.; Limtrakul, P.; Badmaev, V.; Aggarwal, B.B. Curcumin, demethoxycurcumin, bisdemethoxycurcumin, tetrahydrocurcumin and turmerones differentially regulate anti-inflammatory and anti-proliferative responses through a ROS-independent mechanism. Carcinogenesis 2007, 28, 1765–1773. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Satyal, P.; Setzer, W.N. Variations in the volatile compositions of Curcuma species. Foods 2019, 8, 53. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: Multitargeting for multiple chronic diseases. Br. J. Pharm. 2017, 174, 1325–1348. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Acharya, A.; Ray, R.S.; Agrawal, R.; Raghuwanshi, R.; Jain, P. Cellular and molecular mechanisms of curcumin in prevention and treatment of disease. Crit. Rev. Food Sci. Nutr. 2020, 60, 887–939. [Google Scholar] [CrossRef] [PubMed]
- Adamczak, A.; Ozarowski, M.; Karpinski, T.M. Curcumin, a natural antimicrobial agent with strain-specific activity. Pharmaceuticals 2020, 13, 153. [Google Scholar] [CrossRef]
- Jennings, M.R.; Parks, R.J. Curcumin as an antiviral agent. Viruses 2020, 12, 1242. [Google Scholar] [CrossRef]
- Chen, C.; Long, L.; Zhang, F.; Chen, Q.; Chen, C.; Yu, X.; Liu, Q.; Bao, J.; Long, Z. Antifungal activity, main active components and mechanism of Curcuma longa extract against Fusarium graminearum. PLoS ONE 2018, 13, e0194284. [Google Scholar] [CrossRef]
- Jagetia, G.C. Antioxidant activity of curcumin protects against the radiation-induced micronuclei formation in cultured human peripheral blood lymphocytes exposed to various doses of gamma-Radiation. Int. J. Radiat. Biol. 2021, 97, 485–493. [Google Scholar] [CrossRef]
- Busari, Z.A.; Dauda, K.A.; Morenikeji, O.A.; Afolayan, F.; Oyeyemi, O.T.; Meena, J.; Sahu, D.; Panda, A.K. Antiplasmodial activity and toxicological assessment of curcumin PLGA-encapsulated nanoparticles. Front. Pharm. 2017, 8, 622. [Google Scholar] [CrossRef]
- Fadus, M.C.; Lau, C.; Bikhchandani, J.; Lynch, H.T. Curcumin: An age-old anti-inflammatory and anti-neoplastic agent. J. Tradit. Complement. Med. 2017, 7, 339–346. [Google Scholar] [CrossRef]
- Zia, A.; Farkhondeh, T.; Pourbagher-Shahri, A.M.; Samarghandian, S. The role of curcumin in aging and senescence: Molecular mechanisms. Biomed. Pharmacother. Biomed. Pharmacother. 2021, 134, 111119. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, K.; Liu, J.; Yang, J.; Tian, Y.; Yang, C.; Li, Y.; Shao, M.; Su, W.; Song, N. Curcumin regulates cancer progression: Focus on ncRNAs and molecular signaling pathways. Front. Oncol. 2021, 11, 660712. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Bordoloi, D.; Harsha, C.; Banik, K.; Gupta, S.C.; Aggarwal, B.B. Curcumin mediates anticancer effects by modulating multiple cell signaling pathways. Clin. Sci. 2017, 131, 1781–1799. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Park, K.S.; Kim, K.T.; Gil, E.Y. The inhibitory effect of curcumin via fascin suppression through JAK/STAT3 pathway on metastasis and recurrence of ovary cancer cells. BMC Women’s Health 2020, 20, 256. [Google Scholar] [CrossRef]
- Liu, J.L.; Pan, Y.Y.; Chen, O.; Luan, Y.; Xue, X.; Zhao, J.J.; Liu, L.; Jia, H.Y. Curcumin inhibits MCF-7 cells by modulating the NF-kappaB signaling pathway. Oncol. Lett. 2017, 14, 5581–5584. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, F.; Shafiee, M.; Banikazemi, Z.; Pourhanifeh, M.H.; Khanbabaei, H.; Shamshirian, A.; Amiri Moghadam, S.; ArefNezhad, R.; Sahebkar, A.; Avan, A.; et al. Curcumin inhibits NF-kB and Wnt/beta-catenin pathways in cervical cancer cells. Pathol. Res. Pract. 2019, 215, 152556. [Google Scholar] [CrossRef] [PubMed]
- Kuttikrishnan, S.; Siveen, K.S.; Prabhu, K.S.; Khan, A.Q.; Ahmed, E.I.; Akhtar, S.; Ali, T.A.; Merhi, M.; Dermime, S.; Steinhoff, M.; et al. Curcumin induces apoptotic cell death via inhibition of PI3-kinase/AKT pathway in B-precursor acute lymphoblastic leukemia. Front. Oncol. 2019, 9, 484. [Google Scholar] [CrossRef]
- Mortezaee, K.; Salehi, E.; Mirtavoos-Mahyari, H.; Motevaseli, E.; Najafi, M.; Farhood, B.; Rosengren, R.J.; Sahebkar, A. Mechanisms of apoptosis modulation by curcumin: Implications for cancer therapy. J. Cell. Physiol. 2019, 234, 12537–12550. [Google Scholar] [CrossRef]
- Wahlstrom, B.; Blennow, G. A study on the fate of curcumin in the rat. Acta Pharmacol. Toxicol. 1978, 43, 86–92. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A review of curcumin and its derivatives as anticancer agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; He, Y.; Zhang, R.; Zheng, G.; Zhou, D. The curcumin analog EF24 is a novel senolytic agent. Aging 2019, 11, 771–782. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, W.; Hu, G.; Sun, H.; Kong, Q. Bioactivities of EF24, a novel curcumin analog: A review. Front. Oncol. 2018, 8, 614. [Google Scholar] [CrossRef] [PubMed]
- Jamwal, R. Bioavailable curcumin formulations: A review of pharmacokinetic studies in healthy volunteers. J. Integr. Med. 2018, 16, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Adiwidjaja, J.; McLachlan, A.J.; Boddy, A.V. Curcumin as a clinically-promising anti-cancer agent: Pharmacokinetics and drug interactions. Expert Opin. Drug Metab. Toxicol. 2017, 13, 953–972. [Google Scholar] [CrossRef] [PubMed]
- Olotu, F.; Agoni, C.; Soremekun, O.; Soliman, M.E.S. An update on the pharmacological usage of curcumin: Has it failed in the drug discovery pipeline? Cell Biochem. Biophys. 2020, 78, 267–289. [Google Scholar] [CrossRef]
- Yallapu, M.M.; Nagesh, P.K.; Jaggi, M.; Chauhan, S.C. Therapeutic applications of curcumin nanoformulations. AAPS J. 2015, 17, 1341–1356. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Senthil, N.; Min, T. Nanocurcumin: A promising candidate for therapeutic applications. Front. Pharmacol. 2020, 11, 487. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, N.; He, H.; Tang, X. Pharmaceutical strategies of improving oral systemic bioavailability of curcumin for clinical application. J. Control. Release Off. J. Control. Release Soc. 2019, 316, 359–380. [Google Scholar] [CrossRef]
- Kotecha, R.; Takami, A.; Espinoza, J.L. Dietary phytochemicals and cancer chemoprevention: A review of the clinical evidence. Oncotarget 2016, 7, 52517–52529. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Al Zohairy, M.A.; Aly, S.M.; Khan, M.A. Curcumin: A potential candidate in prevention of cancer via modulation of molecular pathways. BioMed Res. Int. 2014, 2014, 761608. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhang, M.; Dai, E.; Luo, Y. Molecular targets of curcumin in breast cancer (Review). Mol. Med. Rep. 2019, 19, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; Banerjee, S.; Stafford, L.J.; Xia, C.; Liu, M.; Aggarwal, B.B. Curcumin-induced suppression of cell proliferation correlates with down-regulation of cyclin D1 expression and CDK4-mediated retinoblastoma protein phosphorylation. Oncogene 2002, 21, 8852–8861. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.M.; Wang, X.F.; Liu, X.J.; Zhang, H.; Lu, Y.Y.; Su, S.B. Curcumin enhanced antiproliferative effect of mitomycin C in human breast cancer MCF-7 cells in vitro and in vivo. Acta Pharmacol. Sin. 2011, 32, 1402–1410. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Banerjee, S.; Bharadwaj, U.; Sung, B.; Shishodia, S.; Sethi, G. Curcumin induces the degradation of cyclin E expression through ubiquitin-dependent pathway and up-regulates cyclin-dependent kinase inhibitors p21 and p27 in multiple human tumor cell lines. Biochem. Pharmacol. 2007, 73, 1024–1032. [Google Scholar] [CrossRef]
- Prasad, C.P.; Rath, G.; Mathur, S.; Bhatnagar, D.; Ralhan, R. Potent growth suppressive activity of curcumin in human breast cancer cells: Modulation of Wnt/beta-catenin signaling. Chem. Biol. Interact. 2009, 181, 263–271. [Google Scholar] [CrossRef]
- Colomer, C.; Marruecos, L.; Vert, A.; Bigas, A.; Espinosa, L. NF-kappaB members left home: NF-kappaB-independent roles in cancer. Biomedicines 2017, 5, 26. [Google Scholar] [CrossRef]
- Zhou, Q.; Ye, M.; Lu, Y.; Zhang, H.; Chen, Q.; Huang, S.; Su, S. Curcumin improves the tumoricidal effect of mitomycin C by suppressing ABCG2 expression in stem cell-like breast cancer cells. PLoS ONE 2015, 10, e0136694. [Google Scholar] [CrossRef]
- Zhou, X.; Jiao, D.; Dou, M.; Zhang, W.; Lv, L.; Chen, J.; Li, L.; Wang, L.; Han, X. Curcumin inhibits the growth of triple-negative breast cancer cells by silencing EZH2 and restoring DLC1 expression. J. Cell. Mol. Med. 2020, 24, 10648–10662. [Google Scholar] [CrossRef]
- Chase, A.; Cross, N.C. Aberrations of EZH2 in cancer. Clin. Cancer Res. 2011, 17, 2613–2618. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Li, G. Tumor suppressor gene DLC1: Its modifications, interactive molecules, and potential prospects for clinical cancer application. Int. J. Biol. Macromol. 2021, 182, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Pricci, M.; Girardi, B.; Giorgio, F.; Losurdo, G.; Ierardi, E.; Di Leo, A. Curcumin and colorectal cancer: From basic to clinical evidences. Int. J. Mol. Sci. 2020, 21, 2364. [Google Scholar] [CrossRef]
- Calibasi-Kocal, G.; Pakdemirli, A.; Bayrak, S.; Ozupek, N.M.; Sever, T.; Basbinar, Y.; Ellidokuz, H.; Yigitbasi, T. Curcumin effects on cell proliferation, angiogenesis and metastasis in colorectal cancer. J. BUON Off. J. Balk. Union Oncol. 2019, 24, 1482–1487. [Google Scholar]
- Lim, T.G.; Lee, S.Y.; Huang, Z.; Lim, D.Y.; Chen, H.; Jung, S.K.; Bode, A.M.; Lee, K.W.; Dong, Z. Curcumin suppresses proliferation of colon cancer cells by targeting CDK2. Cancer Prev. Res. 2014, 7, 466–474. [Google Scholar] [CrossRef]
