
Longitudinal Pathogenic Properties and N-Glycosylation Profile of Antibodies from Patients with Pemphigus after Corticosteroid Treatment

 , , , , , , , , ,
, , , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Sera and IgG Purification
2.3. Antibodies
2.4. In Vitro Modification of the N-Glycans of Purified IgG
2.5. Eastern Blot
2.6. Dispase-Based Dissociation Assay
2.7. Immunofluorescence Assays
2.8. Mass Spectrometry Analysis of IgG N-Glycans
2.9. Statistical Analysis
3. Results
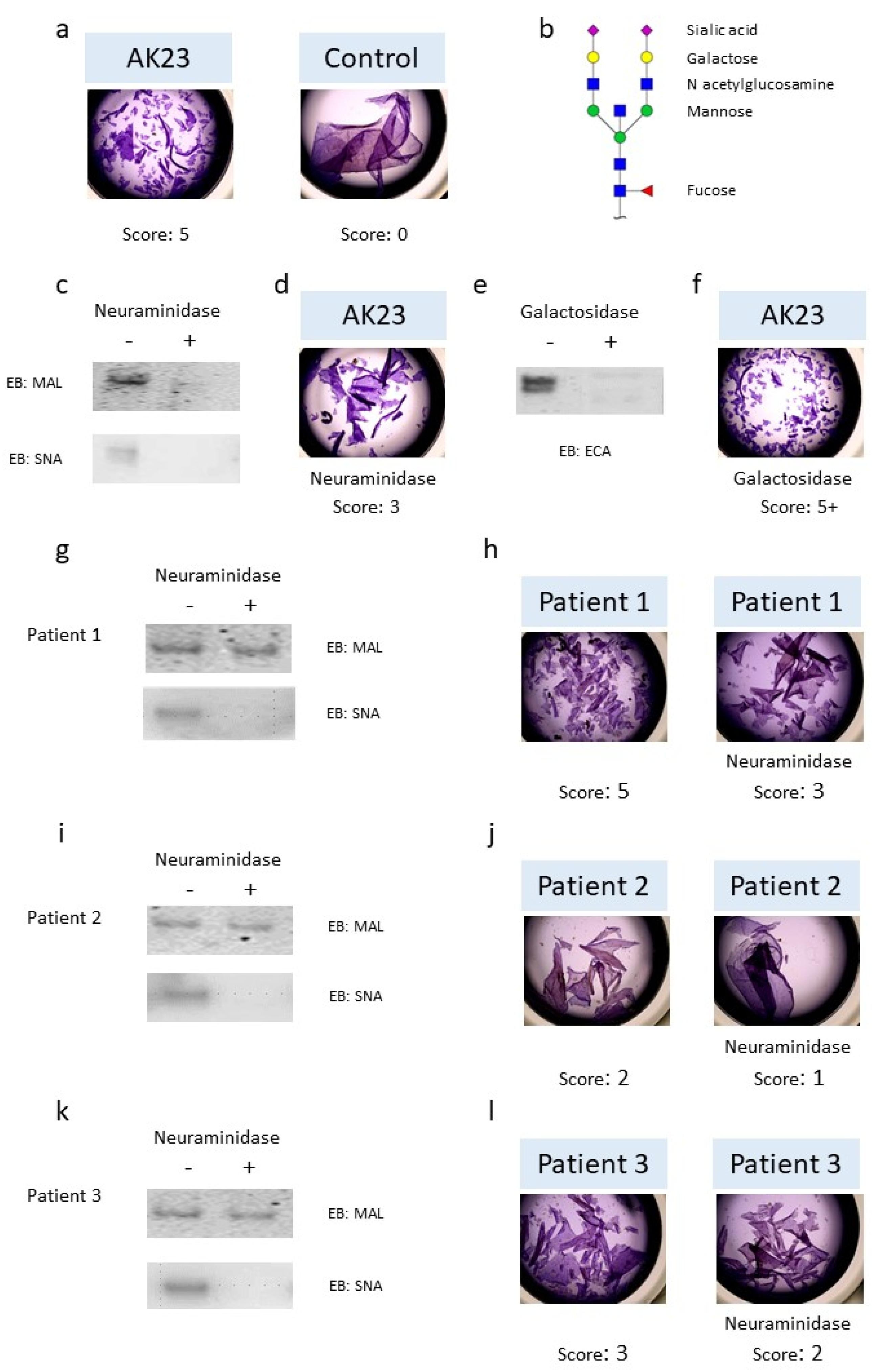
3.1. Role of N-Glycans in IgG Pathogenic Activity
3.2. Pathogenic Activity of IgG Antibodies from Patients with Pemphigus
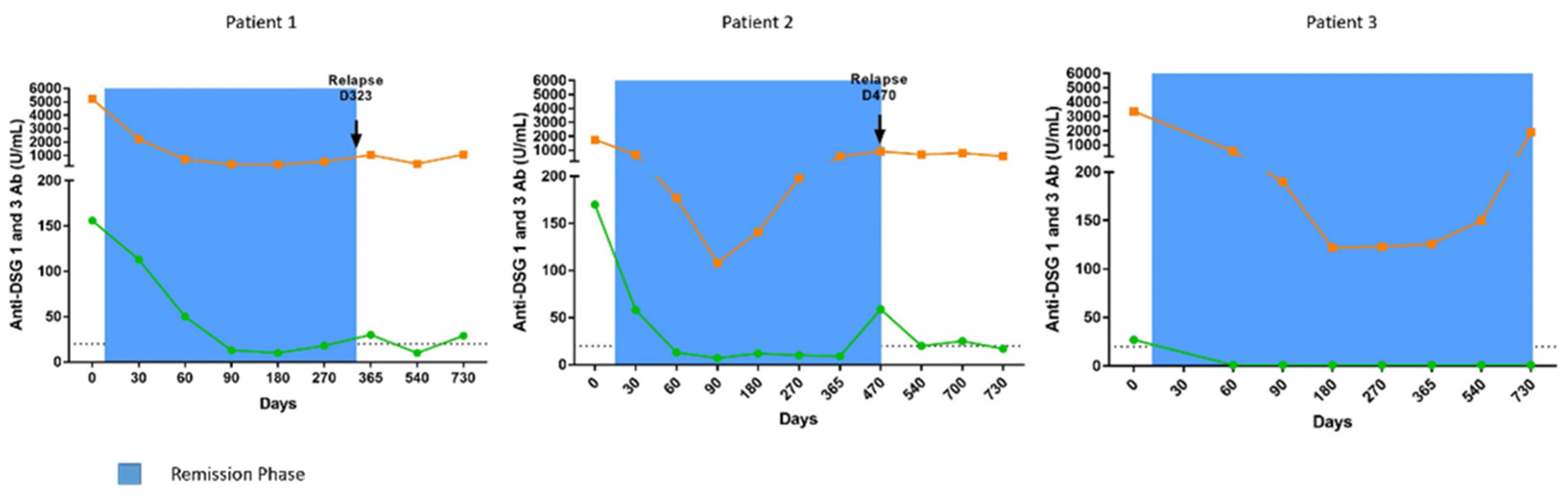
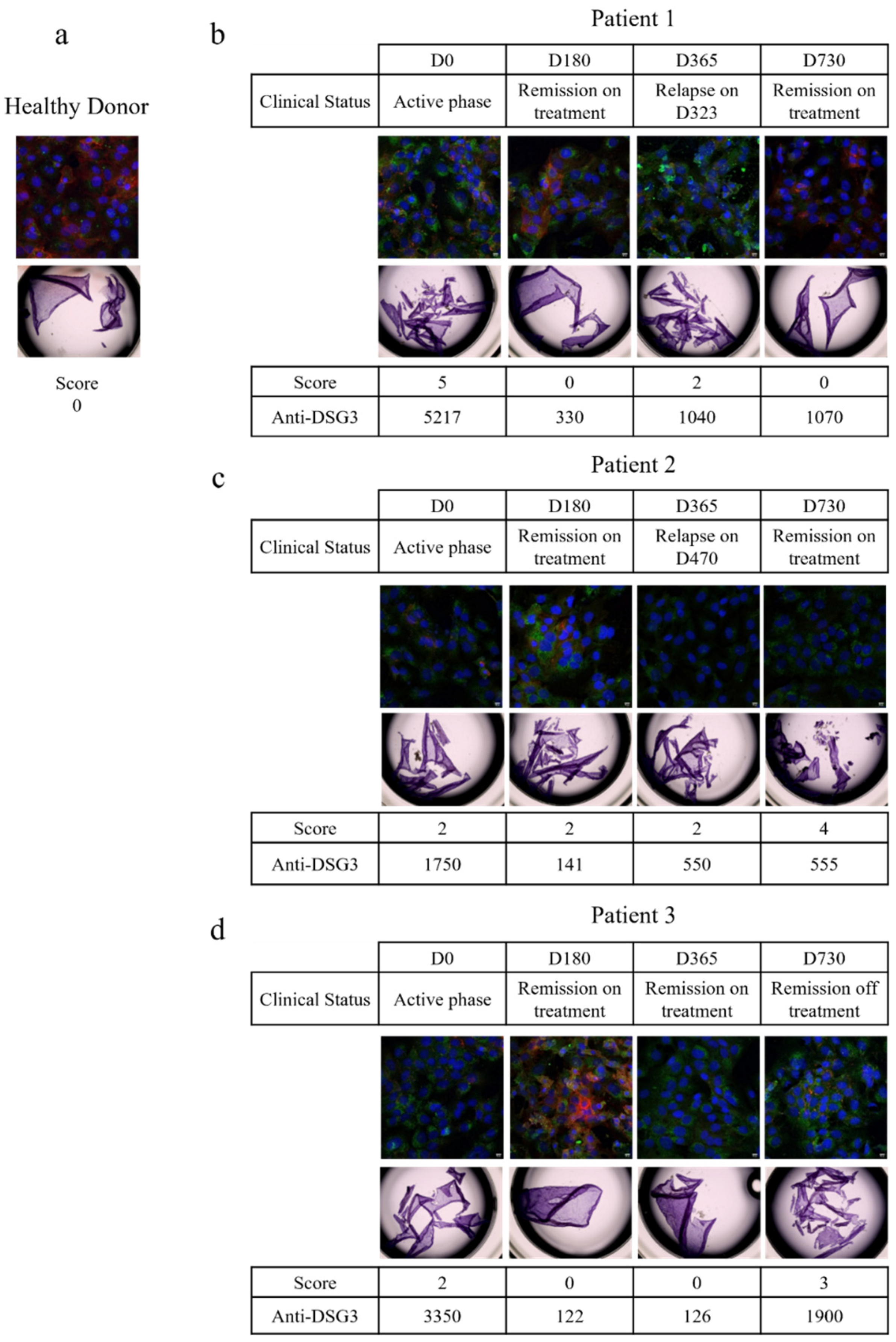
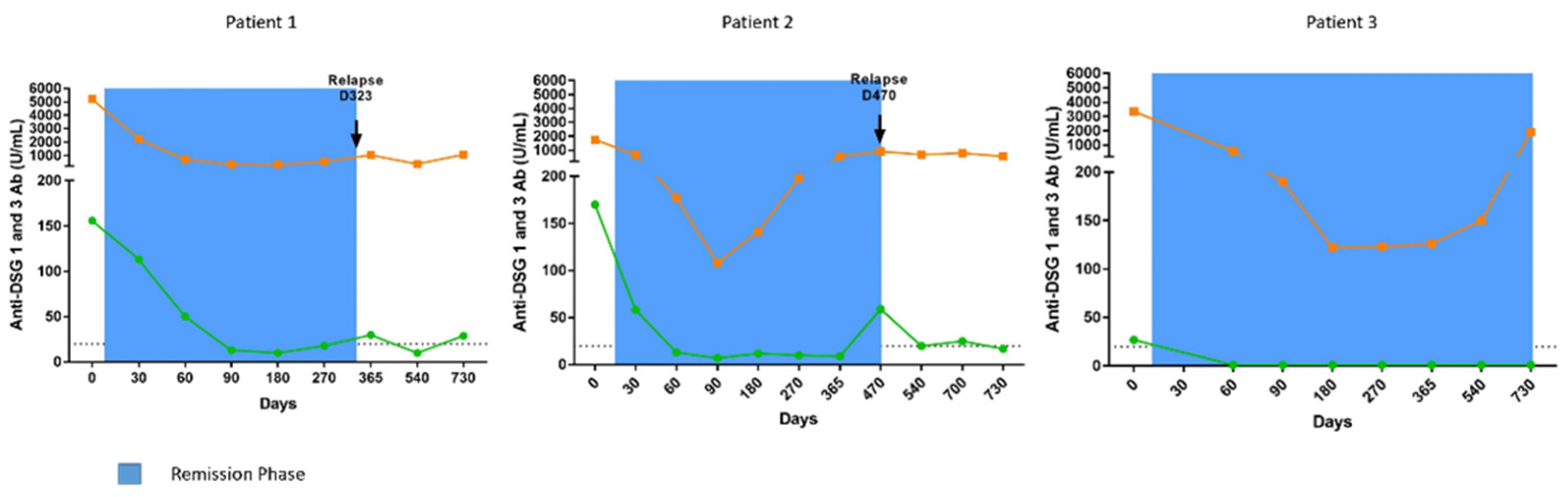
3.2.1. Patient 1
3.2.2. Patient 2
3.2.3. Patient 3
3.3. IgG N-glycosylation Profiles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schmidt, E.; Kasperkiewicz, M.; Joly, P. Pemphigus. Lancet 2019, 394, 882–894. [Google Scholar] [CrossRef]
- Anhalt, G.J.; Labib, R.S.; Voorhees, J.J.; Beals, T.F.; Diaz, L.A. Induction of pemphigus in neonatal mice by passive transfer of IgG from patients with the disease. N. Engl. J. Med. 1982, 306, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Roscoe, J.T.; Diaz, L.; Sampaio, S.A.; Castro, R.M.; Labib, R.S.; Takahashi, Y.; Patel, H.; Anhalt, G.J. Brazilian pemphigus foliaceus autoantibodies are pathogenic to BALB/c mice by passive transfer. J. Investig. Dermatol. 1985, 85, 538–541. [Google Scholar] [CrossRef] [Green Version]
- Drerup, K.; Folster-Holst, R. Neonatal pemphigus vulgaris. J. Dtsch. Dermatol. Ges. 2020, 18, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Kardos, M.; Levine, D.; Gurcan, H.M.; Ahmed, R.A. Pemphigus vulgaris in pregnancy: Analysis of current data on the management and outcomes. Obstet. Gynecol. Surv. 2009, 64, 739–749. [Google Scholar] [CrossRef]
