Clinicopathological and Functional Evaluation Reveal NBS1 as a Predictor of Platinum Resistance in Epithelial Ovarian Cancers

 ,
, 

Abstract
1. Introduction
2. Experimental Section
2.1. Clinical Study
2.1.1. NBS1 Protein Expression in Ovarian Cancers
2.1.2. Tissue Microarray (TMA) and Immunohistochemistry (IHC)
2.1.3. Evaluation of Immune Staining
2.1.4. Statistical Analysis
2.1.5. NBS1 Transcript in Ovarian Cancers
2.2. Pre-Clinical Study
2.2.1. Cell Lines and Tissue Culture
2.2.2. Nuclear/Cytoplasmic Extracts and Western Blot Analysis
2.2.3. Protein Quantification
2.2.4. Transient Knockdowns of NBS1
2.2.5. Clonogenic Assays
2.2.6. DSB Accumulation, Cell Cycle and Apoptosis Analysis by Flow Cytometry
2.2.7. Statistical Analysis
2.2.8. Next Generation Sequencing and Bioinformatics
3. Results
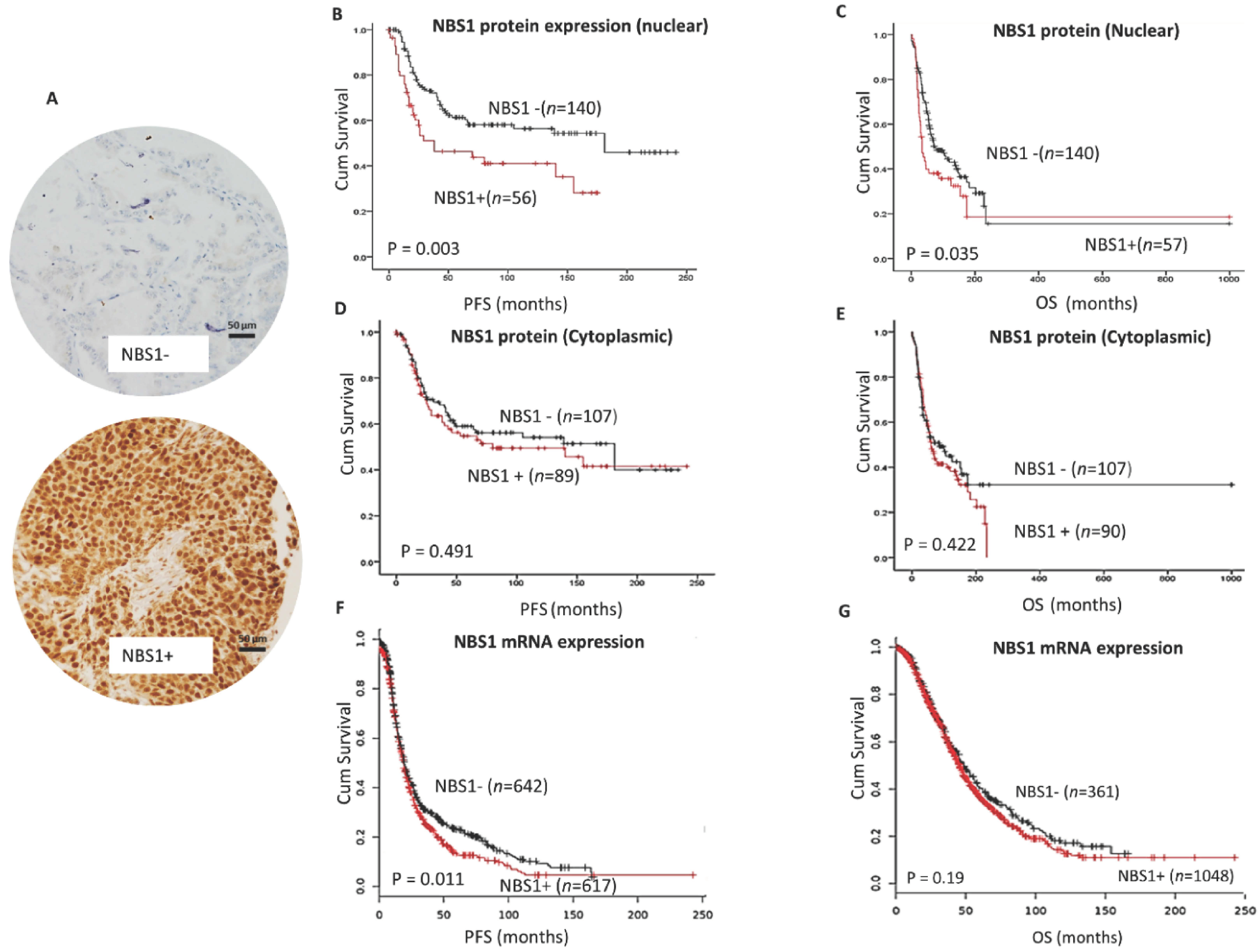
3.1. NBS1 Overexpression and Platinum Resistant Aggressive Ovarian Cancers
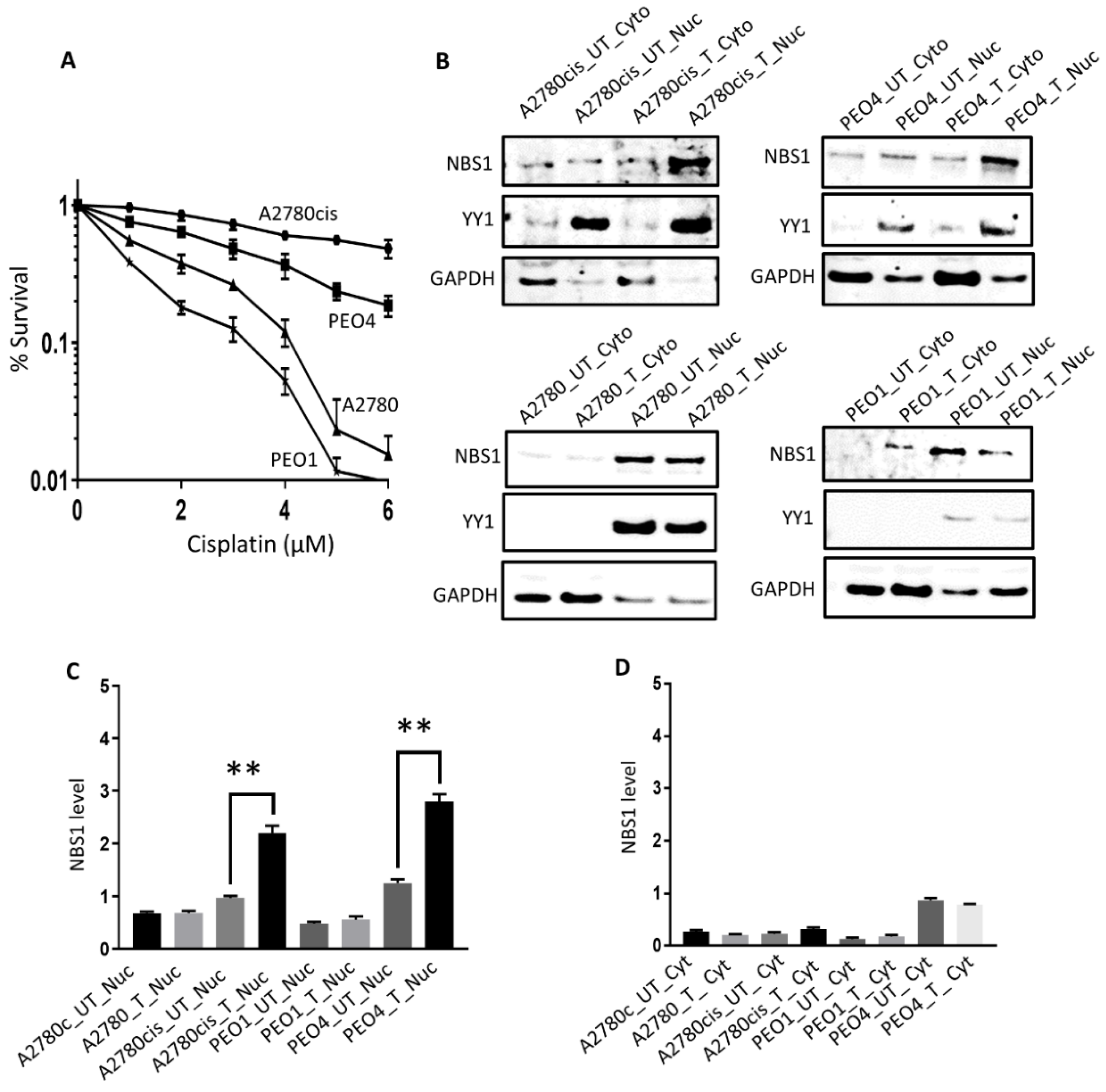
3.2. Sub-Cellular Localization of NBS1 in Ovarian Cancer Cells Following Cisplatin Therapy
3.3. NBS1 Variant Profiling in A2780, A2780cis, PEO1, and PEO4 Cells
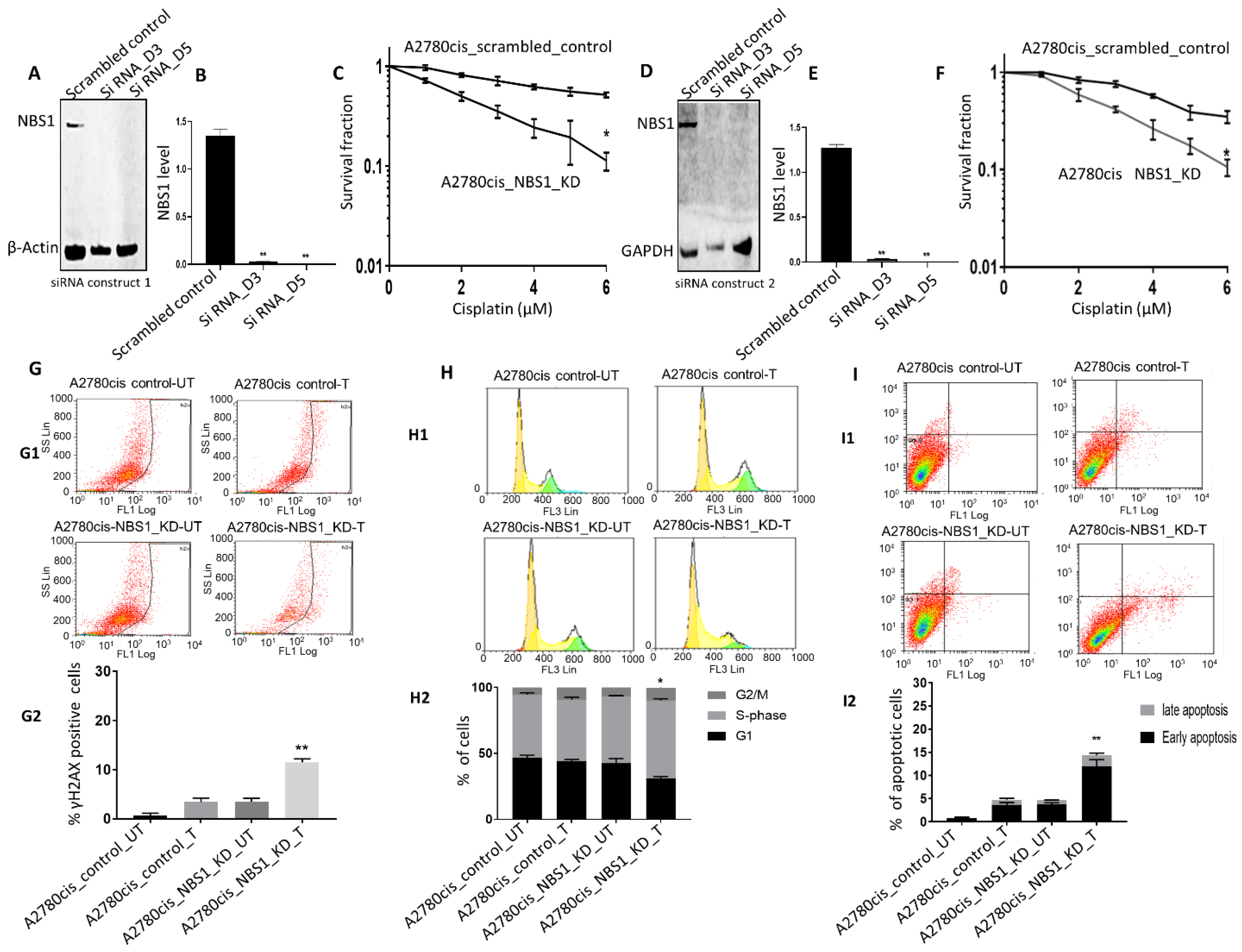
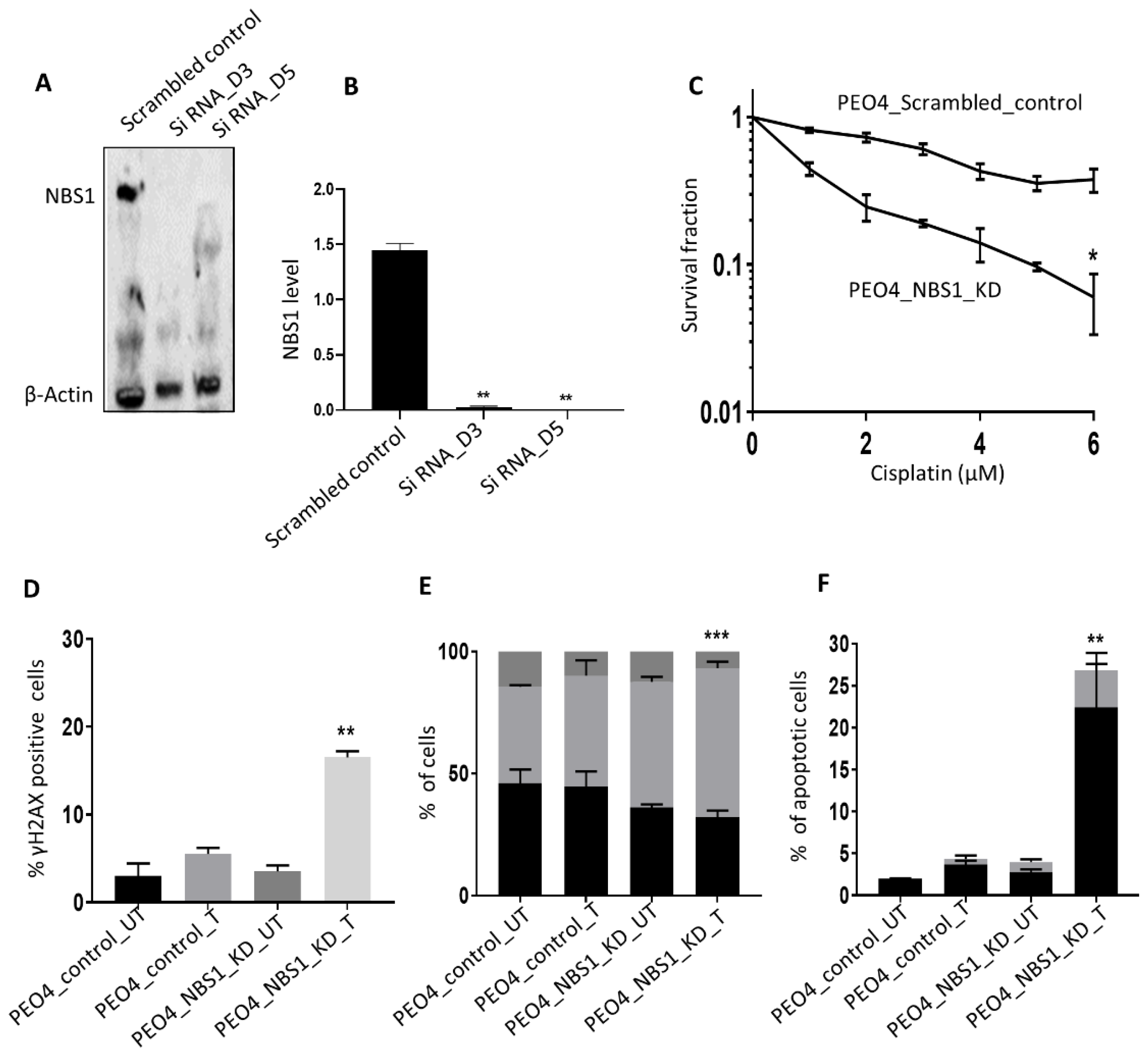
3.4. NBS1 Depletion and Platinum Sensitivity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raja, F.A.; Chopra, N.; Ledermann, J.A. Optimal first-line treatment in ovarian cancer. Ann. Oncol. 2012, 23 (Suppl. 10), x118–x127. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, S.; Coleman, R.L.; Rauh-Hain, J.A. Management and Treatment of Recurrent Epithelial Ovarian Cancer. Hematol. Oncol. Clin. N. Am. 2018, 32, 965–982. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.Y.; Lu, Y. New developments in molecular targeted therapy of ovarian cancer. Discov. Med. 2018, 26, 219–229. [Google Scholar] [PubMed]