- Kim, K.C.; Lee, C. Curcumin induces downregulation of E2F4 expression and apoptotic cell death in HCT116 human colon cancer cells; Involvement of reactive oxygen species. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2010, 14, 391–397. [Google Scholar] [CrossRef]
- Watson, J.L.; Hill, R.; Yaffe, P.B.; Greenshields, A.; Walsh, M.; Lee, P.W.; Giacomantonio, C.A.; Hoskin, D.W. Curcumin causes superoxide anion production and p53-independent apoptosis in human colon cancer cells. Cancer Lett. 2010, 297, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mosieniak, G.; Adamowicz, M.; Alster, O.; Jaskowiak, H.; Szczepankiewicz, A.A.; Wilczynski, G.M.; Ciechomska, I.A.; Sikora, E. Curcumin induces permanent growth arrest of human colon cancer cells: Link between senescence and autophagy. Mech. Ageing Dev. 2012, 133, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, G.; Zhang, R.; Dong, L.; Chen, H.; Bo, J.; Xue, W.; Huang, Y. Curcumin inhibits cell proliferation and motility via suppression of TROP2 in bladder cancer cells. Int. J. Oncol. 2018, 53, 515–526. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, X.; Shi, T.; Li, H. Antitumor effects of curcumin in human bladder cancer in vitro. Oncol. Lett. 2017, 14, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.C.; Mittal, S. Antitumor activity of curcumin in glioblastoma. Int. J. Mol. Sci. 2020, 21, 9435. [Google Scholar] [CrossRef]
- Choi, B.H.; Kim, C.G.; Bae, Y.S.; Lim, Y.; Lee, Y.H.; Shin, S.Y. P21(Waf1/Cip1) expression by curcumin in U-87MG human glioma cells: Role of early growth response-1 expression. Cancer Res. 2008, 68, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Jiao, J.T.; Qian, Y.; Guo, X.Y.; Huang, J.; Dai, M.C.; Zhang, L.; Ding, X.P.; Zong, D.; Shao, J.F. Curcumin induces G2/M arrest and triggers apoptosis via FoxO1 signaling in U87 human glioma cells. Mol. Med. Rep. 2016, 13, 3763–3770. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ye, X.; Cai, X.; Su, J.; Ma, R.; Yin, X.; Zhou, X.; Li, H.; Wang, Z. Curcumin suppresses cell growth and invasion and induces apoptosis by down-regulation of Skp2 pathway in glioma cells. Oncotarget 2015, 6, 18027–18037. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Su, H.K.; Yu, Z.H.; Xi, S.Y.; Guo, C.C.; Hu, Z.Y.; Qu, Y.; Cai, H.P.; Zhao, Y.Y.; Zhao, H.F.; et al. Skp2 modulates proliferation, senescence and tumorigenesis of glioma. Cancer Cell Int. 2020, 20, 71. [Google Scholar] [CrossRef]
- Luo, Q.; Luo, H.; Fu, H.; Huang, H.; Huang, H.; Luo, K.; Li, C.; Hu, R.; Zheng, C.; Lan, C.; et al. Curcumin suppresses invasiveness and migration of human glioma cells in vitro by inhibiting HDGF/beta-catenin complex. Nan Fang Yi Ke Da Xue Xue Bao J. South. Med. Univ. 2019, 39, 911–916. [Google Scholar] [CrossRef]
- Fratantonio, D.; Molonia, M.S.; Bashllari, R.; Muscara, C.; Ferlazzo, G.; Costa, G.; Saija, A.; Cimino, F.; Speciale, A. Curcumin potentiates the antitumor activity of Paclitaxel in rat glioma C6 cells. Phytomed. Int. J. Phytother. Phytopharm. 2019, 55, 23–30. [Google Scholar] [CrossRef]
- Seyithanoglu, M.H.; Abdallah, A.; Kitis, S.; Guler, E.M.; Kocyigit, A.; Dundar, T.T.; Gundag Papaker, M. Investigation of cytotoxic, genotoxic, and apoptotic effects of curcumin on glioma cells. Cell. Mol. Biol. 2019, 65, 101–108. [Google Scholar] [CrossRef]
- Murray-Stewart, T.; Dunworth, M.; Lui, Y.; Giardiello, F.M.; Woster, P.M.; Casero, R.A., Jr. Curcumin mediates polyamine metabolism and sensitizes gastrointestinal cancer cells to antitumor polyamine-targeted therapies. PLoS ONE 2018, 13, e0202677. [Google Scholar] [CrossRef]
- Cai, X.Z.; Wang, J.; Li, X.D.; Wang, G.L.; Liu, F.N.; Cheng, M.S.; Li, F. Curcumin suppresses proliferation and invasion in human gastric cancer cells by downregulation of PAK1 activity and cyclin D1 expression. Cancer Biol. Ther. 2009, 8, 1360–1368. [Google Scholar] [CrossRef]
- Zheng, R.; Deng, Q.; Liu, Y.; Zhao, P. Curcumin inhibits gastric carcinoma cell growth and induces apoptosis by suppressing the Wnt/beta-catenin signaling pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2017, 23, 163–171. [Google Scholar] [CrossRef]
- Fu, H.; Wang, C.; Yang, D.; Wei, Z.; Xu, J.; Hu, Z.; Zhang, Y.; Wang, W.; Yan, R.; Cai, Q. Curcumin regulates proliferation, autophagy, and apoptosis in gastric cancer cells by affecting PI3K and P53 signaling. J. Cell. Physiol. 2018, 233, 4634–4642. [Google Scholar] [CrossRef]
- Liu, G.; Xiang, T.; Wu, Q.F.; Wang, W.X. Curcumin suppresses the proliferation of gastric cancer cells by downregulating H19. Oncol. Lett. 2016, 12, 5156–5162. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Esmaeili, M.; Taheri, M. H19 lncRNA: Roles in tumorigenesis. Biomed. Pharmacother. 2020, 123, 109774. [Google Scholar] [CrossRef]
- Yang, F.; Bi, J.; Xue, X.; Zheng, L.; Zhi, K.; Hua, J.; Fang, G. Up-regulated long non-coding RNA H19 contributes to proliferation of gastric cancer cells. FEBS J. 2012, 279, 3159–3165. [Google Scholar] [CrossRef]
- Yoshimura, H.; Matsuda, Y.; Yamamoto, M.; Kamiya, S.; Ishiwata, T. Expression and role of long non-coding RNA H19 in carcinogenesis. Front. Biosci. 2018, 23, 614–625. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, S.; Liu, C.; Liu, X. Curcumin promoted miR-34a expression and suppressed proliferation of gastric cancer cells. Cancer Biother. Radiopharm. 2019, 34, 634–641. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, W.; Guo, Y.; Li, Z.; Chen, X.; Wang, Y.; Du, Y.; Zang, W.; Zhao, G. Curcumin inhibits cell growth and induces cell apoptosis through upregulation of miR-33b in gastric cancer. Tumor Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 13177–13184. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.M.; Russo, G.L.; Tedesco, I.; Daglia, M.; Orhan, I.E.; Nabavi, S.F.; Bishayee, A.; Nagulapalli Venkata, K.C.; Abdollahi, M.; Hajheydari, Z. Curcumin and melanoma: From chemistry to medicine. Nutr. Cancer 2018, 70, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Ekmekcioglu, S.; Walch, E.T.; Tang, C.H.; Grimm, E.A. Inhibition of nuclear factor-kappaB and nitric oxide by curcumin induces G2/M cell cycle arrest and apoptosis in human melanoma cells. Melanoma Res. 2004, 14, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Abusnina, A.; Keravis, T.; Yougbare, I.; Bronner, C.; Lugnier, C. Anti-proliferative effect of curcumin on melanoma cells is mediated by PDE1A inhibition that regulates the epigenetic integrator UHRF1. Mol. Nutr. Food Res. 2011, 55, 1677–1689. [Google Scholar] [CrossRef]
- Zhao, G.; Han, X.; Zheng, S.; Li, Z.; Sha, Y.; Ni, J.; Sun, Z.; Qiao, S.; Song, Z. Curcumin induces autophagy, inhibits proliferation and invasion by downregulating AKT/mTOR signaling pathway in human melanoma cells. Oncol. Rep. 2016, 35, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Siwak, D.R.; Shishodia, S.; Aggarwal, B.B.; Kurzrock, R. Curcumin-induced antiproliferative and proapoptotic effects in melanoma cells are associated with suppression of I kappa B kinase and nuclear factor KB activity and are independent of the B-Raf/mitogen-activated/extracellular signal-regulated protein kinase pathway and the Akt pathway. Cancer 2005, 104, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Cianfruglia, L.; Minnelli, C.; Laudadio, E.; Scire, A.; Armeni, T. Side effects of curcumin: Epigenetic and antiproliferative implications for normal dermal fibroblast and breast cancer cells. Antioxidants 2019, 8, 382. [Google Scholar] [CrossRef] [PubMed]
- Talib, W.H.; Al-Hadid, S.A.; Ali, M.B.W.; Al-Yasari, I.H.; Ali, M.R.A. Role of curcumin in regulating p53 in breast cancer: An overview of the mechanism of action. Breast Cancer 2018, 10, 207–217. [Google Scholar] [CrossRef]
- Bae, Y.H.; Ryu, J.H.; Park, H.J.; Kim, K.R.; Wee, H.J.; Lee, O.H.; Jang, H.O.; Bae, M.K.; Kim, K.W.; Bae, S.K. Mutant p53-Notch1 signaling axis is involved in curcumin-induced apoptosis of breast cancer cells. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2013, 17, 291–297. [Google Scholar] [CrossRef]
- Patel, P.B.; Thakkar, V.R.; Patel, J.S. Cellular effect of curcumin and citral combination on breast cancer cells: Induction of apoptosis and cell cycle arrest. J. Breast Cancer 2015, 18, 225–234. [Google Scholar] [CrossRef]
- Moghtaderi, H.; Sepehri, H.; Attari, F. Combination of arabinogalactan and curcumin induces apoptosis in breast cancer cells in vitro and inhibits tumor growth via overexpression of p53 level in vivo. Biomed. Pharmacother. 2017, 88, 582–594. [Google Scholar] [CrossRef]
- Chiu, T.L.; Su, C.C. Curcumin inhibits proliferation and migration by increasing the Bax to Bcl-2 ratio and decreasing NF-kappaBp65 expression in breast cancer MDA-MB-231 cells. Int. J. Mol. Med. 2009, 23, 469–475. [Google Scholar] [CrossRef][Green Version]
- Fan, H.; Liang, Y.; Jiang, B.; Li, X.; Xun, H.; Sun, J.; He, W.; Lau, H.T.; Ma, X. Curcumin inhibits intracellular fatty acid synthase and induces apoptosis in human breast cancer MDA-MB-231 cells. Oncol. Rep. 2016, 35, 2651–2656. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Li, Y.Q.; Lv, Y.T.; Wang, J.M. Effect of curcumin on the proliferation, apoptosis, migration, and invasion of human melanoma A375 cells. Genet. Mol. Res. 2015, 14, 1056–1067. [Google Scholar] [CrossRef]