- Hammers, C.M.; Stanley, J.R. Mechanisms of Disease: Pemphigus and Bullous Pemphigoid. Annu. Rev. Pathol. 2016, 11, 175–197. [Google Scholar] [CrossRef] [Green Version]
- Koneczny, I. A New Classification System for IgG4 Autoantibodies. Front. Immunol. 2018, 9, 97. [Google Scholar] [CrossRef]
- Schroeder, H.W., Jr.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [Green Version]
- Funakoshi, T.; Lunardon, L.; Ellebrecht, C.T.; Nagler, A.R.; O’Leary, C.E.; Payne, A.S. Enrichment of total serum IgG4 in patients with pemphigus. Br. J. Dermatol. 2012, 167, 1245–1253. [Google Scholar] [CrossRef]
- Kricheli, D.; David, M.; Frusic-Zlotkin, M.; Goldsmith, D.; Rabinov, M.; Sulkes, J.; Milner, Y. The distribution of pemphigus vulgaris-IgG subclasses and their reactivity with desmoglein 3 and 1 in pemphigus patients and their first-degree relatives. Br. J. Dermatol. 2000, 143, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Futei, Y.; Amagai, M.; Ishii, K.; Kuroda-Kinoshita, K.; Ohya, K.; Nishikawa, T. Predominant IgG4 subclass in autoantibodies of pemphigus vulgaris and foliaceus. J. Dermatol. Sci. 2001, 26, 55–61. [Google Scholar] [CrossRef]
- Torzecka, J.D.; Wozniak, K.; Kowalewski, C.; Waszczykowska, E.; Sysa-Jedrzejowska, A.; Pas, H.H.; Narbutt, J. Circulating pemphigus autoantibodies in healthy relatives of pemphigus patients: Coincidental phenomenon with a risk of disease development? Arch. Dermatol. Res. 2007, 299, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Haddad, G.; Lorenzen, J.M.; Ma, H.; de Haan, N.; Seeger, H.; Zaghrini, C.; Brandt, S.; Kolling, M.; Wegmann, U.; Kiss, B.; et al. Altered glycosylation of IgG4 promotes lectin complement pathway activation in anti-PLA2R1-associated membranous nephropathy. J. Clin. Investig. 2021, 131, e140453. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, R.; Wormald, M.R.; Rudd, P.M.; Fischer, P.B.; Dwek, R.A.; Sim, R.B. Glycosylation changes of IgG associated with rheumatoid arthritis can activate complement via the mannose-binding protein. Nat. Med. 1995, 1, 237–243. [Google Scholar] [CrossRef]
