Reference Gene Validation in the Brain Regions of Young Rats after Pentylenetetrazole-Induced Seizures

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
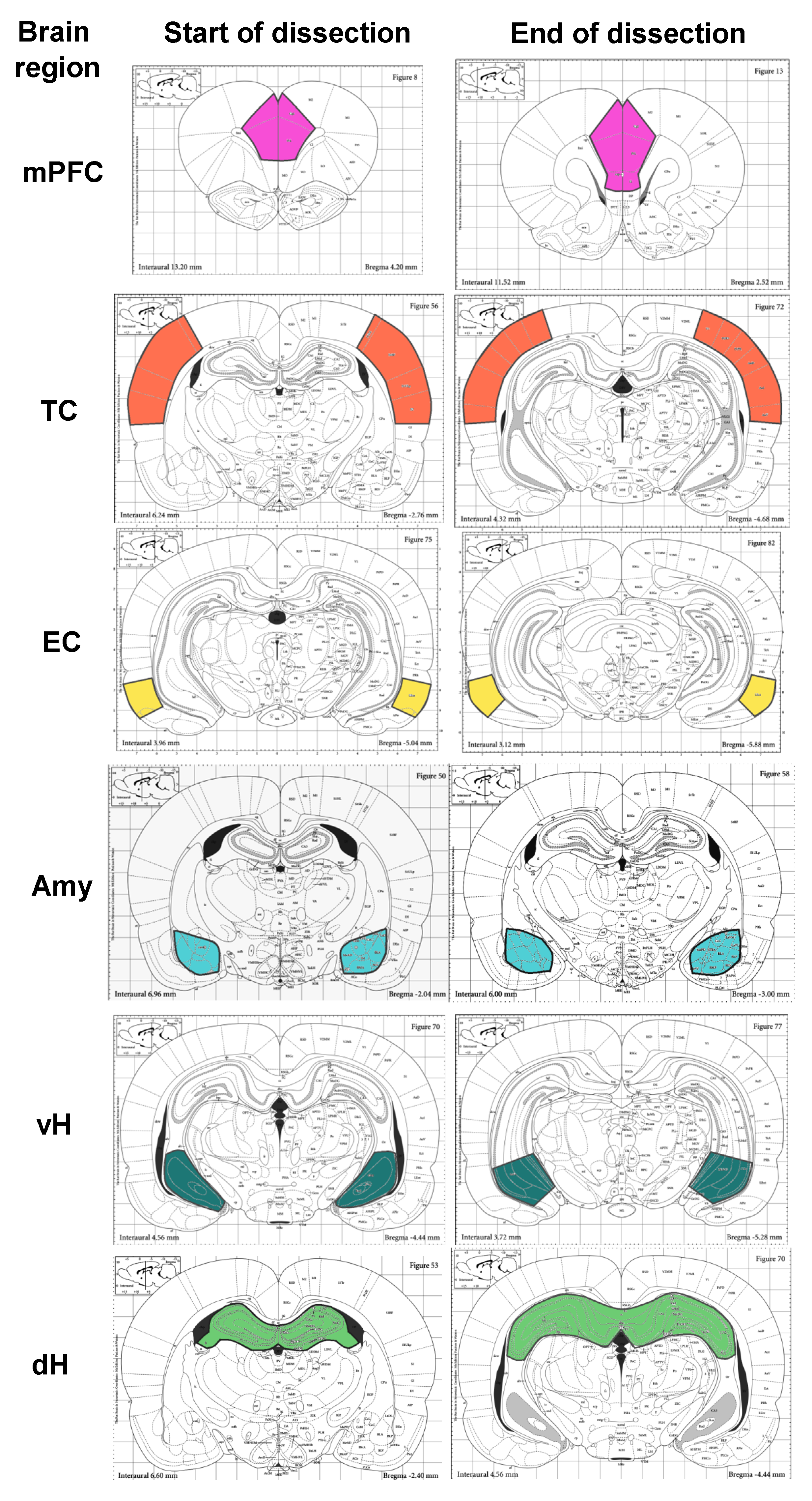
2.1. PTZ Model of Seizures
2.2. mRNA Extraction and cDNA Synthesis
2.3. qRT-PCR
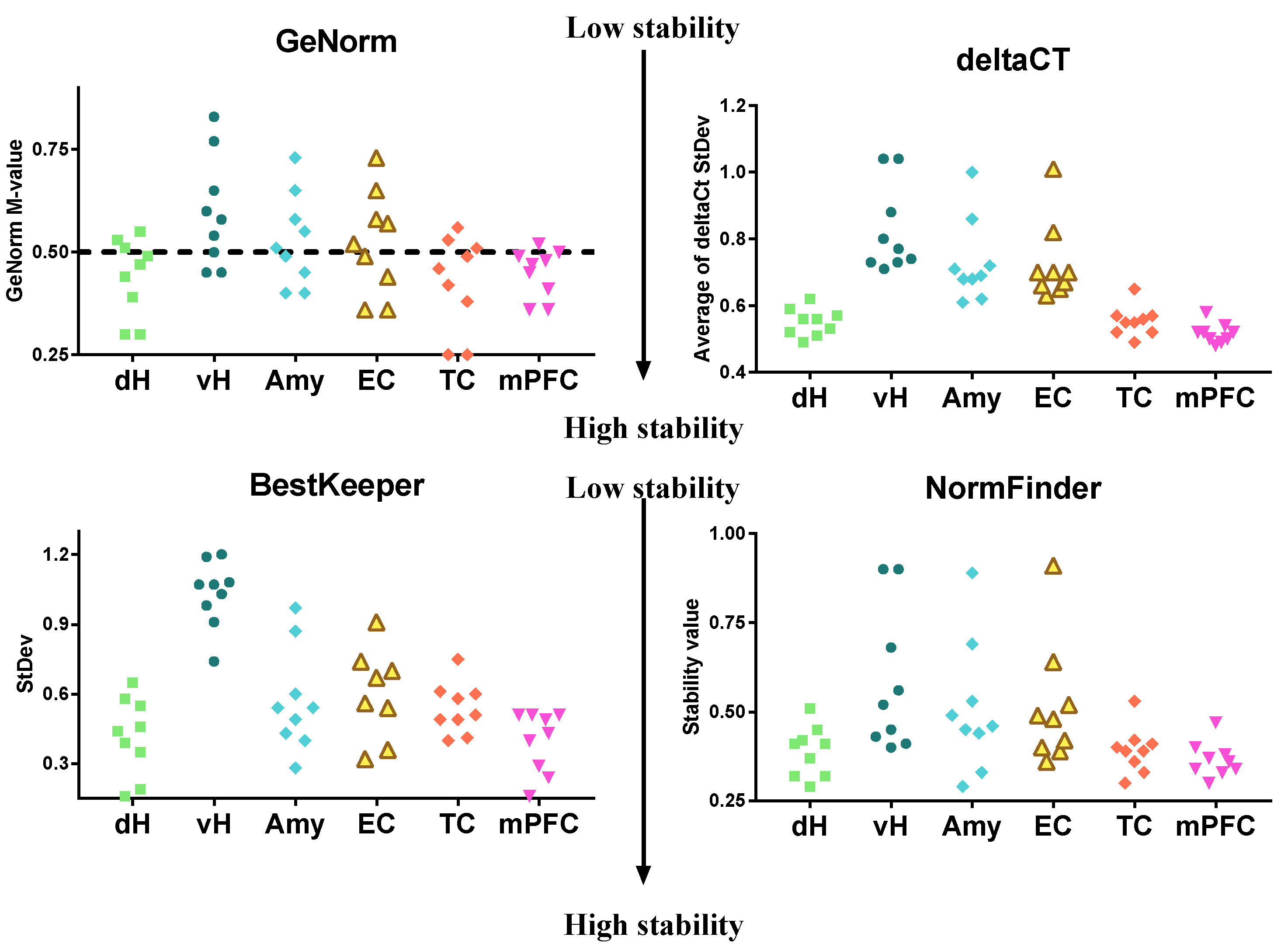
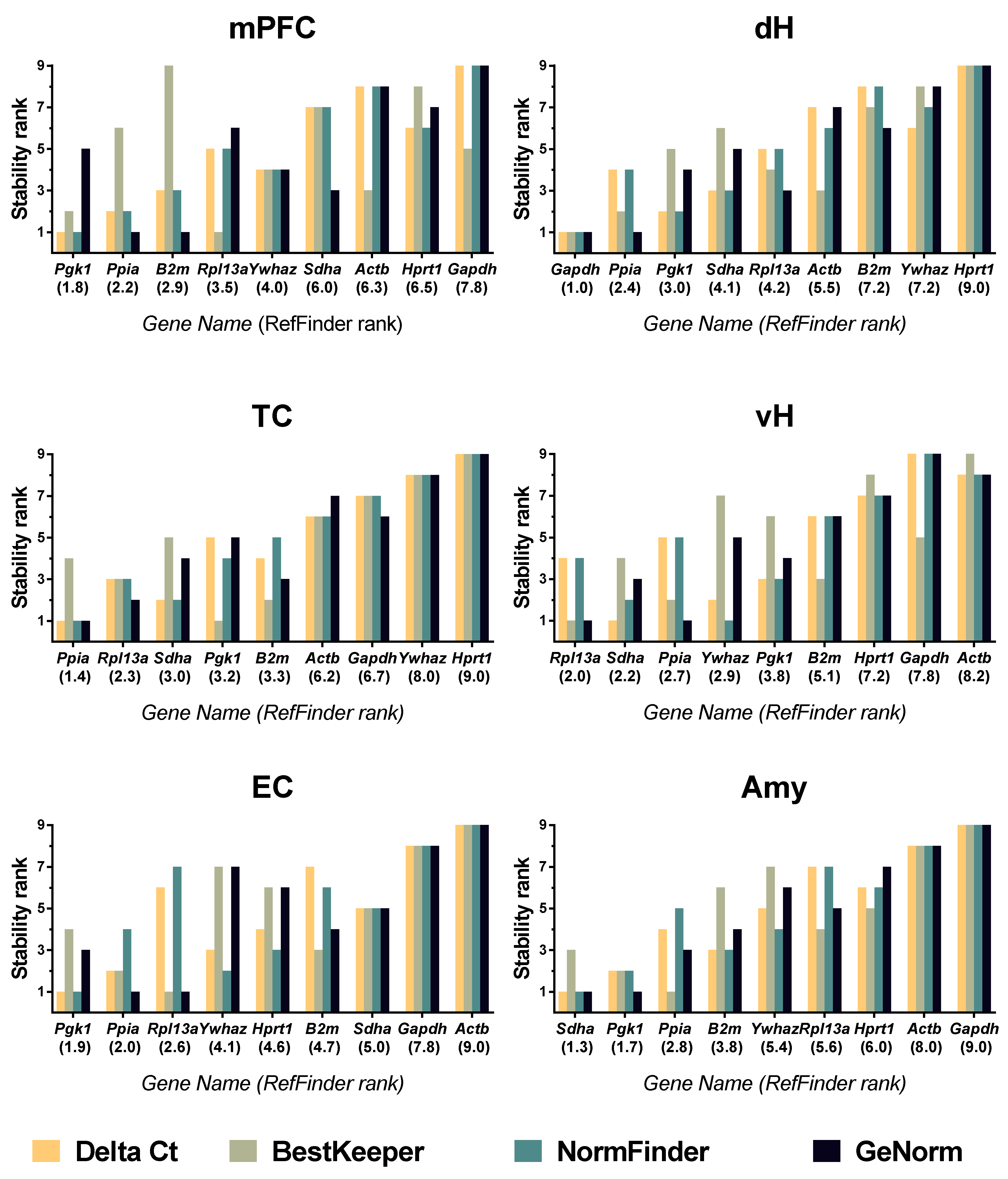
2.4. Data Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol RefSeq Accession Number | The Encoded Protein and Its Function | Primer 5′–3′ (Forward, Reverse, Probe) | Amplicon Length and Position on mRNA | Reference |
|---|---|---|---|---|
| Gapdh1 NM_017008 | Glyceraldehyde-3-phosphate dehydrogenase Glycolytic enzyme | TGCACCACCAACTGCTTAG GGATGCAGGGATGATGTTC ATCACGCCACAGCTTTCCAGAGGG * | 177 bp (523–699) | [49] |
| B2m1 NM_012512 | Beta-2-microglobulin Assembly and surface expression of MHC class I molecules | TGCCATTCAGAAAACTCCCC GAGGAAGTTGGGCTTCCCATT ATTCAAGTGTACTCTCGCCATCCACCG& | 73 bp (77–149) | [50] |
| Actb1 NM_031144 | Beta-actin Cytoskeletal protein | TGTCACCAACTGGGACGATA GGGGTGTTGAAGGTCTCAAA CGTGTGGCCCCTGAGGAGCAC # | 165 bp (303–467) | [51] (forward and reverse primers) [30] (hydrolysis probe) |
| Ppia2 NM_017101 | Peptidyl prolyl isomerase A, cyclophilin A Protein folding | AGGATTCATGTGCCAGGGTG CTCAGTCTTGGCAGTGCAGA CACGCCATAATGGCACTGGTGGCA & | 187 bp (216–402) | [33] |
| Rpl13a2 NM_173340 | Ribosomal protein L13A 60S ribosomal subunit protein | GGATCCCTCCACCCTATGACA CTGGTACTTCCACCCGACCTC CTGCCCTCAAGGTTGTGCGGCT # | 131 bp (334–464) | [2] (forward and reverse primers) [30] (hydrolysis probe) |
| Sdha3 NM_130428 | Succinate dehydrogenase complex, subunit a, flavoprotein Component of electron transport chain | AGACGTTTGACAGGGGAATG TCATCAATCCGCACCTTGTA ACCTGGTGGAGACGCTGGAGCT * | 160 bp (1666–1825) | [52] (forward and reverse primers) [30] (hydrolysis probe) |
| Ywhaz3 NM_013011 | Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, zeta polypeptide Cell signaling | GATGAAGCCATTGCTGAACTTG GTCTCCTTGGGTATCCGATGTC TGAAGAGTCGTACAAAGACAGCACGC & | 117 bp (650–766) | [14] (forward and reverse primers) [30] (hydrolysis probe) |