- Bush, J.A.; Cheung, K.J., Jr.; Li, G. Curcumin induces apoptosis in human melanoma cells through a Fas receptor/caspase-8 pathway independent of p53. Exp. Cell Res. 2001, 271, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.L.; Greenshields, A.; Hill, R.; Hilchie, A.; Lee, P.W.; Giacomantonio, C.A.; Hoskin, D.W. Curcumin-induced apoptosis in ovarian carcinoma cells is p53-independent and involves p38 mitogen-activated protein kinase activation and downregulation of Bcl-2 and survivin expression and Akt signaling. Mol. Carcinog. 2010, 49, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, Y.; Song, Y.; Xu, L.; Zhao, J.; Fang, B. A preliminary study of the effect of curcumin on the expression of p53 protein in a human multiple myeloma cell line. Oncol. Lett. 2015, 9, 1719–1724. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Park, S.Y.; Kim, Y.M.; Park, O.J. Regulatory effect of the AMPK-COX-2 signaling pathway in curcumin-induced apoptosis in HT-29 colon cancer cells. Ann. N. Y. Acad. Sci. 2009, 1171, 489–494. [Google Scholar] [CrossRef]
- Sandur, S.K.; Deorukhkar, A.; Pandey, M.K.; Pabon, A.M.; Shentu, S.; Guha, S.; Aggarwal, B.B.; Krishnan, S. Curcumin modulates the radiosensitivity of colorectal cancer cells by suppressing constitutive and inducible NF-kappaB activity. Int. J. Radiat. Oncol. Biol. Phys. 2009, 75, 534–542. [Google Scholar] [CrossRef]
- Narayan, S. Curcumin, a multi-functional chemopreventive agent, blocks growth of colon cancer cells by targeting beta-catenin-mediated transactivation and cell-cell adhesion pathways. J. Mol. Histol. 2004, 35, 301–307. [Google Scholar] [CrossRef]
- Jung, E.M.; Lim, J.H.; Lee, T.J.; Park, J.W.; Choi, K.S.; Kwon, T.K. Curcumin sensitizes tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis through reactive oxygen species-mediated upregulation of death receptor 5 (DR5). Carcinogenesis 2005, 26, 1905–1913. [Google Scholar] [CrossRef]
- Cao, A.; Li, Q.; Yin, P.; Dong, Y.; Shi, H.; Wang, L.; Ji, G.; Xie, J.; Wu, D. Curcumin induces apoptosis in human gastric carcinoma AGS cells and colon carcinoma HT-29 cells through mitochondrial dysfunction and endoplasmic reticulum stress. Apoptosis 2013, 18, 1391–1402. [Google Scholar] [CrossRef]
- Moragoda, L.; Jaszewski, R.; Majumdar, A.P. Curcumin induced modulation of cell cycle and apoptosis in gastric and colon cancer cells. Anticancer Res. 2001, 21, 873–878. [Google Scholar]
- Su, C.C.; Lin, J.G.; Li, T.M.; Chung, J.G.; Yang, J.S.; Ip, S.W.; Lin, W.C.; Chen, G.W. Curcumin-induced apoptosis of human colon cancer colo 205 cells through the production of ROS, Ca2+ and the activation of caspase-3. Anticancer Res. 2006, 26, 4379–4389. [Google Scholar]
- Klinger, N.V.; Mittal, S. Therapeutic potential of curcumin for the treatment of brain tumors. Oxidative Med. Cell. Longev. 2016, 2016, 9324085. [Google Scholar] [CrossRef]
- Lee, D.S.; Lee, M.K.; Kim, J.H. Curcumin induces cell cycle arrest and apoptosis in human osteosarcoma (HOS) cells. Anticancer Res. 2009, 29, 5039–5044. [Google Scholar] [PubMed]
- Yang, J.; Cao, Y.; Sun, J.; Zhang, Y. Curcumin reduces the expression of Bcl-2 by upregulating miR-15a and miR-16 in MCF-7 cells. Med. Oncol. 2010, 27, 1114–1118. [Google Scholar] [CrossRef]
- Zhang, J.; Du, Y.; Wu, C.; Ren, X.; Ti, X.; Shi, J.; Zhao, F.; Yin, H. Curcumin promotes apoptosis in human lung adenocarcinoma cells through miR-186* signaling pathway. Oncol. Rep. 2010, 24, 1217–1223. [Google Scholar] [CrossRef]
- Sohn, E.J.; Bak, K.M.; Nam, Y.K.; Park, H.T. Upregulation of microRNA 344a-3p is involved in curcumin induced apoptosis in RT4 schwannoma cells. Cancer Cell Int. 2018, 18, 199. [Google Scholar] [CrossRef]
- Gao, D.; Mittal, V.; Ban, Y.; Lourenco, A.R.; Yomtoubian, S.; Lee, S. Metastatic tumor cells—Genotypes and phenotypes. Front. Biol. 2018, 13, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, M.; Kemmerling, U.; Aguayo, F.; Bleak, T.C.; Munoz, J.P.; Calaf, G.M. Curcumin rescues breast cells from epithelialmesenchymal transition and invasion induced by antimiR34a. Int. J. Oncol. 2020, 56, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Pires, B.R.; Mencalha, A.L.; Ferreira, G.M.; de Souza, W.F.; Morgado-Diaz, J.A.; Maia, A.M.; Correa, S.; Abdelhay, E.S. NF-kappaB is involved in the regulation of EMT genes in breast cancer cells. PLoS ONE 2017, 12, e0169622. [Google Scholar] [CrossRef] [PubMed]
- Zong, H.; Wang, F.; Fan, Q.X.; Wang, L.X. Curcumin inhibits metastatic progression of breast cancer cell through suppression of urokinase-type plasminogen activator by NF-kappa B signaling pathways. Mol. Biol. Rep. 2012, 39, 4803–4808. [Google Scholar] [CrossRef]
- Bachmeier, B.E.; Mohrenz, I.V.; Mirisola, V.; Schleicher, E.; Romeo, F.; Hohneke, C.; Jochum, M.; Nerlich, A.G.; Pfeffer, U. Curcumin downregulates the inflammatory cytokines CXCL1 and -2 in breast cancer cells via NFkappaB. Carcinogenesis 2008, 29, 779–789. [Google Scholar] [CrossRef]
- Irani, S. Emerging insights into the biology of metastasis: A review article. Iran. J. Basic Med. Sci. 2019, 22, 833–847. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Duan, X.; Cai, H.; Liu, X.; Yang, Y.; Li, M.; Zhang, X.; Wang, J. Curcumin inhibits LPA-induced invasion by attenuating RhoA/ROCK/MMPs pathway in MCF7 breast cancer cells. Clin. Exp. Med. 2016, 16, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, W.; Han, N.; Zou, Y.; Yin, D. Curcumin inhibits proliferation, migration, invasion and promotes apoptosis of retinoblastoma cell lines through modulation of miR-99a and JAK/STAT pathway. BMC Cancer 2018, 18, 1230. [Google Scholar] [CrossRef]
- Zaman, S.; Jadid, H.; Denson, A.C.; Gray, J.E. Targeting Trop-2 in solid tumors: Future prospects. Onco Targets Ther. 2019, 12, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.S.; Chen, W.C.; Lin, Y.Y.; Tsai, C.H.; Liao, C.I.; Shyu, H.Y.; Ko, C.J.; Tzeng, S.F.; Huang, C.Y.; Yang, P.C.; et al. Curcumin-targeting pericellular serine protease matriptase role in suppression of prostate cancer cell invasion, tumor growth, and metastasis. Cancer Prev. Res. 2013, 6, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; He, B.; Liu, Q.; Hu, D.; Liao, W.; Li, R.; Peng, X.; Wang, Q.; Zhao, G. Antitumor effects of curcumin on the proliferation, migration and apoptosis of human colorectal carcinoma HCT116 cells. Oncol. Rep. 2020, 44, 1997–2008. [Google Scholar] [CrossRef]
- Philip, S.; Kundu, G.C. Osteopontin induces nuclear factor kappa B-mediated promatrix metalloproteinase-2 activation through I kappa B alpha /IKK signaling pathways, and curcumin (diferulolylmethane) down-regulates these pathways. J. Biol. Chem. 2003, 278, 14487–14497. [Google Scholar] [CrossRef]
- Jin, W. Role of JAK/STAT3 signaling in the regulation of metastasis, the transition of cancer stem cells, and chemoresistance of cancer by epithelial-mesenchymal transition. Cells 2020, 9, 217. [Google Scholar] [CrossRef]
- Yang, C.L.; Liu, Y.Y.; Ma, Y.G.; Xue, Y.X.; Liu, D.G.; Ren, Y.; Liu, X.B.; Li, Y.; Li, Z. Curcumin blocks small cell lung cancer cells migration, invasion, angiogenesis, cell cycle and neoplasia through Janus kinase-STAT3 signalling pathway. PLoS ONE 2012, 7, e37960. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, J.; Zhang, K.; Zhao, Y. Curcumin inhibits cell viability, migration, and invasion of thymic carcinoma cells via downregulation of microRNA-27a. Phytother. Res. PTR 2020, 34, 1629–1637. [Google Scholar] [CrossRef]
- Wang, N.; Feng, T.; Liu, X.; Liu, Q. Curcumin inhibits migration and invasion of non-small cell lung cancer cells through up-regulation of miR-206 and suppression of PI3K/AKT/mTOR signaling pathway. Acta Pharm. 2020, 70, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Deng, J.; Yuan, J.; Tang, X.; Wang, Y.; Chen, H.; Liu, Y.; Zhou, L. Curcumin exerts its tumor suppressive function via inhibition of NEDD4 oncoprotein in glioma cancer cells. Int. J. Oncol. 2017, 51, 467–477. [Google Scholar] [CrossRef]
- Zhang, H.; Nie, W.; Zhang, X.; Zhang, G.; Li, Z.; Wu, H.; Shi, Q.; Chen, Y.; Ding, Z.; Zhou, X.; et al. NEDD4-1 regulates migration and invasion of glioma cells through CNrasGEF ubiquitination in vitro. PLoS ONE 2013, 8, e82789. [Google Scholar] [CrossRef]
- Ravindranath, V.; Chandrasekhara, N. Absorption and tissue distribution of curcumin in rats. Toxicology 1980, 16, 259–265. [Google Scholar] [CrossRef]
- Pan, M.H.; Huang, T.M.; Lin, J.K. Biotransformation of curcumin through reduction and glucuronidation in mice. Drug Metab. Dispos. 1999, 27, 486–494. [Google Scholar]
- Basile, V.; Ferrari, E.; Lazzari, S.; Belluti, S.; Pignedoli, F.; Imbriano, C. Curcumin derivatives: Molecular basis of their anti-cancer activity. Biochem. Pharmacol. 2009, 78, 1305–1315. [Google Scholar] [CrossRef]
- Sribalan, R.; Kirubavathi, M.; Banuppriya, G.; Padmini, V. Synthesis and biological evaluation of new symmetric curcumin derivatives. Bioorg. Med. Chem. Lett. 2015, 25, 4282–4286. [Google Scholar] [CrossRef]
- Bayomi, S.M.; El-Kashef, H.A.; El-Ashmawy, M.B.; Nasr, M.N.A.; El-Sherbeny, M.A.; Badria, F.A.; Abou-Zeid, L.A.; Ghaly, M.A.; Abdel-Aziz, N.I. Synthesis and biological evaluation of new curcumin derivatives as antioxidant and antitumor agents. Med. Chem. Res. 2013, 22, 1147–1162. [Google Scholar] [CrossRef]
- Xu, G.; Chu, Y.; Jiang, N.; Yang, J.; Li, F. The three dimensional quantitative structure activity relationships (3D-QSAR) and docking studies of curcumin derivatives as androgen receptor antagonists. Int. J. Mol. Sci. 2012, 13, 6138–6155. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, Z.; Liang, G. Promising curcumin-based drug design: Mono-carbonyl analogues of curcumin (MACs). Curr. Pharm. Des. 2013, 19, 2114–2135. [Google Scholar] [PubMed]