- Liu, H.; Bulseco, G.G.; Sun, J. Effect of posttranslational modifications on the thermal stability of a recombinant monoclonal antibody. Immunol. Lett. 2006, 106, 144–153. [Google Scholar] [CrossRef]
- Arnold, J.N.; Wormald, M.R.; Sim, R.B.; Rudd, P.M.; Dwek, R.A. The impact of glycosylation on the biological function and structure of human immunoglobulins. Annu. Rev. Immunol. 2007, 25, 21–50. [Google Scholar] [CrossRef]
- Shields, R.L.; Lai, J.; Keck, R.; O’Connell, L.Y.; Hong, K.; Meng, Y.G.; Weikert, S.H.; Presta, L.G. Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human Fcgamma RIII and antibody-dependent cellular toxicity. J. Biol. Chem. 2002, 277, 26733–26740. [Google Scholar] [CrossRef] [Green Version]
- van de Bovenkamp, F.S.; Hafkenscheid, L.; Rispens, T.; Rombouts, Y. The Emerging Importance of IgG Fab Glycosylation in Immunity. J. Immunol. 2016, 196, 1435–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeling, M.; Bruckner, C.; Nimmerjahn, F. Differential antibody glycosylation in autoimmunity: Sweet biomarker or modulator of disease activity? Nat. Rev. Rheumatol. 2017, 13, 621–630. [Google Scholar] [CrossRef]
- Kemna, M.J.; Plomp, R.; van Paassen, P.; Koeleman, C.A.M.; Jansen, B.C.; Damoiseaux, J.; Cohen Tervaert, J.W.; Wuhrer, M. Galactosylation and Sialylation Levels of IgG Predict Relapse in Patients With PR3-ANCA Associated Vasculitis. EBioMedicine 2017, 17, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, E.; Shinzaki, S.; Fujii, H.; Iijima, H.; Kamada, Y.; Takehara, T. Role of aberrant IgG glycosylation in the pathogenesis of inflammatory bowel disease. Proteom. Clin. Appl. 2016, 10, 384–390. [Google Scholar] [CrossRef]