- Orr, B.; Edwards, R.P. Diagnosis and Treatment of Ovarian Cancer. Hematol. Oncol. Clin. N. Am. 2018, 32, 943–964. [Google Scholar] [CrossRef] [PubMed]
- van Zyl, B.; Tang, D.; Bowden, N.A. Biomarkers of platinum resistance in ovarian cancer: What can we use to improve treatment. Endocr. Relat. Cancer 2018, 25, R303–R318. [Google Scholar] [CrossRef]
- Pujade-Lauraine, E.; Banerjee, S.; Pignata, S. Management of Platinum-Resistant, Relapsed Epithelial Ovarian Cancer and New Drug Perspectives. J. Clin. Oncol. 2019, 37, 2437–2448. [Google Scholar] [CrossRef]
- Binju, M.; Padilla, M.A.; Singomat, T.; Kaur, P.; Suryo Rahmanto, Y.; Cohen, P.A.; Yu, Y. Mechanisms underlying acquired platinum resistance in high grade serous ovarian cancer—A mini review. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 371–378. [Google Scholar] [CrossRef]
- Fuertes, M.A.; Castilla, J.; Alonso, C.; Perez, J.M. Cisplatin biochemical mechanism of action: From cytotoxicity to induction of cell death through interconnections between apoptotic and necrotic pathways. Curr. Med. Chem. 2003, 10, 257–266. [Google Scholar] [CrossRef]
- Damia, G.; Broggini, M. Platinum Resistance in Ovarian Cancer: Role of DNA Repair. Cancers 2019, 11, 119. [Google Scholar] [CrossRef]
- Pothuri, B.; O’Cearbhaill, R.; Eskander, R.; Armstrong, D. Frontline PARP inhibitor maintenance therapy in ovarian cancer: A Society of Gynecologic Oncology practice statement. Gynecol. Oncol. 2020, 159, 8–12. [Google Scholar] [CrossRef]