| Pgk13 NM_053291 | Phosphoglycerate kinase 1 Glycolytic enzyme | ATGCAAAGACTGGCCAAGCTAC AGCCACAGCCTCAGCATATTTC TGCTGGCTGGATGGGCTTGGA * | 104 bp (888–991) | [14] (forward and reverse primers) [30] (hydrolysis probe) |
| Hprt3 NM_012583 | Hypoxanthine phosphoribosyl transferase 1 Purine synthesis | TCCTCAGACCGCTTTTCCCGC TCATCATCACTAATCACGACGCTGG CCGACCGGTTCTGTCATGTCGACCCT # | 80 bp (28–107) | [53] (forward and reverse primers) [30] (hydrolysis probe) |
References
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Swijsen, A.; Nelissen, K.; Janssen, D.; Rigo, J.-M.; Hoogland, G. Validation of reference genes for quantitative real-time PCR studies in the dentate gyrus after experimental febrile seizures. BMC Res. Notes 2012, 5, 685. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhang, X.; Ren, J.; Wang, P.; Zhang, J.; Wei, Z.; Tian, Y. Validation of reference genes for quantitative real-time PCR in valproic acid rat models of autism. Mol. Biol. Rep. 2016, 43, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.L.; Kleinig, T.J.; Heuvel, C.V.D.; Vink, R. Reference genes for normalising gene expression data in collagenase-induced rat intracerebral haemorrhage. BMC Mol. Biol. 2010, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Zhao, L.; Feng, J.; You, G.; Sun, Q.; Li, P.; Han, D.; Zhou, H. Validation of reliable reference genes for real-time PCR in human umbilical vein endothelial cells on substrates with different stiffness. PLoS ONE 2013, 8, e67360. [Google Scholar] [CrossRef]
- Nair, A.R.; Smeets, K.; Keunen, E.; Lee, W.; Thévenod, F. Renal cells exposed to cadmium in vitro and in vivo: Normalizing gene expression data. J. Appl. Toxicol. 2014. [Google Scholar] [CrossRef]
- Crans, R.A.J.; Janssens, J.; Daelemans, S.; Wouters, E.; Raedt, R.; Van Dam, D.; De Deyn, P.P.; Van Craenenbroeck, K.; Stove, C.P. The validation of Short Interspersed Nuclear Elements (SINEs) as a RT-qPCR normalization strategy in a rodent model for temporal lobe epilepsy. PLoS ONE 2019, 14, e0210567. [Google Scholar] [CrossRef]
- Paiva, B.; Fraga, L.; Meirelles, S.; Nardi, N.B.; Camassola, M. Identi fi cation of suitable reference genes for quantitative gene expression analysis in rat adipose stromal cells induced to trilineage differentiation. Gene 2016. [Google Scholar] [CrossRef]