- Vuckovic, F.; Kristic, J.; Gudelj, I.; Teruel, M.; Keser, T.; Pezer, M.; Pucic-Bakovic, M.; Stambuk, J.; Trbojevic-Akmacic, I.; Barrios, C.; et al. Association of systemic lupus erythematosus with decreased immunosuppressive potential of the IgG glycome. Arthritis Rheumatol. 2015, 67, 2978–2989. [Google Scholar] [CrossRef]
- Almugairen, N.; Hospital, V.; Bedane, C.; Duvert-Lehembre, S.; Picard, D.; Tronquoy, A.F.; Houivet, E.; D’Incan, M.; Joly, P. Assessment of the rate of long-term complete remission off therapy in patients with pemphigus treated with different regimens including medium- and high-dose corticosteroids. J. Am. Acad. Dermatol. 2013, 69, 583–588. [Google Scholar] [CrossRef]
- Joly, P.; Maho-Vaillant, M.; Prost-Squarcioni, C.; Hebert, V.; Houivet, E.; Calbo, S.; Caillot, F.; Golinski, M.L.; Labeille, B.; Picard-Dahan, C.; et al. First-line rituximab combined with short-term prednisone versus prednisone alone for the treatment of pemphigus (Ritux 3): A prospective, multicentre, parallel-group, open-label randomised trial. Lancet 2017, 389, 2031–2040. [Google Scholar] [CrossRef]
- Maho-Vaillant, M.; Perals, C.; Golinski, M.L.; Hebert, V.; Caillot, F.; Mignard, C.; Riou, G.; Petit, M.; Viguier, M.; Hertl, M.; et al. Rituximab and Corticosteroid Effect on Desmoglein-Specific B-Cells and Desmoglein-Specific T-Follicular-Helper-Cells in Pemphigus. J. Investig. Dermatol. 2021, 16, 31. [Google Scholar] [CrossRef]
- Ishii, K.; Harada, R.; Matsuo, I.; Shirakata, Y.; Hashimoto, K.; Amagai, M. In vitro keratinocyte dissociation assay for evaluation of the pathogenicity of anti-desmoglein 3 IgG autoantibodies in pemphigus vulgaris. J. Investig Dermatol. 2005, 124, 939–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsunoda, K.; Ota, T.; Aoki, M.; Yamada, T.; Nagai, T.; Nakagawa, T.; Koyasu, S.; Nishikawa, T.; Amagai, M. Induction of pemphigus phenotype by a mouse monoclonal antibody against the amino-terminal adhesive interface of desmoglein 3. J. Immunol. 2003, 170, 2170–2178. [Google Scholar] [CrossRef] [Green Version]