- D’Andrea, A.D. Mechanisms of PARP inhibitor sensitivity and resistance. DNA Repair 2018, 71, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Symington, L.S. Role of the Mre11 Complex in Preserving Genome Integrity. Genes 2018, 9, 589. [Google Scholar] [CrossRef] [PubMed]
- Paull, T.T. 20 Years of Mre11 Biology: No End in Sight. Mol. Cell 2018, 71, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.; Tainer, J.A. The MRE11-RAD50-NBS1 Complex Conducts the Orchestration of Damage Signaling and Outcomes to Stress in DNA Replication and Repair. Annu. Rev. Biochem. 2018, 87, 263–294. [Google Scholar] [CrossRef] [PubMed]
- Tisi, R.; Vertemara, J.; Zampella, G.; Longhese, M.P. Functional and structural insights into the MRX/MRN complex, a key player in recognition and repair of DNA double-strand breaks. Comput. Struct. Biotechnol. J. 2020, 18, 1137–1152. [Google Scholar] [CrossRef]
- Bian, L.; Meng, Y.; Zhang, M.; Li, D. MRE11-RAD50-NBS1 complex alterations and DNA damage response: Implications for cancer treatment. Mol. Cancer 2019, 18, 169. [Google Scholar] [CrossRef]
- Komatsu, K. NBS1 and multiple regulations of DNA damage response. J. Radiat. Res. 2016, 57 (Suppl. 1), i11–i17. [Google Scholar] [CrossRef]
- Antoccia, A.; Kobayashi, J.; Tauchi, H.; Matsuura, S.; Komatsu, K. Nijmegen breakage syndrome and functions of the responsible protein, NBS1. Genome Dyn. 2006, 1, 191–205. [Google Scholar] [CrossRef]
- Difilippantonio, S.; Nussenzweig, A. The NBS1-ATM connection revisited. Cell Cycle 2007, 6, 2366–2370. [Google Scholar] [CrossRef]