- Somparn, P.; Phisalaphong, C.; Nakornchai, S.; Unchern, S.; Morales, N.P. Comparative antioxidant activities of curcumin and its demethoxy and hydrogenated derivatives. Biol. Pharm. Bull. 2007, 30, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Thomas, S.G.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Sung, B.; Tharakan, S.T.; Misra, K.; Priyadarsini, I.K.; Rajasekharan, K.N.; et al. Biological activities of curcumin and its analogues (Congeners) made by man and Mother Nature. Biochem. Pharmacol. 2008, 76, 1590–1611. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.R.; Dinesh, P.; Prabhakar, A.S.; Umasankar, K.; Shireesha, B.; Raju, M.B. A comprehensive review on SAR of curcumin. Mini Rev. Med. Chem. 2013, 13, 1769–1777. [Google Scholar] [CrossRef] [PubMed]
- Sherin, D.R.; Rajasekharan, K.N. Mechanochemical synthesis and antioxidant activity of curcumin-templated azoles. Arch. Pharm. 2015, 348, 908–914. [Google Scholar] [CrossRef]
- Wanninger, S.; Lorenz, V.; Subhan, A.; Edelmann, F.T. Metal complexes of curcumin--synthetic strategies, structures and medicinal applications. Chem. Soc. Rev. 2015, 44, 4986–5002. [Google Scholar] [CrossRef]
- Paulraj, F.; Abas, F.; Lajis, N.H.; Othman, I.; Naidu, R. Molecular pathways modulated by curcumin analogue, diarylpentanoids in cancer. Biomolecules 2019, 9, 270. [Google Scholar] [CrossRef]
- Yerdelen, K.O.; Gul, H.I.; Sakagami, H.; Umemura, N.; Sukuroglu, M. Synthesis and cytotoxic activities of a curcumin analogue and its bis-mannich derivatives. Lett. Drug Des. Discov. 2015, 12, 643–649. [Google Scholar] [CrossRef]
- Reid, J.M.; Buhrow, S.A.; Gilbert, J.A.; Jia, L.; Shoji, M.; Snyder, J.P.; Ames, M.M. Mouse pharmacokinetics and metabolism of the curcumin analog, 4-piperidinone,3,5-bis[(2-fluorophenyl)methylene]-acetate(3E,5E) (EF-24; NSC 716993). Cancer Chemother. Pharmacol. 2014, 73, 1137–1146. [Google Scholar] [CrossRef]
- Selvendiran, K.; Tong, L.; Vishwanath, S.; Bratasz, A.; Trigg, N.J.; Kutala, V.K.; Hideg, K.; Kuppusamy, P. EF24 induces G2/M arrest and apoptosis in cisplatin-resistant human ovarian cancer cells by increasing PTEN expression. J. Biol. Chem. 2007, 282, 28609–28618. [Google Scholar] [CrossRef]
- He, G.; Feng, C.; Vinothkumar, R.; Chen, W.; Dai, X.; Chen, X.; Ye, Q.; Qiu, C.; Zhou, H.; Wang, Y.; et al. Curcumin analog EF24 induces apoptosis via ROS-dependent mitochondrial dysfunction in human colorectal cancer cells. Cancer Chemother. Pharmacol. 2016, 78, 1151–1161. [Google Scholar] [CrossRef]
- Kasinski, A.L.; Du, Y.; Thomas, S.L.; Zhao, J.; Sun, S.Y.; Khuri, F.R.; Wang, C.Y.; Shoji, M.; Sun, A.; Snyder, J.P.; et al. Inhibition of IkappaB kinase-nuclear factor-kappaB signaling pathway by 3,5-bis(2-flurobenzylidene)piperidin-4-one (EF24), a novel monoketone analog of curcumin. Mol. Pharmacol. 2008, 74, 654–661. [Google Scholar] [CrossRef]
- Thomas, S.L.; Zhong, D.; Zhou, W.; Malik, S.; Liotta, D.; Snyder, J.P.; Hamel, E.; Giannakakou, P. EF24, a novel curcumin analog, disrupts the microtubule cytoskeleton and inhibits HIF-1. Cell Cycle 2008, 7, 2409–2417. [Google Scholar] [CrossRef]
- Tan, X.; Sidell, N.; Mancini, A.; Huang, R.P.; Shenming, W.; Horowitz, I.R.; Liotta, D.C.; Taylor, R.N.; Wieser, F. Multiple anticancer activities of EF24, a novel curcumin analog, on human ovarian carcinoma cells. Reprod. Sci. 2010, 17, 931–940. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-kappaB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.L.; Liang, Y.J.; Zheng, T.S.; Song, R.P.; Wang, J.B.; Sun, B.S.; Pan, S.H.; Qu, L.D.; Liu, J.R.; Jiang, H.C.; et al. EF24 inhibits tumor growth and metastasis via suppressing NF-kappaB dependent pathways in human cholangiocarcinoma. Sci. Rep. 2016, 6, 32167. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, S.; Natarajan, M.; Herman, T.S.; Awasthi, V.; Aravindan, N. Molecular basis of ‘hypoxic’ breast cancer cell radio-sensitization: Phytochemicals converge on radiation induced Rel signaling. Radiat. Oncol 2013, 8, 46. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, S.; Natarajan, M.; Awasthi, V.; Herman, T.S.; Aravindan, N. Novel synthetic monoketone transmute radiation-triggered NFkappaB-dependent TNFalpha cross-signaling feedback maintained NFkappaB and favors neuroblastoma regression. PLoS ONE 2013, 8, e72464. [Google Scholar] [CrossRef][Green Version]
- Liang, Y.; Zheng, T.; Song, R.; Wang, J.; Yin, D.; Wang, L.; Liu, H.; Tian, L.; Fang, X.; Meng, X.; et al. Hypoxia-mediated sorafenib resistance can be overcome by EF24 through Von Hippel-Lindau tumor suppressor-dependent HIF-1alpha inhibition in hepatocellular carcinoma. Hepatology 2013, 57, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.K.; Cai, J.; Armstrong, J.; Herold, M.; Lu, Y.J.; Sun, A.; Snyder, J.P.; Liotta, D.C.; Jones, D.P.; Shoji, M. EF24, a novel synthetic curcumin analog, induces apoptosis in cancer cells via a redox-dependent mechanism. Anti-Cancer Drugs 2005, 16, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Xia, Y.; Chen, W.; Chen, X.; Ying, S.; Feng, Z.; Chen, T.; Ye, Q.; Wang, Z.; Qiu, C.; et al. EF24 induces ROS-mediated apoptosis via targeting thioredoxin reductase 1 in gastric cancer cells. Oncotarget 2016, 7, 18050–18064. [Google Scholar] [CrossRef] [PubMed]
- Selvendiran, K.; Ahmed, S.; Dayton, A.; Kuppusamy, M.L.; Rivera, B.K.; Kalai, T.; Hideg, K.; Kuppusamy, P. HO-3867, a curcumin analog, sensitizes cisplatin-resistant ovarian carcinoma, leading to therapeutic synergy through STAT3 inhibition. Cancer Biol. Ther. 2011, 12, 837–845. [Google Scholar] [CrossRef]
- Ismail, N.I.; Othman, I.; Abas, F.; Lajis, N.H.; Naidu, R. The curcumin analogue, MS13 (1,5-Bis(4-hydroxy-3- methoxyphenyl)-1,4-pentadiene-3-one), inhibits cell proliferation and induces apoptosis in primary and metastatic human colon cancer cells. Molecules 2020, 25, 3798. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Shao, L.; Wang, Y.; Zhao, C.; Chu, Y.; Xiao, J.; Zhao, Y.; Li, X.; Yang, S. Exploration and synthesis of curcumin analogues with improved structural stability both in vitro and in vivo as cytotoxic agents. Bioorg. Med. Chem. 2009, 17, 2623–2631. [Google Scholar] [CrossRef]
- Lin, L.; Shi, Q.; Nyarko, A.K.; Bastow, K.F.; Wu, C.C.; Su, C.Y.; Shih, C.C.; Lee, K.H. Antitumor agents. 250. Design and synthesis of new curcumin analogues as potential anti-prostate cancer agents. J. Med. Chem. 2006, 49, 3963–3972. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Shen, J.; Pan, H.; Xu, L.; Sheng, H.; Liu, B.; Yao, M. Curcumin analog B14 has high bioavailability and enhances the effect of anti-breast cancer cells in vitro and in vivo. Cancer Sci. 2021, 112, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; McClements, D.J. Formulation of more efficacious curcumin delivery systems using colloid science: Enhanced solubility, stability, and bioavailability. Molecules 2020, 25, 2791. [Google Scholar] [CrossRef]
- Bansal, S.S.; Goel, M.; Aqil, F.; Vadhanam, M.V.; Gupta, R.C. Advanced drug delivery systems of curcumin for cancer chemoprevention. Cancer Prev. Res. 2011, 4, 1158–1171. [Google Scholar] [CrossRef] [PubMed]
- Munjal, B.; Pawar, Y.B.; Patel, S.B.; Bansal, A.K. Comparative oral bioavailability advantage from curcumin formulations. Drug Deliv. Transl. Res. 2011, 1, 322–331. [Google Scholar] [CrossRef]
- Sun, M.; Su, X.; Ding, B.; He, X.; Liu, X.; Yu, A.; Lou, H.; Zhai, G. Advances in nanotechnology-based delivery systems for curcumin. Nanomedicine 2012, 7, 1085–1100. [Google Scholar] [CrossRef]
- Praditya, D.; Kirchhoff, L.; Bruning, J.; Rachmawati, H.; Steinmann, J.; Steinmann, E. Anti-infective Properties of the Golden Spice Curcumin. Front. Microbiol. 2019, 10, 912. [Google Scholar] [CrossRef]
- Yang, S.; Liu, L.; Han, J.; Tang, Y. Encapsulating plant ingredients for dermocosmetic application: An updated review of delivery systems and characterization techniques. Int. J. Cosmet. Sci. 2020, 42, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Lim, S.J.; Choi, H.G.; Lee, M.K. Solid lipid nanoparticles as drug delivery system for water-insoluble drugs. J. Pharm. Investig. 2010, 40, 63–73. [Google Scholar] [CrossRef][Green Version]
- Yallapu, M.M.; Jaggi, M.; Chauhan, S.C. beta-Cyclodextrin-curcumin self-assembly enhances curcumin delivery in prostate cancer cells. Colloids Surf. B Biointerfaces 2010, 79, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A review of its effects on human health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Umerska, A.; Gaucher, C.; Oyarzun-Ampuero, F.; Fries-Raeth, I.; Colin, F.; Villamizar-Sarmiento, M.G.; Maincent, P.; Sapin-Minet, A. Polymeric nanoparticles for increasing oral bioavailability of curcumin. Antioxidants 2018, 7, 46. [Google Scholar] [CrossRef]
- Ferrari, R.; Sponchioni, M.; Morbidelli, M.; Moscatelli, D. Polymer nanoparticles for the intravenous delivery of anticancer drugs: The checkpoints on the road from the synthesis to clinical translation. Nanoscale 2018, 10, 22701–22719. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Shi, M.; Li, N.; Xu, R. Application of functional biocompatible nanomaterials to improve curcumin bioavailability. Front. Chem. 2020, 8, 589957. [Google Scholar] [CrossRef]
- Zaman, M.S.; Chauhan, N.; Yallapu, M.M.; Gara, R.K.; Maher, D.M.; Kumari, S.; Sikander, M.; Khan, S.; Zafar, N.; Jaggi, M.; et al. Curcumin nanoformulation for cervical cancer treatment. Sci. Rep. 2016, 6, 20051. [Google Scholar] [CrossRef]
- Yallapu, M.M.; Gupta, B.K.; Jaggi, M.; Chauhan, S.C. Fabrication of curcumin encapsulated PLGA nanoparticles for improved therapeutic effects in metastatic cancer cells. J. Colloid Interface Sci. 2010, 351, 19–29. [Google Scholar] [CrossRef]
- Mukerjee, A.; Vishwanatha, J.K. Formulation, characterization and evaluation of curcumin-loaded PLGA nanospheres for cancer therapy. Anticancer Res. 2009, 29, 3867–3875. [Google Scholar]