- Chapman, J.R.; Waldenström, J. With reference to reference genes: A systematic review of endogenous controls in gene expression studies. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langnaese, K.; John, R.; Schweizer, H.; Ebmeyer, U.; Keilhoff, G. Selection of reference genes for quantitative real-time PCR in a rat asphyxial cardiac arrest model. BMC Mol. Biol. 2008, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Sadangi, C.; Rosenow, F.; Norwood, B.A. Validation of reference genes for quantitative gene expression analysis in experimental epilepsy. J. Neurosci. Res. 2017, 95, 2357–2366. [Google Scholar] [CrossRef]
- Janssens, J.; Crans, R.A.J.; Van Craenenbroeck, K.; Vandesompele, J.; Stove, C.P.; Van Dam, D.; De Deyn, P.P. Evaluating the applicability of mouse SINEs as an alternative normalization approach for RT-qPCR in brain tissue of the APP23 model for Alzheimer’s disease. J. Neurosci. Methods 2019, 320, 128–137. [Google Scholar] [CrossRef]
- Augustyniak, J.; Lenart, J.; Lipka, G.; Stepien, P.P.; Buzanska, L. Reference Gene Validation via RT–qPCR for Human iPSC-Derived Neural Stem Cells and Neural Progenitors. Mol. Neurobiol. 2019, 56, 6820–6832. [Google Scholar] [CrossRef] [Green Version]
- Jacob, F.; Guertler, R.; Naim, S.; Nixdorf, S.; Fedier, A.; Hacker, N.F.; Heinzelmann-Schwarz, V. Careful selection of reference genes is required for reliable performance of RT-qPCR in human normal and cancer cell lines. PLoS ONE 2013, 8, e59180. [Google Scholar] [CrossRef] [Green Version]
- De Spiegelaere, W.; Dern-Wieloch, J.; Weigel, R.; Schumacher, V.; Schorle, H.; Nettersheim, D.; Bergmann, M.; Brehm, R.; Kliesch, S.; Vandekerckhove, L.; et al. Reference Gene Validation for RT-qPCR, a Note on Different Available Software Packages. PLoS ONE 2015, 10, e0122515. [Google Scholar] [CrossRef] [Green Version]
- Erdoǧan, F.; Gölgeli, A.; Küçük, A.; Arman, F.; Karaman, Y.; Ersoy, A. Effects of pentylenetetrazole-induced status epilepticus on behavior, emotional memory and learning in immature rats. Epilepsy Behav. 2005, 6, 537–542. [Google Scholar] [CrossRef]