- Kobayashi, J.; Antoccia, A.; Tauchi, H.; Matsuura, S.; Komatsu, K. NBS1 and its functional role in the DNA damage response. DNA Repair 2004, 3, 855–861. [Google Scholar] [CrossRef]
- Gao, P.; Ma, N.; Li, M.; Tian, Q.B.; Liu, D.W. Functional variants in NBS1 and cancer risk: Evidence from a meta-analysis of 60 publications with 111 individual studies. Mutagenesis 2013, 28, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Lei, R.; Ding, S.W.; Zhu, S. Skewer: A fast and accurate adapter trimmer for next-generation sequencing paired-end reads. BMC Bioinform. 2014, 15, 182. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Mose, L.E.; Wilkerson, M.D.; Hayes, D.N.; Perou, C.M.; Parker, J.S. ABRA: Improved coding indel detection via assembly-based realignment. Bioinformatics 2014, 30, 2813–2815. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef]
- Negorev, D.G.; Vladimirova, O.V.; Kossenkov, A.V.; Nikonova, E.V.; Demarest, R.M.; Capobianco, A.J.; Showe, M.K.; Rauscher, F.J., 3rd; Showe, L.C.; Maul, G.G. Sp100 as a potent tumor suppressor: Accelerated senescence and rapid malignant transformation of human fibroblasts through modulation of an embryonic stem cell program. Cancer Res. 2010, 70, 9991–10001. [Google Scholar] [CrossRef]
- Lee, Y.K.; Park, N.H.; Lee, H. Clinicopathological values of NBS1 and DNA damage response genes in epithelial ovarian cancers. Exp. Mol. Med. 2015, 47, e195. [Google Scholar] [CrossRef]
- Brandt, S.; Samartzis, E.P.; Zimmermann, A.K.; Fink, D.; Moch, H.; Noske, A.; Dedes, K.J. Lack of MRE11-RAD50-NBS1 (MRN) complex detection occurs frequently in low-grade epithelial ovarian cancer. BMC Cancer 2017, 17, 44. [Google Scholar] [CrossRef]
