The Role of Cutibacterium acnes in Intervertebral Disc Inflammation

 ,
,  and
and 
Abstract

1. Introduction
2. Results
2.1. Prevalence of C. acnes Infection in 30 IVD Biopsies
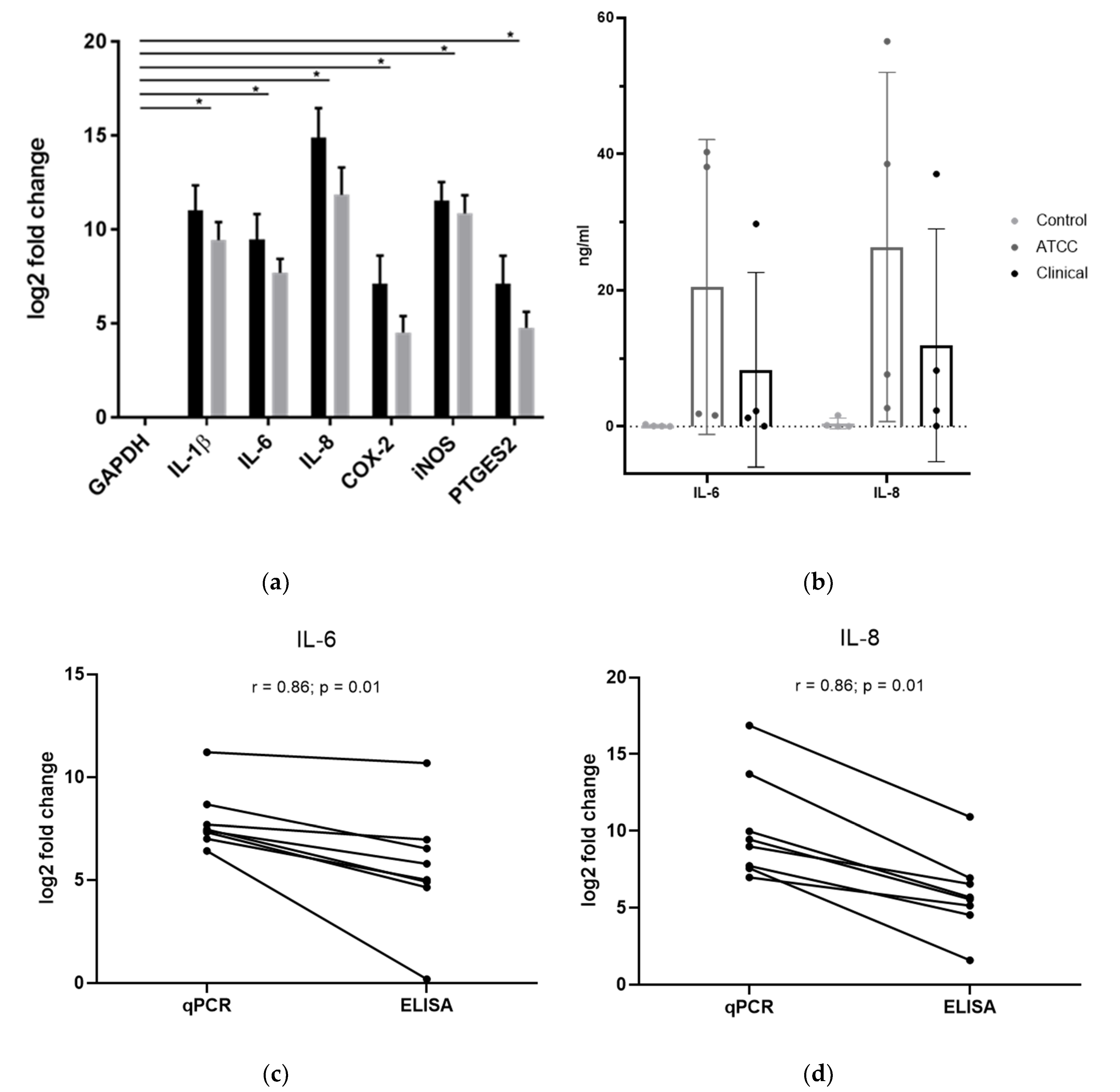
2.2. Changes in IVD Cell Gene Expression upon Exposure to C. acnes
2.3. Changes in IVD Cell Protein Expression upon Exposure to C. acnes
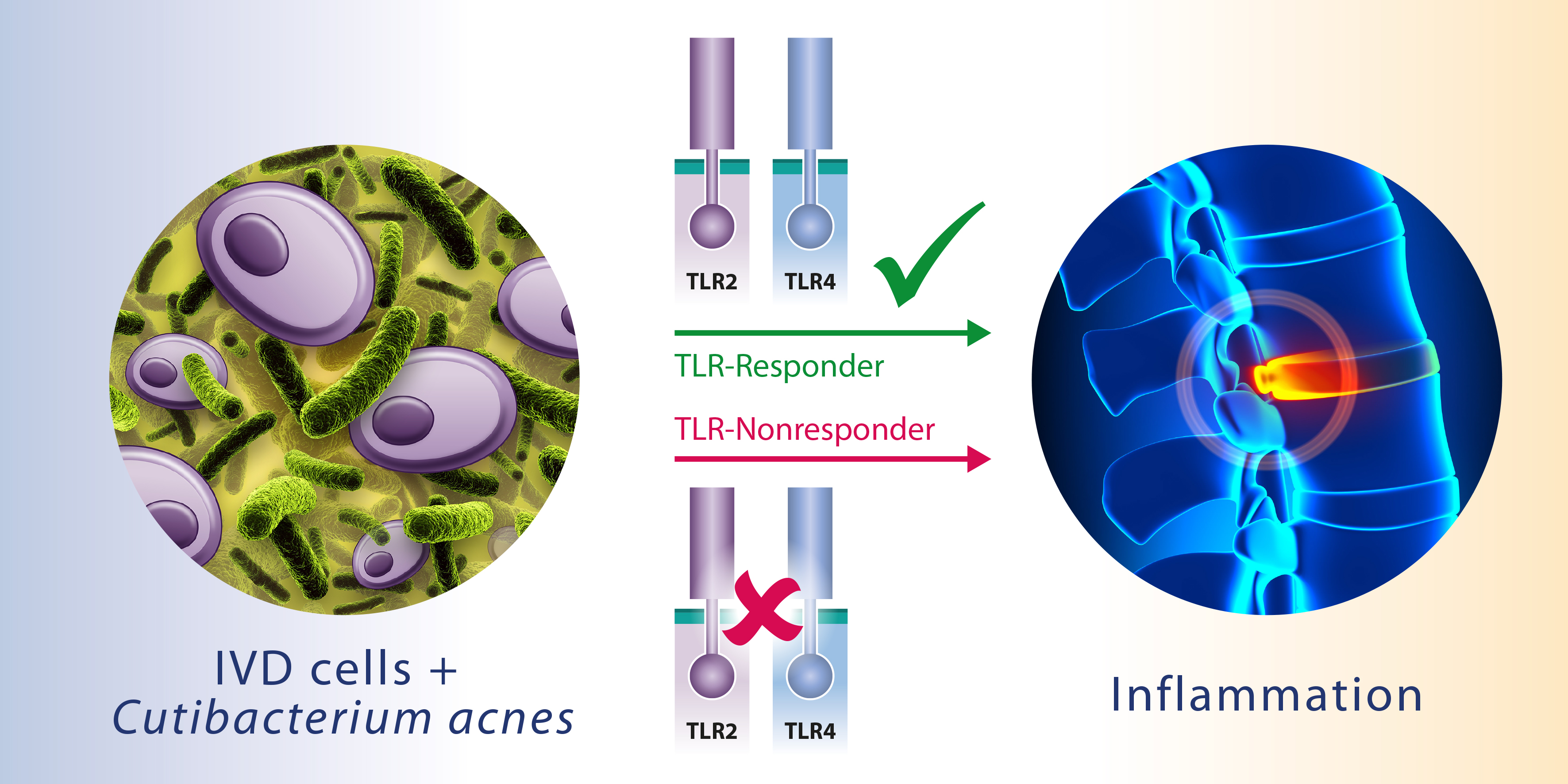
2.4. Involvement of TLR2/4
2.5. Basal Expression of TLR2/4
3. Discussion
4. Materials and Methods
4.1. Detection of C. acnes in Clinical Samples of IVD Biopsies
4.2. Isolation of IVD Cells and Primary Cell Culture
4.3. Bacterial Culture
4.4. Metabolic Activity Assay (MTT)
4.5. Infection of IVD Cells with C. acnes
4.6. Infection of IVD Cells with C. acnes with TLR2/4 Inhibition
4.7. RNA Isolation and Gene Expression Analysis
4.8. ELISA
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IVD | Intervertebral disc |
| C. acnes | Cutibacterium acnes |
| LBP | Low back pain |
| TLR | Toll-like receptor |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| COX | cyclooxygenase |
| DD | Disc degeneration |
| DH | Disc herniation |
| AF | Annulus fibrosus |
| NP | Nucleus pulposus |
| DAMP | Damage-associated molecular pattern |
| PAMP | Pathogen-associated molecular pattern |
| MOI | Multiplicity of infection |
| PTGES2 | Human prostaglandin E synthase 2 |
| JNK | c-Jun N-terminal kinase |
| BHI | Brain Heart Infusion |
| MALDI-TOF | Matrix-assisted laser desorption ionization–time of flight |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| FCS | Fetal calf serum |
| A/A | Antibiotics/antimycotics |
| MTT | 3-[4–dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide formazan |
| PBS | Phosphate Buffered Saline |
References
- Balague, F.; Mannion, A.F.; Pellise, F.; Cedraschi, C. Non-specific low back pain. Lancet 2012, 379, 482–491. [Google Scholar] [CrossRef]
- Wieser, S.; Horisberger, B.; Schmidhauser, S.; Eisenring, C.; Brügger, U.; Ruckstuhl, A.; Dietrich, J.; Mannion, A.F.; Elfering, A.; Tamcan, Ö.; et al. Cost of low back pain in Switzerland in 2005. Eur. J. Health Econ. HEPAC Health Econ. Prev. Care 2011, 12, 455–467. [Google Scholar] [CrossRef]