- Vollner, F.; Ali, J.; Kurrle, N.; Exner, Y.; Eming, R.; Hertl, M.; Banning, A.; Tikkanen, R. Loss of flotillin expression results in weakened desmosomal adhesion and Pemphigus vulgaris-like localisation of desmoglein-3 in human keratinocytes. Sci. Rep. 2016, 6, 28820. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.C.; Bardor, M.; Feng, H.; Tong, Y.W.; Song, Z.; Yap, M.G.; Yang, Y. IRES-mediated Tricistronic vectors for enhancing generation of high monoclonal antibody expressing CHO cell lines. J. Biotechnol. 2012, 157, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Vanier, G.; Lucas, P.L.; Loutelier-Bourhis, C.; Vanier, J.; Plasson, C.; Walet-Balieu, M.L.; Tchi-Song, P.C.; Remy-Jouet, I.; Richard, V.; Bernard, S.; et al. Heterologous expression of the N-acetylglucosaminyltransferase I dictates a reinvestigation of the N-glycosylation pathway in Chlamydomonas reinhardtii. Sci. Rep. 2017, 7, 10156. [Google Scholar] [CrossRef] [Green Version]
- Lucas, P.L.; Dumontier, R.; Loutelier-Bourhis, C.; Mareck, A.; Afonso, C.; Lerouge, P.; Mati-Baouche, N.; Bardor, M. User-friendly extraction and multistage tandem mass spectrometry based analysis of lipid-linked oligosaccharides in microalgae. Plant Methods 2018, 14, 107. [Google Scholar] [CrossRef]
- Neelamegham, S.; Aoki-Kinoshita, K.; Bolton, E.; Frank, M.; Lisacek, F.; Lutteke, T.; O’Boyle, N.; Packer, N.H.; Stanley, P.; Toukach, P.; et al. Updates to the Symbol Nomenclature for Glycans guidelines. Glycobiology 2019, 29, 620–624. [Google Scholar] [CrossRef]
- Alter, G.; Ottenhoff, T.H.M.; Joosten, S.A. Antibody glycosylation in inflammation, disease and vaccination. Semin. Immunol. 2018, 39, 102–110. [Google Scholar] [CrossRef]