- Hsu, D.S.; Chang, S.Y.; Liu, C.J.; Tzeng, C.H.; Wu, K.J.; Kao, J.Y.; Yang, M.H. Identification of increased NBS1 expression as a prognostic marker of squamous cell carcinoma of the oral cavity. Cancer Sci. 2010, 101, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.H.; Chiang, W.C.; Chou, T.Y.; Chang, S.Y.; Chen, P.M.; Teng, S.C.; Wu, K.J. Increased NBS1 expression is a marker of aggressive head and neck cancer and overexpression of NBS1 contributes to transformation. Clin. Cancer Res. 2006, 12, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, J.P.; Harbour, J.W. NBS1 expression as a prognostic marker in uveal melanoma. Clin. Cancer Res. 2005, 11, 1849–1853. [Google Scholar] [CrossRef] [PubMed]
- Altan, B.; Yokobori, T.; Ide, M.; Bai, T.; Yanoma, T.; Kimura, A.; Kogure, N.; Suzuki, M.; Bao, P.; Mochiki, E.; et al. High Expression of MRE11-RAD50-NBS1 Is Associated with Poor Prognosis and Chemoresistance in Gastric Cancer. Anticancer Res. 2016, 36, 5237–5247. [Google Scholar] [CrossRef] [PubMed]
- Situ, Y.; Chung, L.; Lee, C.S.; Ho, V. MRN (MRE11-RAD50-NBS1) Complex in Human Cancer and Prognostic Implications in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 816. [Google Scholar] [CrossRef]
- Araki, K.; Yamashita, T.; Reddy, N.; Wang, H.; Abuzeid, W.M.; Khan, K.; O’Malley, B.W., Jr.; Li, D. Molecular disruption of NBS1 with targeted gene delivery enhances chemosensitisation in head and neck cancer. Br. J. Cancer 2010, 103, 1822–1830. [Google Scholar] [CrossRef]
- Tran, H.M.; Shi, G.; Li, G.; Carney, J.P.; O’Malley, B.; Li, D. Mutant Nbs1 enhances cisplatin-induced DNA damage and cytotoxicity in head and neck cancer. Otolaryngol. Head Neck Surg. 2004, 131, 477–484. [Google Scholar] [CrossRef]