- Katz, J.N. Lumbar disc disorders and low-back pain: Socioeconomic factors and consequences. J. Bone Jt. Surg. Am. Vol. 2006, 88 (Suppl. 2), 21–24. [Google Scholar] [CrossRef]
- Adams, M.A.; Roughley, P.J. What is intervertebral disc degeneration, and what causes it? Spine 2006, 31, 2151–2161. [Google Scholar] [CrossRef]
- Johnson, Z.I.; Schoepflin, Z.R.; Choi, H.; Shapiro, I.M.; Risbud, M.V. Disc in flames: Roles of TNF-alpha and IL-1beta in intervertebral disc degeneration. Eur. Cells Mater. 2015, 30, 104–116; discussion 16–17. [Google Scholar] [CrossRef]
- Olmarker, K.; Nutu, M.; Storkson, R. Changes in spontaneous behavior in rats exposed to experimental disc herniation are blocked by selective TNF-alpha inhibition. Spine 2003, 28, 1635–1641; discussion 42. [Google Scholar] [CrossRef]
- Molinos, M.; Almeida, C.R.; Caldeira, J.; Cunha, C.; Gonçalves, R.M.; Barbosa, M.A. Inflammation in intervertebral disc degeneration and regeneration. J. R. Soc. Interface 2015, 12, 20141191. [Google Scholar] [CrossRef]
- Wuertz, K.; Haglund, L. Inflammatory mediators in intervertebral disk degeneration and discogenic pain. Glob. Spine J. 2013, 3, 175–184. [Google Scholar] [CrossRef]
- Zhou, Y.Q.; Liu, Z.; Liu, Z.H.; Chen, S.P.; Li, M.; Shahveranov, A.; Ye, D.W.; Tian, Y.K. Interleukin-6: An emerging regulator of pathological pain. J. Neuroinflammation 2016, 13, 141. [Google Scholar] [CrossRef]
- Krock, E.; Currie, J.B.; Weber, M.H.; Ouellet, J.A.; Stone, L.S.; Rosenzweig, D.H.; Haglund, L. Nerve Growth Factor Is Regulated by Toll-Like Receptor 2 in Human Intervertebral Discs. J. Biol. Chem. 2016, 291, 3541–3551. [Google Scholar] [CrossRef]
- Krock, E.; Rosenzweig, D.H.; Currie, J.B.; Bisson, D.G.; Ouellet, J.A.; Haglund, L. Toll-like Receptor Activation Induces Degeneration of Human Intervertebral Discs. Sci. Rep. UK 2017, 7, 17184. [Google Scholar] [CrossRef]