- Nehlig, A.; De Vasconcelos, A.P. The model of pentylenetetrazol-induced status epilepticus in the immature rat: Short- and long-term effects. Epilepsy Res. 1996, 26, 93–103. [Google Scholar] [CrossRef]
- Aniol, V.A.; Ivanova-Dyatlova, A.Y.; Keren, O.; Guekht, A.B.; Sarne, Y.; Gulyaeva, N.V. A single pentylenetetrazole-induced clonic-tonic seizure episode is accompanied by a slowly developing cognitive decline in rats. Epilepsy Behav. 2013, 26, 196–202. [Google Scholar] [CrossRef]
- Kouis, P.; Mikroulis, A.; Psarropoulou, C. A single episode of juvenile status epilepticus reduces the threshold to adult seizures in a stimulus-specific way. Epilepsy Res. 2014, 108, 1564–1571. [Google Scholar] [CrossRef] [PubMed]
- Postnikova, T.Y.; Trofimova, A.M.; Ergina, J.L.; Zubareva, O.E.; Kalemenev, S.V.; Zaitsev, A.V. Transient Switching of NMDA-Dependent Long-Term Synaptic Potentiation in CA3-CA1 Hippocampal Synapses to mGluR 1 -Dependent Potentiation After Pentylenetetrazole-Induced Acute Seizures in Young Rats. Cell. Mol. Neurobiol. 2019, 39, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A. Mapping of neuronal networks underlying generalized seizures induced by increasing doses of pentylenetetrazol in the immature and adult rat: A c-Fos immunohistochemical study. Eur. J. Neurosci. 1998, 10, 2094–2106. [Google Scholar] [CrossRef]
- Van Camp, N.; D’Hooge, R.; Verhoye, M.; Peeters, R.R.; De Deyn, P.P.; Van Der Linden, A. Simultaneous electroencephalographic recording and functional magnetic resonance imaging during pentylenetetrazol-induced seizures in rat. Neuroimage 2003, 19, 627–636. [Google Scholar] [CrossRef]
- Motte, J.E.; da Silva Fernandes, M.J.; Marescaux, C.; Nehlig, A. Effects of pentylenetetrazol-induced status epilepticus on c-Fos and HSP72 immunoreactivity in the immature rat brain. Mol. Brain Res. 1997, 50, 79–84. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2005; ISBN 9780120884728. [Google Scholar]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques 1993, 15, 532–537. [Google Scholar] [CrossRef]