- Hoppe, M.M.; Sundar, R.; Tan, D.S.P.; Jeyasekharan, A.D. Biomarkers for Homologous Recombination Deficiency in Cancer. J. Natl. Cancer Inst. 2018, 110, 704–713. [Google Scholar] [CrossRef]
- Barroso, S.I.; Aguilera, A. Detection of DNA Double-Strand Breaks by gamma-H2AX Immunodetection. In Homologous Recombination; Humana: New York, NY, USA, 2020; Volume 2153, pp. 1–8. [Google Scholar] [CrossRef]
- McCabe, N.; Turner, N.C.; Lord, C.J.; Kluzek, K.; Bialkowska, A.; Swift, S.; Giavara, S.; O’Connor, M.J.; Tutt, A.N.; Zdzienicka, M.Z.; et al. Deficiency in the repair of DNA damage by homologous recombination and sensitivity to poly(ADP-ribose) polymerase inhibition. Cancer Res. 2006, 66, 8109–8115. [Google Scholar] [CrossRef]
- Daemen, A.; Wolf, D.M.; Korkola, J.E.; Griffith, O.L.; Frankum, J.R.; Brough, R.; Jakkula, L.R.; Wang, N.J.; Natrajan, R.; Reis-Filho, J.S.; et al. Cross-platform pathway-based analysis identifies markers of response to the PARP inhibitor olaparib. Breast Cancer Res. Treat. 2012, 135, 505–517. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NBS Protein Expression (Nuclear) | p-Value | ||
| Low | High | 0.0001 | |
| Sensitive | 131 (74.9%) | 44 (25.1%) | |
| Resistant * | 5 (31.3%) | 11 (68.8%) | |
| NBS Protein Expression (Cytoplasmic) | p-Value | ||
| Low | High | 0.676 | |
| Sensitive | 97 (55.4%) | 78 (44.6%) | |
| Resistant * | 8 (50%) | 8 (50%) | |
| Progression Free Survival (PFS) | ||||
| Sig. | Exp(B) | 95.0% CI for Exp(B) | ||
| Lower | Upper | |||
| NBS1 (Nuclear) | 0.017 | 1.803 | 1.111 | 2.925 |
| NBS1 (cytoplasmic) | 0.476 | 1.187 | 0.741 | 1.903 |