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nat. Rev. Drug Discov. 2009, 8, 226–234. [Google Scholar] [CrossRef]
- Mahan, A.E.; Jennewein, M.F.; Suscovich, T.; Dionne, K.; Tedesco, J.; Chung, A.W.; Streeck, H.; Pau, M.; Schuitemaker, H.; Francis, D.; et al. Antigen-Specific Antibody Glycosylation Is Regulated via Vaccination. PLoS Pathog. 2016, 12, e1005456. [Google Scholar] [CrossRef] [Green Version]
- Selman, M.H.; de Jong, S.E.; Soonawala, D.; Kroon, F.P.; Adegnika, A.A.; Deelder, A.M.; Hokke, C.H.; Yazdanbakhsh, M.; Wuhrer, M. Changes in antigen-specific IgG1 Fc N-glycosylation upon influenza and tetanus vaccination. Mol. Cell Proteom. 2012, 11, M111.014563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifle, R.; Rothe, T.; Ipseiz, N.; Scherer, H.U.; Culemann, S.; Harre, U.; Ackermann, J.A.; Seefried, M.; Kleyer, A.; Uderhardt, S.; et al. Regulation of autoantibody activity by the IL-23-TH17 axis determines the onset of autoimmune disease. Nat. Immunol. 2017, 18, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Gudelj, I.; Salo, P.P.; Trbojevic-Akmacic, I.; Albers, M.; Primorac, D.; Perola, M.; Lauc, G. Low galactosylation of IgG associates with higher risk for future diagnosis of rheumatoid arthritis during 10years of follow-up. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2034–2039. [Google Scholar] [CrossRef] [PubMed]
- Hennig, R.; Cajic, S.; Borowiak, M.; Hoffmann, M.; Kottler, R.; Reichl, U.; Rapp, E. Towards personalized diagnostics via longitudinal study of the human plasma N-glycome. Biochim. Biophys. Acta 2016, 1860, 1728–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, Y.; Nimmerjahn, F.; Ravetch, J.V. Anti-inflammatory activity of immunoglobulin G resulting from Fc sialylation. Science 2006, 313, 670–673. [Google Scholar] [CrossRef] [Green Version]