- Quero, L.; Klawitter, M.; Schmaus, A.; Rothley, M.; Sleeman, J.; Tiaden, A.N.; Klasen, J.; Boos, N.; Hottiger, M.O.; Wuertz, K.; et al. Hyaluronic acid fragments enhance the inflammatory and catabolic response in human intervertebral disc cells through modulation of toll-like receptor 2 signalling pathways. Arthritis Res. Ther. 2013, 15, R94. [Google Scholar] [CrossRef]
- Wuertz, K.; Vo, N.; Kletsas, D.; Boos, N. Inflammatory and catabolic signalling in intervertebral discs: The roles of NF-kappaB and MAP kinases. Eur. Cells Mater. 2012, 23, 103–119; discussion 19–20. [Google Scholar] [CrossRef]
- Stirling, A.; Worthington, T.; Rafiq, M.; Lambert, P.A.; Elliott, T.S. Association between sciatica and Propionibacterium acnes. Lancet 2001, 357, 2024–2025. [Google Scholar] [CrossRef]
- Chen, Z.; Cao, P.; Zhou, Z.; Yuan, Y.; Jiao, Y.; Zheng, Y. Overview: The role of Propionibacterium acnes in nonpyogenic intervertebral discs. Int. Orthop. 2016, 40, 1291–1298. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Mollerup, S.; Friis-Nielsen, J.; Vinner, L.; Hansen, T.A.; Richter, S.R.; Fridholm, H.; Herrera, J.A.; Lund, O.; Brunak, S.; Izarzugaza, J.M.; et al. Propionibacterium acnes: Disease-Causing Agent or Common Contaminant? Detection in Diverse Patient Samples by Next-Generation Sequencing. J. Clin. Microbiol. 2016, 54, 980–987. [Google Scholar] [CrossRef]
- Albert, H.B.; Manniche, C.; Sorensen, J.S.; Deleuran, B.W. Antibiotic treatment in patients with low-back pain associated with Modic changes Type 1 (bone oedema): A pilot study. Br. J. Sports Med. 2008, 42, 969–973. [Google Scholar] [CrossRef]
- Albert, H.B.; Sorensen, J.S.; Christensen, B.S.; Manniche, C. Antibiotic treatment in patients with chronic low back pain and vertebral bone edema (Modic type 1 changes): A double-blind randomized clinical controlled trial of efficacy. Eur. Spine J. 2013, 22, 697–707. [Google Scholar] [CrossRef]
- Bråten, L.C.; Rolfsen, M.P.; Espeland, A.; Wigemyr, M.; Aßmus, J.; Froholdt, A.; Haugen, A.J.; Marchand, G.H.; Kristoffersen, P.M.; Lutro, O.; et al. Efficacy of antibiotic treatment in patients with chronic low back pain and Modic changes (the AIM study): Double blind, randomised, placebo controlled, multicentre trial. BMJ 2019, 367, l5654. [Google Scholar] [CrossRef]