- Bisht, S.; Feldmann, G.; Soni, S.; Ravi, R.; Karikar, C.; Maitra, A.; Maitra, A. Polymeric nanoparticle-encapsulated curcumin (“nanocurcumin”): A novel strategy for human cancer therapy. J. Nanobiotechnol. 2007, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.T.; Zhang, D.G.; Wu, X.L.; Huang, X.E.; Chen, G. In vivo evaluation of curcumin-loaded nanoparticles in a A549 xenograft mice model. Asian Pac. J. Cancer Prev. APJCP 2013, 14, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Fan, D.; Li, Y.; He, X.; Chen, X.; Chen, Y.; Zhu, J.; Xu, G.; Wu, X.; Lan, P. Supercritical carbon dioxide-developed silk fibroin nanoplatform for smart colon cancer therapy. Int. J. Nanomed. 2017, 12, 7751–7761. [Google Scholar] [CrossRef] [PubMed]
- Baspinar, Y.; Ustundas, M.; Bayraktar, O.; Sezgin, C. Curcumin and piperine loaded zein-chitosan nanoparticles: Development and in-vitro characterisation. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2018, 26, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Bandyopadhyay, A.; Chakraborty, A.; Sarkar, K. Enhancement of anticancer activity and drug delivery of chitosan-curcumin nanoparticle via molecular docking and simulation analysis. Carbohydr. Polym. 2018, 182, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Manjunath, K.; Reddy, J.S.; Venkateswarlu, V. Solid lipid nanoparticles as drug delivery systems. Methods Find. Exp. Clin. Pharmacol. 2005, 27, 127–144. [Google Scholar] [CrossRef]
- Mukherjee, S.; Ray, S.; Thakur, R.S. Solid lipid nanoparticles: A modern formulation approach in drug delivery system. Indian J. Pharm. Sci. 2009, 71, 349–358. [Google Scholar] [CrossRef]
- Jenning, V.; Lippacher, A.; Gohla, S.H. Medium scale production of solid lipid nanoparticles (SLN) by high pressure homogenization. J. Microencapsul. 2002, 19, 1–10. [Google Scholar] [CrossRef]
- Baek, J.S.; Cho, C.W. Surface modification of solid lipid nanoparticles for oral delivery of curcumin: Improvement of bioavailability through enhanced cellular uptake, and lymphatic uptake. Eur. J. Pharm. Biopharm. 2017, 117, 132–140. [Google Scholar] [CrossRef]
- Guorgui, J.; Wang, R.; Mattheolabakis, G.; Mackenzie, G.G. Curcumin formulated in solid lipid nanoparticles has enhanced efficacy in Hodgkin’s lymphoma in mice. Arch. Biochem. Biophys. 2018, 648, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Gota, V.S.; Maru, G.B.; Soni, T.G.; Gandhi, T.R.; Kochar, N.; Agarwal, M.G. Safety and pharmacokinetics of a solid lipid curcumin particle formulation in osteosarcoma patients and healthy volunteers. J. Agric. Food Chem. 2010, 58, 2095–2099. [Google Scholar] [CrossRef]
- Ban, C.; Jo, M.; Park, Y.H.; Kim, J.H.; Han, J.Y.; Lee, K.W.; Kweon, D.H.; Choi, Y.J. Enhancing the oral bioavailability of curcumin using solid lipid nanoparticles. Food Chem. 2020, 302, 125328. [Google Scholar] [CrossRef]
- Huang, S.; He, J.; Cao, L.; Lin, H.; Zhang, W.; Zhong, Q. Improved physicochemical properties of curcumin-loaded solid lipid nanoparticles stabilized by sodium caseinate-lactose Maillard conjugate. J. Agric. Food Chem. 2020, 68, 7072–7081. [Google Scholar] [CrossRef]
- Bhatt, H.; Rompicharla, S.V.K.; Komanduri, N.; Aashma, S.; Paradkar, S.; Ghosh, B.; Biswas, S. Development of curcumin-loaded solid lipid nanoparticles utilizing glyceryl monostearate as single lipid using QbD approach: Characterization and evaluation of anticancer activity against human breast cancer cell line. Curr. Drug Deliv. 2018, 15, 1271–1283. [Google Scholar] [CrossRef]
- Wang, F.; Li, C.; Cheng, J.; Yuan, Z. Recent advances on inorganic nanoparticle-based cancer therapeutic agents. Int. J. Environ. Res. Public Health 2016, 13, 1182. [Google Scholar] [CrossRef]
- Huynh, K.H.; Pham, X.H.; Kim, J.; Lee, S.H.; Chang, H.; Rho, W.Y.; Jun, B.H. Synthesis, properties, and biological applications of metallic alloy nanoparticles. Int. J. Mol. Sci. 2020, 21, 5174. [Google Scholar] [CrossRef]
- Kouhpanji, M.R.Z.; Stadler, B.J.H. A guideline for effectively synthesizing and characterizing magnetic nanoparticles for advancing nanobiotechnology: A review. Sensors 2020, 20, 2554. [Google Scholar] [CrossRef] [PubMed]
- Yallapu, M.M.; Ebeling, M.C.; Khan, S.; Sundram, V.; Chauhan, N.; Gupta, B.K.; Puumala, S.E.; Jaggi, M.; Chauhan, S.C. Novel curcumin-loaded magnetic nanoparticles for pancreatic cancer treatment. Mol. Cancer Ther. 2013, 12, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Saikia, C.; Das, M.K.; Ramteke, A.; Maji, T.K. Evaluation of folic acid tagged aminated starch/ZnO coated iron oxide nanoparticles as targeted curcumin delivery system. Carbohydr. Polym. 2017, 157, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Barick, K.C.; Ekta; Gawali, S.L.; Sarkar, A.; Kunwar, A.; Priyadarsinicd, K.I.; Hassan, P.A. Pluronic stabilized Fe3O4 magnetic nanoparticles for intracellular delivery of curcumin. RSC Adv. 2016, 6, 98674–98681. [Google Scholar] [CrossRef]
- Ayubi, M.; Karimi, M.; Abdpour, S.; Rostamizadeh, K.; Parsa, M.; Zamani, M.; Saedi, A. Magnetic nanoparticles decorated with PEGylated curcumin as dual targeted drug delivery: Synthesis, toxicity and biocompatibility study. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 104, 109810. [Google Scholar] [CrossRef] [PubMed]
- Gangwar, R.K.; Tomar, G.B.; Dhumale, V.A.; Zinjarde, S.; Sharma, R.B.; Datar, S. Curcumin conjugated silica nanoparticles for improving bioavailability and its anticancer applications. J. Agric. Food Chem. 2013, 61, 9632–9637. [Google Scholar] [CrossRef]
- Manju, S.; Sreenivasan, K. Gold nanoparticles generated and stabilized by water soluble curcumin-polymer conjugate: Blood compatibility evaluation and targeted drug delivery onto cancer cells. J. Colloid Interface Sci. 2012, 368, 144–151. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and challenges of liposome assisted drug delivery. Front. Pharm. 2015, 6, 286. [Google Scholar] [CrossRef]
- Li, R.; Deng, L.; Cai, Z.; Zhang, S.; Wang, K.; Li, L.; Ding, S.; Zhou, C. Liposomes coated with thiolated chitosan as drug carriers of curcumin. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 80, 156–164. [Google Scholar] [CrossRef]
- Huang, M.G.; Liang, C.P.; Tan, C.; Huang, S.; Ying, R.F.; Wang, Y.S.; Wang, Z.J.; Zhang, Y.F. Liposome co-encapsulation as a strategy for the delivery of curcumin and resveratrol. Food Funct. 2019, 10, 6447–6458. [Google Scholar] [CrossRef]
- Cheng, C.; Peng, S.; Li, Z.; Zou, L.; Liu, W.; Liu, C. Improved bioavailability of curcumin in liposomes prepared using a pH-driven, organic solvent-free, easily scalable process. RSC Adv. 2017, 7, 25978–25986. [Google Scholar] [CrossRef]
- Jin, H.H.; Lu, Q.; Jiang, J.G. Curcumin liposomes prepared with milk fat globule membrane phospholipids and soybean lecithin. J. Dairy Sci. 2016, 99, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, F.; Cofelice, M.; Venditti, F.; Ceglie, A.; Miguel, M.; Lindman, B.; Lopez, F. In-vitro digestion of curcumin loaded chitosan-coated liposomes. Colloids Surf. B Biointerfaces 2018, 168, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Hamano, N.; Bottger, R.; Lee, S.E.; Yang, Y.; Kulkarni, J.A.; Ip, S.; Cullis, P.R.; Li, S.D. Robust microfluidic technology and new lipid composition for fabrication of curcumin-loaded liposomes: Effect on the anticancer activity and safety of cisplatin. Mol. Pharm. 2019, 16, 3957–3967. [Google Scholar] [CrossRef]
- Thangapazham, R.L.; Puri, A.; Tele, S.; Blumenthal, R.; Maheshwari, R.K. Evaluation of a nanotechnology-based carrier for delivery of curcumin in prostate cancer cells. Int. J. Oncol. 2008, 32, 1119–1123. [Google Scholar] [CrossRef]
- Dhule, S.S.; Penfornis, P.; Frazier, T.; Walker, R.; Feldman, J.; Tan, G.; He, J.; Alb, A.; John, V.; Pochampally, R. Curcumin-loaded gamma-cyclodextrin liposomal nanoparticles as delivery vehicles for osteosarcoma. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 440–451. [Google Scholar] [CrossRef]
- Tefas, L.R.; Sylvester, B.; Tomuta, I.; Sesarman, A.; Licarete, E.; Banciu, M.; Porfire, A. Development of antiproliferative long-circulating liposomes co-encapsulating doxorubicin and curcumin, through the use of a quality-by-design approach. Drug Des. Dev. Ther. 2017, 11, 1605–1621. [Google Scholar] [CrossRef]
- Xu, H.; Gong, Z.; Zhou, S.; Yang, S.; Wang, D.; Chen, X.; Wu, J.; Liu, L.; Zhong, S.; Zhao, J.; et al. Liposomal curcumin targeting endometrial cancer through the NF-kappaB pathway. Cell. Physiol. Biochem. 2018, 48, 569–582. [Google Scholar] [CrossRef]
- Swami Vetha, B.S.; Oh, P.S.; Kim, S.H.; Jeong, H.J. Curcuminoids encapsulated liposome nanoparticles as a blue light emitting diode induced photodynamic therapeutic system for cancer treatment. J. Photochem. Photobiol. B: Biol. 2020, 205, 111840. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Qiu, Q.; Luo, X.; Liu, X.; Sun, J.; Wang, C.; Lin, X.; Deng, Y.; Song, Y. Phyto-phospholipid complexes (phytosomes): A novel strategy to improve the bioavailability of active constituents. Asian J. Pharm. Sci. 2019, 14, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Marczylo, T.H.; Verschoyle, R.D.; Cooke, D.N.; Morazzoni, P.; Steward, W.P.; Gescher, A.J. Comparison of systemic availability of curcumin with that of curcumin formulated with phosphatidylcholine. Cancer Chemother. Pharmacol. 2007, 60, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Maiti, K.; Mukherjee, K.; Gantait, A.; Saha, B.P.; Mukherjee, P.K. Curcumin-phospholipid complex: Preparation, therapeutic evaluation and pharmacokinetic study in rats. Int. J. Pharm. 2007, 330, 155–163. [Google Scholar] [CrossRef]
- Stohs, S.J.; Ji, J.; Bucci, L.R.; Preuss, H.G. A comparative pharmacokinetic assessment of a novel highly bioavailable curcumin formulation with 95% curcumin: A randomized, double-blind, crossover study. J. Am. Coll. Nutr. 2018, 37, 51–59. [Google Scholar] [CrossRef]