| Tumor Stage | 0.000 | 2.296 | 1.762 | 2.992 |
| Overall Survival (OS) | ||||
| Sig. | Exp(B) | 95.0% CI for Exp(B) | ||
| Lower | Upper | |||
| NBS1 (nuclear) | 0.009 | 1.746 | 1.147 | 2.658 |
| NBS1 (cytoplasmic) | 0.612 | 0.902 | 0.605 | 1.344 |
| Tumor Stage | 0.000 | 2.160 | 1.727 | 2.700 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alblihy, A.; Alabdullah, M.L.; Ali, R.; Algethami, M.; Toss, M.S.; Mongan, N.P.; Rakha, E.A.; Madhusudan, S. Clinicopathological and Functional Evaluation Reveal NBS1 as a Predictor of Platinum Resistance in Epithelial Ovarian Cancers. Biomedicines 2021, 9, 56. https://doi.org/10.3390/biomedicines9010056
Alblihy A, Alabdullah ML, Ali R, Algethami M, Toss MS, Mongan NP, Rakha EA, Madhusudan S. Clinicopathological and Functional Evaluation Reveal NBS1 as a Predictor of Platinum Resistance in Epithelial Ovarian Cancers. Biomedicines. 2021; 9(1):56. https://doi.org/10.3390/biomedicines9010056
Chicago/Turabian StyleAlblihy, Adel, Muslim L. Alabdullah, Reem Ali, Mashael Algethami, Michael S. Toss, Nigel P. Mongan, Emad A. Rakha, and Srinivasan Madhusudan. 2021. "Clinicopathological and Functional Evaluation Reveal NBS1 as a Predictor of Platinum Resistance in Epithelial Ovarian Cancers" Biomedicines 9, no. 1: 56. https://doi.org/10.3390/biomedicines9010056
APA StyleAlblihy, A., Alabdullah, M. L., Ali, R., Algethami, M., Toss, M. S., Mongan, N. P., Rakha, E. A., & Madhusudan, S. (2021). Clinicopathological and Functional Evaluation Reveal NBS1 as a Predictor of Platinum Resistance in Epithelial Ovarian Cancers. Biomedicines, 9(1), 56. https://doi.org/10.3390/biomedicines9010056