- Schwarz, A.P.; Malygina, D.A.; Kovalenko, A.A.; Trofimov, A.N.; Zaitsev, A.V. Multiplex qPCR assay for assessment of reference gene expression stability in rat tissues/samples. Mol. Cell. Probes 2020, 53, 101611. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Marques, T.E.B.S.; De Mendonça, L.R.; Pereira, M.G.; De Andrade, T.G.; Garcia-Cairasco, N.; Paçó-Larson, M.L.; Gitaí, D.L.G. Validation of Suitable Reference Genes for Expression Studies in Different Pilocarpine-Induced Models of Mesial Temporal Lobe Epilepsy. PLoS ONE 2013, 8, e71892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malkin, S.L.; Amakhin, D.V.; Veniaminova, E.A.; Kim, K.K.; Zubareva, O.E.; Magazanik, L.G.; Zaitsev, A.V. Changes of ampa receptor properties in the neocortex and hippocampus following pilocarpine-induced status epilepticus in rats. Neuroscience 2016. [Google Scholar] [CrossRef] [PubMed]
- Postnikova, T.Y.; Amakhin, D.V.; Trofimova, A.M.; Smolensky, I.V.; Zaitsev, A.V. Changes in Functional Properties of Rat Hippocampal Neurons Following Pentylenetetrazole-induced Status Epilepticus. Neuroscience 2019, 399, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, D.S.; Tumanova, N.L.; Kim, K.K.; Lavrentyeva, V.V.; Lukomskaya, N.Y.; Zhuravin, I.A.; Magazanik, L.G.; Zaitsev, A.V. Transient Morphological Alterations in the Hippocampus After Pentylenetetrazole-Induced Seizures in Rats. Neurochem. Res. 2018, 43, 1671–1682. [Google Scholar] [CrossRef]
- Zaitsev, A.V.; Kim, K.K.; Vasilev, D.S.; Lukomskaya, N.Y.; Lavrentyeva, V.V.; Tumanova, N.L.; Zhuravin, I.A.; Magazanik, L.G. N-methyl-D-aspartate receptor channel blockers prevent pentylenetetrazole-induced convulsions and morphological changes in rat brain neurons. J. Neurosci. Res. 2015. [Google Scholar] [CrossRef]
- Vizuete, A.F.K.; Hennemann, M.M.; Gonçalves, C.A.; De Oliveira, D.L. Phase-Dependent Astroglial Alterations in Li–Pilocarpine-Induced Status Epilepticus in Young Rats. Neurochem. Res. 2017, 42, 2730–2742. [Google Scholar] [CrossRef]
- Cilio, M.R.; Sogawa, Y.; Cha, B.H.; Liu, X.; Huang, L.T.; Holmes, G.L. Long-term effects of status epilepticus in the immature brain are specific for age and model. Epilepsia 2003, 44, 518–528. [Google Scholar] [CrossRef]