- Cuomo, J.; Appendino, G.; Dern, A.S.; Schneider, E.; McKinnon, T.P.; Brown, M.J.; Togni, S.; Dixon, B.M. Comparative absorption of a standardized curcuminoid mixture and its lecithin formulation. J. Nat. Prod. 2011, 74, 664–669. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Zhang, L.; He, D.; Ju, J.; Li, W. Studies on the curcumin phospholipid complex solidified with Soluplus®. J. Pharm. Pharm. 2018, 70, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, M.; Pescina, S.; Padula, C.; Santi, P.; Del Favero, E.; Cantu, L.; Nicoli, S. Polymeric micelles in drug delivery: An insight of the techniques for their characterization and assessment in biorelevant conditions. J. Control Release 2021, 332, 312–336. [Google Scholar] [CrossRef] [PubMed]
- Kheiri Manjili, H.; Ghasemi, P.; Malvandi, H.; Mousavi, M.S.; Attari, E.; Danafar, H. Pharmacokinetics and in vivo delivery of curcumin by copolymeric mPEG-PCL micelles. Eur. J. Pharm. Biopharm. 2017, 116, 17–30. [Google Scholar] [CrossRef]
- Letchford, K.; Liggins, R.; Burt, H. Solubilization of hydrophobic drugs by methoxy poly(ethylene glycol)-block-polycaprolactone diblock copolymer micelles: Theoretical and experimental data and correlations. J. Pharm. Sci. 2008, 97, 1179–1190. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Shayeganpour, A.; Brocks, D.R.; Lavasanifar, A.; Samuel, J. High-performance liquid chromatography analysis of curcumin in rat plasma: Application to pharmacokinetics of polymeric micellar formulation of curcumin. Biomed. Chromatogr. BMC 2007, 21, 546–552. [Google Scholar] [CrossRef]
- Schiborr, C.; Kocher, A.; Behnam, D.; Jandasek, J.; Toelstede, S.; Frank, J. The oral bioavailability of curcumin from micronized powder and liquid micelles is significantly increased in healthy humans and differs between sexes. Mol. Nutr. Food Res. 2014, 58, 516–527. [Google Scholar] [CrossRef]
- Frank, J.; Schiborr, C.; Kocher, A.; Meins, J.; Behnam, D.; Schubert-Zsilavecz, M.; Abdel-Tawab, M. Transepithelial transport of curcumin in Caco-2 cells is significantly enhanced by micellar solubilisation. Plant Foods Hum. Nutr. 2017, 72, 48–53. [Google Scholar] [CrossRef][Green Version]
- Parikh, A.; Kathawala, K.; Song, Y.; Zhou, X.F.; Garg, S. Curcumin-loaded self-nanomicellizing solid dispersion system: Part I: Development, optimization, characterization, and oral bioavailability. Drug Deliv. Transl. Res. 2018, 8, 1389–1405. [Google Scholar] [CrossRef]
- Chen, S.; Li, Q.; Li, H.; Yang, L.; Yi, J.Z.; Xie, M.; Zhang, L.M. Long-circulating zein-polysulfobetaine conjugate-based nanocarriers for enhancing the stability and pharmacokinetics of curcumin. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 109, 110636. [Google Scholar] [CrossRef]
- Song, L.; Shen, Y.Y.; Hou, J.W.; Lei, L.; Guo, S.R.; Qian, C.Y. Polymeric micelles for parenteral delivery of curcumin: Preparation, characterization and in vitro evaluation. Colloid Surf. A 2011, 390, 25–32. [Google Scholar] [CrossRef]
- Patil, S.; Choudhary, B.; Rathore, A.; Roy, K.; Mahadik, K. Enhanced oral bioavailability and anticancer activity of novel curcumin loaded mixed micelles in human lung cancer cells. Phytomed. Int. J. Phytother. Phytopharm. 2015, 22, 1103–1111. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, B.; Chu, L.; Tong, H.H.; Liu, W.; Zhai, G. Evaluation in vitro and in vivo of curcumin-loaded mPEG-PLA/TPGS mixed micelles for oral administration. Colloids Surf. B Biointerfaces 2016, 141, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Hani, U.; Shivakumar, H.G. Solubility enhancement and delivery systems of curcumin a herbal medicine: A review. Curr. Drug Deliv. 2014, 11, 792–804. [Google Scholar] [CrossRef]
- Szejtli, J. Introduction and general overview of cyclodextrin chemistry. Chem. Rev. 1998, 98, 1743–1754. [Google Scholar] [CrossRef]
- Bakshi, P.R.; Londhe, V.Y. Widespread applications of host-guest interactive cyclodextrin functionalized polymer nanocomposites: Its meta-analysis and review. Carbohydr. Polym. 2020, 242, 116430. [Google Scholar] [CrossRef]
- Mangolim, C.S.; Moriwaki, C.; Nogueira, A.C.; Sato, F.; Baesso, M.L.; Neto, A.M.; Matioli, G. Curcumin-beta-cyclodextrin inclusion complex: Stability, solubility, characterisation by FT-IR, FT-Raman, X-ray diffraction and photoacoustic spectroscopy, and food application. Food Chem. 2014, 153, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Guo, S. Encapsulation of curcumin into β-cyclodextrins inclusion: A review. E3S Web Conf. 2019, 131, 01100. [Google Scholar] [CrossRef]
- Jantarat, C.; Sirathanarun, P.; Ratanapongsai, S.; Watcharakan, P.; Sunyapong, S.; Wadu, A. Curcumin-hydroxypropyl-β-cyclodextrin inclusion complex preparation methods: Effect of common solvent evaporation, freeze drying, and pH shift on solubility and stability of curcumin. Trop. J. Pharm. Res. 2014, 13, 1215–1223. [Google Scholar] [CrossRef]
- Li, N.; Wang, N.; Wu, T.; Qiu, C.; Wang, X.; Jiang, S.; Zhang, Z.; Liu, T.; Wei, C.; Wang, T. Preparation of curcumin-hydroxypropyl-beta-cyclodextrin inclusion complex by cosolvency-lyophilization procedure to enhance oral bioavailability of the drug. Drug Dev. Ind. Pharm. 2018, 44, 1966–1974. [Google Scholar] [CrossRef]
- Ghanghoria, R.; Kesharwani, P.; Agashe, H.B.; Jain, N.K. Transdermal delivery of cyclodextrin-solubilized curcumin. Drug Deliv. Transl. Res. 2013, 3, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Popat, A.; Karmakar, S.; Jambhrunkar, S.; Xu, C.; Yu, C. Curcumin-cyclodextrin encapsulated chitosan nanoconjugates with enhanced solubility and cell cytotoxicity. Colloids Surf. B Biointerfaces 2014, 117, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Man, S.; Qiu, H.; Liu, Z.; Zhang, M.; Ma, L.; Gao, W. Curcumin-cyclodextrin complexes enhanced the anti-cancer effects of curcumin. Environ. Toxicol. Pharmacol. 2016, 48, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Moller, K.; Macaulay, B.; Bein, T. Curcumin encapsulated in crosslinked cyclodextrin nanoparticles enables immediate inhibition of cell growth and efficient killing of cancer cells. Nanomaterials 2021, 11, 489. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Guo, F.; Yu, N.; Ying, S.; Lou, B.; Wu, J.; Gao, Y.; Ji, X.; Wang, H.; Li, A.; et al. A novel folic acid receptor-targeted drug delivery system based on curcumin-loaded beta-cyclodextrin nanoparticles for cancer treatment. Drug Des. Dev. Ther. 2021, 15, 2843–2855. [Google Scholar] [CrossRef]
- Stasilowicz, A.; Tykarska, E.; Lewandowska, K.; Kozak, M.; Miklaszewski, A.; Kobus-Cisowska, J.; Szymanowska, D.; Plech, T.; Jenczyk, J.; Cielecka-Piontek, J. Hydroxypropyl-beta-cyclodextrin as an effective carrier of curcumin–piperine nutraceutical system with improved enzyme inhibition properties. J. Enzym. Inhib. Med. Chem. 2020, 35, 1811–1821. [Google Scholar] [CrossRef]
- Tran, P.H.L.; Tran, T.T.D. Developmental strategies of curcumin solid dispersions for enhancing bioavailability. Anti-Cancer Agents Med. Chem. 2020, 20, 1874–1882. [Google Scholar] [CrossRef]
- Teixeira, C.C.; Mendonca, L.M.; Bergamaschi, M.M.; Queiroz, R.H.; Souza, G.E.; Antunes, L.M.; Freitas, L.A. Microparticles containing curcumin solid dispersion: Stability, bioavailability and anti-inflammatory activity. AAPS PharmSciTech 2016, 17, 252–261. [Google Scholar] [CrossRef]
- Basniwal, R.K.; Khosla, R.; Jain, N. Improving the anticancer activity of curcumin using nanocurcumin dispersion in water. Nutr. Cancer 2014, 66, 1015–1022. [Google Scholar] [CrossRef]
- Seo, S.W.; Han, H.K.; Chun, M.K.; Choi, H.K. Preparation and pharmacokinetic evaluation of curcumin solid dispersion using Solutol® HS15 as a carrier. Int. J. Pharm. 2012, 424, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Sun, Y.; Qi, X.; Tan, F. Improved bioavailability of poorly water-soluble drug curcumin in cellulose acetate solid dispersion. AAPS PharmSciTech 2012, 13, 159–166. [Google Scholar] [CrossRef]
- Mendonca, L.M.; Machado Cda, S.; Teixeira, C.C.; Freitas, L.A.; Bianchi, M.L.; Antunes, L.M. Comparative study of curcumin and curcumin formulated in a solid dispersion: Evaluation of their antigenotoxic effects. Genet. Mol. Biol. 2015, 38, 490–498. [Google Scholar] [CrossRef]
- Silva de Sa, I.; Peron, A.P.; Leimann, F.V.; Bressan, G.N.; Krum, B.N.; Fachinetto, R.; Pinela, J.; Calhelha, R.C.; Barreiro, M.F.; Ferreira, I.; et al. In vitro and in vivo evaluation of enzymatic and antioxidant activity, cytotoxicity and genotoxicity of curcumin-loaded solid dispersions. Food Chem. Toxicol. 2019, 125, 29–37. [Google Scholar] [CrossRef]
- Han, H.K. The effects of black pepper on the intestinal absorption and hepatic metabolism of drugs. Expert Opin. Drug Metab. Toxicol. 2011, 7, 721–729. [Google Scholar] [CrossRef]
- Shoba, G.; Joy, D.; Joseph, T.; Majeed, M.; Rajendran, R.; Srinivas, P.S. Influence of piperine on the pharmacokinetics of curcumin in animals and human volunteers. Planta Med. 1998, 64, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Nehru, B.; Munshi, A.; Jyothy, A. Antioxidant potential of curcumin against oxidative insult induced by pentylenetetrazol in epileptic rats. Methods Find. Exp. Clin. Pharmacol. 2010, 32, 227–232. [Google Scholar] [CrossRef]
- Suresh, D.; Srinivasan, K. Tissue distribution & elimination of capsaicin, piperine & curcumin following oral intake in rats. Indian J. Med. Res. 2010, 131, 682–691. [Google Scholar]