- Parekh, R.B.; Dwek, R.A.; Sutton, B.J.; Fernandes, D.L.; Leung, A.; Stanworth, D.; Rademacher, T.W.; Mizuochi, T.; Taniguchi, T.; Matsuta, K.; et al. Association of rheumatoid arthritis and primary osteoarthritis with changes in the glycosylation pattern of total serum IgG. Nature 1985, 316, 452–457. [Google Scholar] [CrossRef]
- Sonneveld, M.E.; de Haas, M.; Koeleman, C.; de Haan, N.; Zeerleder, S.S.; Ligthart, P.C.; Wuhrer, M.; van der Schoot, C.E.; Vidarsson, G. Patients with IgG1-anti-red blood cell autoantibodies show aberrant Fc-glycosylation. Sci. Rep. 2017, 7, 8187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coloma, M.J.; Trinh, R.K.; Martinez, A.R.; Morrison, S.L. Position effects of variable region carbohydrate on the affinity and in vivo behavior of an anti-(1→6) dextran antibody. J. Immunol. 1999, 162, 2162–2170. [Google Scholar]
- Leibiger, H.; Wustner, D.; Stigler, R.D.; Marx, U. Variable domain-linked oligosaccharides of a human monoclonal IgG: Structure and influence on antigen binding. Biochem. J. 1999, 338, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Wallick, S.C.; Kabat, E.A.; Morrison, S.L. Glycosylation of a VH residue of a monoclonal antibody against alpha (1----6) dextran increases its affinity for antigen. J. Exp. Med. 1988, 168, 1099–1109. [Google Scholar] [CrossRef]
- Sabouri, Z.; Schofield, P.; Horikawa, K.; Spierings, E.; Kipling, D.; Randall, K.L.; Langley, D.; Roome, B.; Vazquez-Lombardi, R.; Rouet, R.; et al. Redemption of autoantibodies on anergic B cells by variable-region glycosylation and mutation away from self-reactivity. Proc. Natl. Acad. Sci. USA 2014, 111, E2567–E2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondt, A.; Wuhrer, M.; Kuijper, T.M.; Hazes, J.M.; Dolhain, R.J. Fab glycosylation of immunoglobulin G does not associate with improvement of rheumatoid arthritis during pregnancy. Arthritis Res. Ther. 2016, 18, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients | 3 |