- Shaydurov, V.A.; Kasianov, A.; Bolshakov, A.P. Analysis of Housekeeping Genes for Accurate Normalization of qPCR Data During Early Postnatal Brain Development. J. Mol. Neurosci. 2018, 64, 431–439. [Google Scholar] [CrossRef]
- Seol, D.; Choe, H.; Zheng, H.; Jang, K.; Ramakrishnan, P.S.; Lim, T.; Martin, J.A. Selection of reference genes for normalization of quantitative real-time PCR in organ culture of the rat and rabbit intervertebral disc. BMC Res. Notes 2011, 4, 162. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Hunter, T. Metabolic Kinases Moonlighting as Protein Kinases. Trends Biochem. Sci. 2018, 43, 301–310. [Google Scholar] [CrossRef]
- Nicholls, C.; Li, H.; Liu, J.-P. GAPDH: A common enzyme with uncommon functions. Clin. Exp. Pharmacol. Physiol. 2012, 39, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.W.; Lu, P.; Zeng, T.; Tang, H.L.; Chen, Y.H.; Liu, S.J.; Gao, M.M.; Zhao, Q.H.; Yi, Y.H.; Long, Y.S. GAPDH-mediated posttranscriptional regulations of sodium channel Scn1a and Scn3a genes under seizure and ketogenic diet conditions. Neuropharmacology 2017, 113, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Fanselow, M.S.; Dong, H.-W. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroniadou-Anderjaska, V.; Fritsch, B.; Qashu, F.; Braga, M.F.M. Pathology and pathophysiology of the amygdala in epileptogenesis and epilepsy. Epilepsy Res. 2008, 78, 102–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vismer, M.S.; Forcelli, P.A.; Skopin, M.D.; Gale, K.; Koubeissi, M.Z. The piriform, perirhinal, and entorhinal cortex in seizure generation. Front. Neural Circuits 2015, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Isaeva, E.; Romanov, A.; Holmes, G.L.; Isaev, D. Status epilepticus results in region-specific alterations in seizure susceptibility along the hippocampal longitudinal axis. Epilepsy Res. 2015, 110, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Gulyaeva, N.V. Functional Neurochemistry of the Ventral and Dorsal Hippocampus: Stress, Depression, Dementia and Remote Hippocampal Damage. Neurochem. Res. 2019, 44, 1306–1322. [Google Scholar] [CrossRef]