- Antony, B.; Merina, B.; Iyer, V.S.; Judy, N.; Lennertz, K.; Joyal, S. A pilot cross-over study to evaluate human oral bioavailability of BCM-95®CG (Biocurcumax™), a novel bioenhanced preparation of curcumin. Indian J. Pharm. Sci. 2008, 70, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.V.; Agarwal, S.; Singh, P.; Godbole, M.M.; Misra, K. Curcumin conjugates induce apoptosis via a mitochondrion dependent pathway in MCF-7 and MDA-MB-231 cell lines. Asian Pac. J. Cancer Prev. APJCP 2013, 14, 5797–5804. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.R.; Qian, Z.Y. Drug delivery systems for overcoming the bioavailability of curcumin: Not only the nanoparticle matters. Nanomed. Nanotechnol. Biol. Med. 2014, 9, 747–750. [Google Scholar] [CrossRef]
- Guo, G.; Fu, S.; Zhou, L.; Liang, H.; Fan, M.; Luo, F.; Qian, Z.; Wei, Y. Preparation of curcumin loaded poly(epsilon-caprolactone)-poly(ethylene glycol)-poly(epsilon-caprolactone) nanofibers and their in vitro antitumor activity against Glioma 9L cells. Nanoscale 2011, 3, 3825–3832. [Google Scholar] [CrossRef]
- Wang, C.; Ma, C.; Wu, Z.K.; Liang, H.; Yan, P.; Song, J.; Ma, N.; Zhao, Q.H. Enhanced bioavailability and anticancer effect of curcumin-loaded electrospun nanofiber: In vitro and in vivo study. Nanoscale Res. Lett. 2015, 10, 439. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Xu, Y.; Zhou, X.; Gong, T.; Zhang, Z.R.; Fu, Y. An injectable micelle-hydrogel hybrid for localized and prolonged drug delivery in the management of renal fibrosis. Acta Pharm. Sin. B 2021, 11, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Stojanovic-Radic, Z.; Matejic, J.; Sharifi-Rad, M.; Anil Kumar, N.V.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of curcumin: A review of clinical trials. Eur. J. Med. Chem. 2019, 163, 527–545. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef]
- Cheng, A.L.; Hsu, C.H.; Lin, J.K.; Hsu, M.M.; Ho, Y.F.; Shen, T.S.; Ko, J.Y.; Lin, J.T.; Lin, B.R.; Ming-Shiang, W.; et al. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res. 2001, 21, 2895–2900. [Google Scholar]
- Dhillon, N.; Aggarwal, B.B.; Newman, R.A.; Wolff, R.A.; Kunnumakkara, A.B.; Abbruzzese, J.L.; Ng, C.S.; Badmaev, V.; Kurzrock, R. Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin. Cancer Res. 2008, 14, 4491–4499. [Google Scholar] [CrossRef]
- Mahammedi, H.; Planchat, E.; Pouget, M.; Durando, X.; Cure, H.; Guy, L.; Van-Praagh, I.; Savareux, L.; Atger, M.; Bayet-Robert, M.; et al. The new combination docetaxel, prednisone and curcumin in patients with castration-resistant prostate cancer: A pilot phase II study. Oncology 2016, 90, 69–78. [Google Scholar] [CrossRef]
- Dutzmann, S.; Schiborr, C.; Kocher, A.; Pilatus, U.; Hattingen, E.; Weissenberger, J.; Gessler, F.; Quick-Weller, J.; Franz, K.; Seifert, V.; et al. Intratumoral concentrations and effects of orally administered micellar curcuminoids in glioblastoma patients. Nutr. Cancer 2016, 68, 943–948. [Google Scholar] [CrossRef]
- Sharma, R.A.; Euden, S.A.; Platton, S.L.; Cooke, D.N.; Shafayat, A.; Hewitt, H.R.; Marczylo, T.H.; Morgan, B.; Hemingway, D.; Plummer, S.M.; et al. Phase I clinical trial of oral curcumin: Biomarkers of systemic activity and compliance. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 6847–6854. [Google Scholar] [CrossRef]
- Carroll, R.E.; Benya, R.V.; Turgeon, D.K.; Vareed, S.; Neuman, M.; Rodriguez, L.; Kakarala, M.; Carpenter, P.M.; McLaren, C.; Meyskens, F.L., Jr.; et al. Phase IIa clinical trial of curcumin for the prevention of colorectal neoplasia. Cancer Prev. Res. 2011, 4, 354–364. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Biological Effects | Mechanisms of action | Cancer type | References |
|---|---|---|---|
| Antiproliferative | |||
| CDK2 decrease | Breast | [35] | |
| CDK4 decrease | [34,35] | ||
| Cell cycle arrest at G1 phase | [16,35,36] | ||
| Cell cycle arrest at G2/M phase | [37] | ||
| Cell viability decrease | [39] | ||
| Cyclin A decrease | [35] | ||
| Cyclin D1 decrease | [34,35,37] | ||
| Cyclin E decrease | [35,36] | ||
| DLC1 increase | [40] | ||
| EZH2 decrease | [40] | ||
| GSK3β increase | [37] | ||
| Increased sensibility to chemotherapeutic agents | [39] | ||
| IκB increase | [16] | ||
| NF-kB p65 decrease | [16] | ||
| p21 and p27 increase | [35,36] | ||
| p53 increase | [36] | ||
| β-catenin decrease | [37] | ||
| CDK2 decrease | Colon | [45] | |
| Cell cycle arrest at G1 phase | [45] | ||
| Cell cycle arrest at G2/M phase | [47,48] | ||
| Cell viability decrease | [44] | ||
| Cyclin A decrease | [46] | ||
| E2F4, decrease | [46] | ||
| p21 and p27 increase | [46] | ||
| p53 increase | [47] | ||
| ROS increase | [46] | ||
| Cell cycle arrest at G2/M phase | Bladder | [49] | |
| Cell viability decrease | [49,50] | ||
| Cyclin E1 decrease | [49] | ||
| p27 increase | [49] | ||
| Trop2 decrease | [49] | ||
| Cell cycle arrest at G2/M phase | Glioma | [53] | |
| Cyclin D1 decrease | [52] | ||
| Egr-1 increase | [52] | ||
| FoxO1 increase | [53] | ||
| p21 increase | [52] | ||
| Cell cycle arrest at G2/M phase | Glioblastoma | [54] | |
| HDGF / β-catenin complex inhibition | [56] | ||
| p57 increase | [54] | ||
| Skp2 decrease | [54] | ||
| CDK4 decrease | Gastric | [67] | |
| Cell cycle arrest at G1 phase | [60] | ||
| Cell cycle arrest at G0/G1-S phase | [67] | ||
| c-myc decrease | [61] | ||
| c-myc/(lncRNA) H19 pathway downregulation | [63] | ||
| Cyclin D1 decrease | [67] | ||
| EGF/PAK1/NF-kB/cyclin D1 pathway inhibition | [60] | ||
| LRP6 ans phospho-LRP6 decrease | [61] | ||
| miR-33b increase | [68] | ||
| miR-34a increase | [67] | ||
| ODC activity decrease | [59] | ||
| p21 increase | [62] | ||
| p53 increase | [62,63] | ||
| PI3K signaling inhibition | [62] | ||
| SMOX mRNA and activity increase | [59] | ||
| Wnt3a decrease | [61] | ||
| XIAP decrease | [68] | ||
| β-catenin and phospho β-catenin decrease | [61] | ||
| Akt/mTOR pathway downregulation | Melanoma | [72] | |
| Cell cycle arrest at G2/M phase | [70,71,72] | ||
| Cyclin A decrease | [71] | ||
| DNMT1 decrease | [71] | ||
| IKK inhibition | [73] | ||
| iNOS inhibition | [70] | ||
| NF-kB inhibition | [70,73] | ||
| p21 and p27 increase | [71] | ||
| PDE decrease | [71] | ||
| UHRF1 decrease | [71] | ||
| Pro-apoptotic | |||
| Bad increase | Breast | [77] | |
| Bax increase | [79] | ||
| Bax/Bcl-2 ratio increase | [78] | ||
| Bcl-2 decrease | [79,94] | ||
| Cleaved caspase 3 increase | [78] | ||
| Cleaved Parp-1 increase | [77] | ||
| FAS inhibition | [80] | ||
| GSH decrease | [78] | ||
| Histone H3 acetylation and glutathionylation increase | [74] | ||
| miR-15a and miR-16 increase | [94] | ||
| NF-kBp65 decrease | [79] | ||
| p53 increase | [77] | ||
| ROS increase | [78] | ||
| Bcl-2 decrease | Melanoma | [81] | |
| JAK-2/STAT-3 signaling inhibition | [81] | ||
| p53-independent Fas/caspase 8 pathway activation | [82] | ||
| Akt signaling inhibition | Ovarian | [83] | |
| Bcl-2 and survivin decrease | [83] | ||
| p38 MAPK activation | [83] | ||
| Bax and p53 increase | Myeloma | [84] | |
| MDM2 decrease | [84] | ||
| AMPK increase | Colon | [85] | |
| APC decrease | [87] | ||
| Bax increase | [89,90,91] | ||
| Bcl-2 decrease | [86,89,90,91] | ||
| Bcl-xL decrease | [86] | ||
| Caspase 3 activation | [87,89] | ||
| Caspase 7 activation | [89] | ||
| COX2 decrease | [86] | ||
| Cyclin D1 decrease | [86] | ||
| DR5 upregulation | [88] | ||
| E-cadherin decrease | [87] | ||
| Fas-mediated caspase 8 activation | [89] | ||
| IAP-2 decrease | [86] | ||
| Mithocondrial [Ca2+] increase | [89] | ||
| Mithocondrial cytochrome c release | [89] | ||
| Mithocondrial membrane potential reduction | [89] | ||
| pAKT decrease | [85] | ||
| β-catenin decrease | [87] | ||
| Caspase 3/7 activation | Bladder | [50] | |
| Caspase 3/7 activation | Glioblastoma | [92] | |
| Bax increase | [92] | ||
| Bcl-2 decrease | [92] | ||
| Caspase 3 activation | Hosteosarcoma | [93] | |
| Parp-1 cleavage | [93] | ||
| DNA damage | Gastric | [59] | |
| ODC activity decrease | [59] | ||
| ROS production | [59] | ||
| miR-186 pathway activation | Lung | [95] | |
| Bax and cleaved caspase 3/9 increase | Bladder | [96] | |
| Bcl2 decrease | [96] | ||
| miRNA 344a-3p increase | [96] | ||
| Antimetastatic | |||
| Axl decrease | Breast | [98] | |
| CD24 decrease | [98] | ||
| CXCL1 and 2 decrease | [101] | ||
| miR-34a increase | [98] | ||
| NF-kB inhibition | [100,101] | ||
| Rho-A decrease | [98] | ||
| RhoA/ROCK/MMPs/Vimentin pathway inhibition | [103] | ||
| Slug decrease | [98] | ||
| uPA decrease | [100] | ||
| JAK/STAT3 pathway inhibition | Retinoblastoma | [104] | |
| miR-99a increase | [104] | ||
| MMP2 decrease | [104] | ||
| RhoA decrease | [104] | ||
| ROCK1 decrease | [104] | ||
| Vimentin decrease | [104] | ||
| MMPs signaling pathways inhibition | Bladder | [50] | |
| Trop2 decrease | [49] | ||
| Cellular matriptase downregulation | Prostate | [106] | |
| MMP9 decrease | [106] | ||
| Angiogenesis inhibition | Colon | [44] | |
| Claudin-3 decrease | [107] | ||
| Metastasis inhibition | [44] | ||
| MMP9 decrease | [107] | ||
| MMP2 decrease | Melanoma | [81,108] | |
| MMP9 decrease | [81] | ||
| NF-kB signaling pathways inhibition | [108] | ||
| TIMP-2 increase | [81] | ||
| NF-kB and Wnt/βcatenin pathways inhibition | Cervical | [17] | |
| ICAM decrease | SCLC | [110] | |
| VEGF decrease | [110] | ||
| MMP2 and MMP7 decrease | [110] | ||
| STAT3 decrease | [110] | ||
| IL-6-inducible JAK/STAT3 phosphorylation reduction | [110] | ||
| Fascin decrease | Ovarian | [15] | |
| JAK/STAT3 signaling pathway inhibition | [15] | ||
| pAkt, pmTOR, pP70S6K downregulation | Melanoma | [72] | |
| miR-27a decrease | Thymic | [111] | |
| mTOR and Notch-1 pathways inhibition | [111] | ||
| miR-206 increase | NSCLC | [112] | |