|---|---|
| Age, mean | 69.4 |
| Sex | |
| Female | 2 |
| Male | 1 |
| BMI, mean | 24.46 |
| Type of pemphigus | |
| Vulgaris | 3 |
| Foliaceus | 0 |
| Initial Presentation | |
| Mucosal | 0 |
| Cutaneous | 0 |
| Mucocutaneous | 3 |
| PDAI score, mean | 45.6 |
| Treatment | corticosteroids |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petit, M.; Walet-Balieu, M.-L.; Schapman, D.; Golinski, M.-L.; Burel, C.; Barray, M.; Drouot, L.; Maho-Vaillant, M.; Hébert, V.; Boyer, O.; et al. Longitudinal Pathogenic Properties and N-Glycosylation Profile of Antibodies from Patients with Pemphigus after Corticosteroid Treatment. Biomedicines 2021, 9, 1411. https://doi.org/10.3390/biomedicines9101411
Petit M, Walet-Balieu M-L, Schapman D, Golinski M-L, Burel C, Barray M, Drouot L, Maho-Vaillant M, Hébert V, Boyer O, et al. Longitudinal Pathogenic Properties and N-Glycosylation Profile of Antibodies from Patients with Pemphigus after Corticosteroid Treatment. Biomedicines. 2021; 9(10):1411. https://doi.org/10.3390/biomedicines9101411
Chicago/Turabian StylePetit, Marie, Marie-Laure Walet-Balieu, Damien Schapman, Marie-Laure Golinski, Carole Burel, Marion Barray, Laurent Drouot, Maud Maho-Vaillant, Vivien Hébert, Olivier Boyer, and et al. 2021. "Longitudinal Pathogenic Properties and N-Glycosylation Profile of Antibodies from Patients with Pemphigus after Corticosteroid Treatment" Biomedicines 9, no. 10: 1411. https://doi.org/10.3390/biomedicines9101411
APA StylePetit, M., Walet-Balieu, M.-L., Schapman, D., Golinski, M.-L., Burel, C., Barray, M., Drouot, L., Maho-Vaillant, M., Hébert, V., Boyer, O., Bardor, M., Joly, P., & Calbo, S. (2021). Longitudinal Pathogenic Properties and N-Glycosylation Profile of Antibodies from Patients with Pemphigus after Corticosteroid Treatment. Biomedicines, 9(10), 1411. https://doi.org/10.3390/biomedicines9101411