- Lin, W.; Burks, C.A.; Hansen, D.R.; Kinnamon, S.C.; Gilbertson, T.A. Taste receptor cells express pH-sensitive leak K+ channels. J. Neurophysiol. 2004, 92, 2909–2919. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Yamauchi, A.; Nishimura, M.; Ueda, N.; Naito, S. Soybean Oil Fat Emulsion Prevents Cytochrome P450 mRNA Down-Regulation Induced by Fat-Free Overdose Total Parenteral Nutrition in Infant Rats. Biol. Pharm. Bull. 2005, 28, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Bonefeld, B.E.; Elfving, B.; Wegener, G. Reference genes for normalization: A study of rat brain tissue. Synapse 2008, 62, 302–309. [Google Scholar] [CrossRef]
- Cook, N.L.; Vink, R.; Donkin, J.J.; Dan den Heuvel, C. Validation of reference genes for normalization of real-time quantitative RT-PCR data in traumatic brain injury. J. Neurosci. Res. 2009, 87, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Kyzar, E.J.; Zhang, H.; Sakharkar, A.J.; Pandey, S.C. Adolescent alcohol exposure alters lysine demethylase 1 (LSD1) expression and histone methylation in the amygdala during adulthood. Addict. Biol. 2017, 22, 1191–1204. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarz, A.P.; Kovalenko, A.A.; Malygina, D.A.; Postnikova, T.Y.; Zubareva, O.E.; Zaitsev, A.V. Reference Gene Validation in the Brain Regions of Young Rats after Pentylenetetrazole-Induced Seizures. Biomedicines 2020, 8, 239. https://doi.org/10.3390/biomedicines8080239
Schwarz AP, Kovalenko AA, Malygina DA, Postnikova TY, Zubareva OE, Zaitsev AV. Reference Gene Validation in the Brain Regions of Young Rats after Pentylenetetrazole-Induced Seizures. Biomedicines. 2020; 8(8):239. https://doi.org/10.3390/biomedicines8080239
Chicago/Turabian StyleSchwarz, Alexander P., Anna A. Kovalenko, Daria A. Malygina, Tatiana Y. Postnikova, Olga E. Zubareva, and Aleksey V. Zaitsev. 2020. "Reference Gene Validation in the Brain Regions of Young Rats after Pentylenetetrazole-Induced Seizures" Biomedicines 8, no. 8: 239. https://doi.org/10.3390/biomedicines8080239
APA StyleSchwarz, A. P., Kovalenko, A. A., Malygina, D. A., Postnikova, T. Y., Zubareva, O. E., & Zaitsev, A. V. (2020). Reference Gene Validation in the Brain Regions of Young Rats after Pentylenetetrazole-Induced Seizures. Biomedicines, 8(8), 239. https://doi.org/10.3390/biomedicines8080239