| PI3K/AKT/mTOR pathway inhibition | [112] | ||
| NEDD4 signaling pathways inhibition | Glioma | [113] | |
| Composition | Outcomes | References |
|---|---|---|
| Polymeric Nanoparticles (NPs) | ||
| PLGA + CUR | Superior anticancer effects respect to free curcumin in cervical cancer cells | [160] |
| PLGA + CUR | Superior cellular uptake and anticancer effects respect to free curcumin in ovarian and breast cancer cells | [161] |
| PLGA + CUR | Superior cellular uptake and anticancer effects respect to free curcumin in prostate cancer cells | [162] |
| NIPAAM+ VP + PEG-A + CUR | Superior cellular absorbition and anticancer effects respect to free curcumin in pancreatic cancer cells | [163] |
| mPEG-PCL + CUR | Superior anticancer effects respect to free curcumin in human lung adenocarcinoma cancer cells | [164] |
| silk fibroin + CUR | Superior cellular uptake and anticancer effects respect to free curcumin in colon cancer cells | [165] |
| zein-chitosan +CUR | High encapsulation efficiencies for curcumin. Superior anticancer effects respect to free curcumin in neuroblastoma cell line | [166] |
| chitosan +CUR | Enhanced curcumin solubility and bioavailability. Sustained drug release from NPs and anticancer effects with respect to free curcumin in cervical cancer cells | [167] |
| Solid Lipid Nanoparticles (SLNs) | ||
| SLNs + N-carboxymethyl chitosan+ CUR | Prolonged release in simulated intestinal fluid, greater absorption and oral bioavailability compared to free curcumin. Superior anticancer effects respect to free curcumin in breast cancer cells | [171] |
| SLNs + CUR, d-α-Tocopheryl polyethylene glycol 1000 succinate-stabilized curcumin (TPGS) + CUR | Superior curcumin plasma levels in mice. Superior anticancer effects respect to free curcumin in Hodgkin lymphoma cells and in Hodgkin’s lymphoma xenograft models | [172] |
| SLNs + tristearin + PEGylated + CUR | Superior bioavailability, absorption and long-term stability after oral administration in the rats | [174] |
| SLNs + NaCas + NaCas-Lac + CUR | Superior stability at pH acid and antioxidant activity with respect to free curcumin | [175] |
| SLNs + glyceryl monostearate + poloxamer 188 + CUR | Superior stability, solubility, cellular uptake, release and anticancer effects respect to free curcumin in breast cancer cells | [176] |
| Inorganic Nanoparticles | ||
| MNPs + CUR | Superior cellular uptake in vitro. Superior bioavailability in vivo. Superior in vitro and in vivo therapeutic efficacy respect to free curcumin in pancreatic cancer cells and in pancreatic cancer xenografts model | [180] |
| Folic-acid-tagged aminated-starch-/ZnO-coated iron oxide nanoparticles + CUR | Significant controlled release of curcumin and reduced hepatic and breast cancer cells viability in vitro. Cellular uptake increase in vitro | [181] |
| PSMNPs + CUR | Aqueous colloidal stability, biocompatibility, high loading affinity for curcumin and better curcumin release in acidic conditions. Superior cellular uptake and anticancer effects respect to free curcumin in breast cancer cells | [182] |
| MNP@PEG + CUR | Higher drug release in acidic conditions, biocompatibility and low cytotoxicity at physiological pH | [183] |
| Silica + CUR | Good stability in aqueous medium, sustained drug release and greater anticancer properties in cervical cancer cells compared to normal fibroblasts | [184] |
| HA-CUR@AuNPs | Good aqueous solubility, superior cellular uptake and anticancer effects respect to free curcumin in cervical, glioma and colon cancer cells | [185] |
| Liposomes | ||
| Liposome + CUR | Improved curcumin aqueous solubility and bioavailability in tumor-bearing mice | [192] |
| DMPC + CUR | Superior anticancer effects respect to free curcumin in prostate cancer cells | [193] |
| 2-hydroxypropyl-γ-cyclodextrin/ liposome + CUR | Superior in vitro and in vivo anticancer effects respect to free curcumin in osteosarcoma cancer cells | [194] |
| Liposome + doxorubicin + CUR | Superior anticancer effects respect to those loaded with doxorubicin alone in colon cancer cells | [195] |
| Liposome + CUR | Superior anticancer effects respect to free curcumin in endometrial cancer cells | [196] |
| Liposome + CUR + BLED-PDT therapy | Enhancement of BLED-PDT therapy effect by curcumin liposome in lung cancer cells | [197] |
| Phytosomes | ||
| Curcuminoids + lecithin (Meriva ®) | Improved absorption and clinical efficacy respect to unformulated curcuminoid mixtures | [198] |
| Soluplus® [polyvinyl caprolactam-polyvinyl acetate polyethylene glycol graft copolymer] + CPC | Improved flowability, dissolution rate and oral bioavailability in rats | [203] |
| Micelles | ||
| MePEG-b-PCL + CUR | Improved water solubility | [206] |
| MePEG-b-PCL + CUR | Improved biological half-life with respect to the free curcumin in rat models | [207] |
| Tween-80 micelles + CUR | Improved drug plasma concentration with respect to free curcumin in volunteers | [208] |
| Micellar formulation + CUR (Sol-CUR) | Superior uptake, transepithelial transport, distribution and bioavailability in colon cancer cell model | [209] |
| Micelles + CUR | Enhancement of aqueous solubility, stability, dissolution and permeability of curcumin formulated in micelles compared to free drug | [210] |
| PSBMA + CUR | Greater stability, cellular uptake and tumor cytotoxicity compared to free curcumin | [211] |
| MPEG-P [CL-co-PDO] + CUR | Superior encapsulation efficiency, prolonged drug release profile and antitumor effects respect to free curcumin in prostate cancer cells | [212] |
| Pluronic F-127 + Gelucire® 44/14 micelles + CUR | Controlled curcumin release, superior oral bioavailability in vivo and in vitro antitumor effects in lung cancer cells respect to free curcumin | [213] |
| CUR-MPP-TPGS-MMs | Small size, high drug-loading and sustained release. Improved intestinal absorption and oral bioavailabil-ity in rats | [214] |
| Curcumin/β-Cyclodextrin (β-CD) and Solid Dispersions (SDs) | ||
| β-CD + CUR | Superior sunlight stability and solubility respect to pure colourant | [218] |
| Liquid-type β-CD + CUR | Improved solubility in water and bioavailability | [220,221] |
| Solid type β-CD + CUR | Improved storage stability and biovailability | [222] |
| CUR-CD-CS | Superior solubility, cellular absorption and antitumor effects compared to free curcumin in skin cancer cells | [223] |
| β-CD + CUR | Improved uptake and therapeutic efficacy in prostate cancer cells | [154] |
| β-CD + CUR | Improved delivery and therapeutic efficacy compared to free curcumin in both in vitro lung carcinoma cell lines and in vivo mouse hepatoma xenograft models | [224] |
| β-CD + CUR | Superior anticancer effects respect to free curcumin in cervical cancer cells | [225] |
| FA-CUR-NPs | Improved drug release rate, cellular uptake efficiency and in vitro and in vivo antitumor activity respect to free curcumin in cervical cancer cells | [226] |
| Hydroxypropyl-β-CD+ CUR+piperine | Improved solubility of the curcumin–piperine system, its permeability through biological membranes, antioxidant and antimicrobial activities and enzymatic inhibition | [227] |
| SDs (with Gelucire®50/13-Aerosil) + CUR | Improved stability, water solubility, dissolution rate, bioavailability and anti-inflammatory activity in rats | [229] |
| Solutol® HS15 SDs + CUR | Improved solubility and bioavailability compared to free curcumin | [231] |
| SDs (with cellulose acetate and mannitol) + CUR | Improved water solubility and oral bioavailability compared to free curcumin | [232] |
| SDs + CUR | Superior water solubility and gastrointestinal absorption in rats | [233] |
| SDs (with Poloxamer 407) + CUR | Improved water dispersibility and cytotoxic effects against breast, lung, cervical and hepatocellular cancer cells | [234] |
| Curcumin Conjugates Formulations | ||
| Piperine + CUR | Curcumin enhanced serum levels, reduced elimination half-life and clearance, increased bioavailability in rats and humans | [236] |
| Piperine + CUR | Curcumin increased bioavailability in epileptic rats | [237] |
| Piperine + CUR | Curcumin enhanced intestinal absorption and bioavailability in rats | [238] |
| BCM-95CG (Biocurcumax®): piperine + lecithin + CUR | Curcumin improved bioavailability and pharmacokinetic profile respect to free drug in healthy subjects | [239] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nocito, M.C.; De Luca, A.; Prestia, F.; Avena, P.; La Padula, D.; Zavaglia, L.; Sirianni, R.; Casaburi, I.; Puoci, F.; Chimento, A.; et al. Antitumoral Activities of Curcumin and Recent Advances to ImProve Its Oral Bioavailability. Biomedicines 2021, 9, 1476. https://doi.org/10.3390/biomedicines9101476
Nocito MC, De Luca A, Prestia F, Avena P, La Padula D, Zavaglia L, Sirianni R, Casaburi I, Puoci F, Chimento A, et al. Antitumoral Activities of Curcumin and Recent Advances to ImProve Its Oral Bioavailability. Biomedicines. 2021; 9(10):1476. https://doi.org/10.3390/biomedicines9101476
Chicago/Turabian StyleNocito, Marta Claudia, Arianna De Luca, Francesca Prestia, Paola Avena, Davide La Padula, Lucia Zavaglia, Rosa Sirianni, Ivan Casaburi, Francesco Puoci, Adele Chimento, and et al. 2021. "Antitumoral Activities of Curcumin and Recent Advances to ImProve Its Oral Bioavailability" Biomedicines 9, no. 10: 1476. https://doi.org/10.3390/biomedicines9101476
APA StyleNocito, M. C., De Luca, A., Prestia, F., Avena, P., La Padula, D., Zavaglia, L., Sirianni, R., Casaburi, I., Puoci, F., Chimento, A., & Pezzi, V. (2021). Antitumoral Activities of Curcumin and Recent Advances to ImProve Its Oral Bioavailability. Biomedicines, 9(10), 1476. https://doi.org/10.3390/biomedicines9101476