- Zhang, B.; Choi, Y.M.; Lee, J.; An, I.S.; Li, L.; He, C.; Dong, Y.; Bae, S.; Meng, H. Toll-like receptor 2 plays a critical role in pathogenesis of acne vulgaris. Biomed. Dermatol. 2019, 3, 4. [Google Scholar] [CrossRef]
- Albert, H.B.; Lambert, P.; Rollason, J.; Sorensen, J.S.; Worthington, T.; Pedersen, M.B.; Nørgaard, H.S.; Vernallis, A.; Busch, F.; Manniche, C.; et al. Does nuclear tissue infected with bacteria following disc herniations lead to Modic changes in the adjacent vertebrae? Eur. Spine J. 2013, 22, 690–696. [Google Scholar] [CrossRef]
- Ahmed-Yahia, S.; Decousser, J.W.; Flouzat-Lachaniette, C.H.; Dervin, G.; Roubineau, F.; Audureau, E.; Hourdille, A.; Royer, G.; Eymard, F.; Chevalier, X. Is the discopathy associated with Modic changes an infectious process? Results from a prospective monocenter study. PLoS ONE 2019, 14, e0221030. [Google Scholar] [CrossRef]
- Yuan, Y.; Chen, Y.; Zhou, Z.; Jiao, Y.; Li, C.; Zheng, Y.; Lin, Y.; Xiao, J.; Chen, Z.; Cao, P. Association between chronic inflammation and latent infection of Propionibacterium acnes in non-pyogenic degenerated intervertebral discs: A pilot study. Eur. Spine J. 2018, 27, 2506–2517. [Google Scholar] [CrossRef] [PubMed]
- Capoor, M.N.; Ruzicka, F.; Machackova, T.; Jancalek, R.; Smrcka, M.; Schmitz, J.E.; Hermanova, M.; Sana, J.; Michu, E.; Baird, J.C.; et al. Prevalence of Propionibacterium acnes in Intervertebral Discs of Patients Undergoing Lumbar Microdiscectomy: A Prospective Cross-Sectional Study. PLoS ONE 2016, 11, e0161676. [Google Scholar] [CrossRef]
- Dudli, S.; Miller, S.; Demir-Deviren, S.; Lotz, J.C. Inflammatory response of disc cells against Propionibacterium acnes depends on the presence of lumbar Modic changes. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2018, 27, 1013–1020. [Google Scholar] [CrossRef]
- Roberts, S.; Evans, H.; Trivedi, J.; Menage, J. Histology and pathology of the human intervertebral disc. J. Bone Jt. Surg. Am. Vol. 2006, 88 (Suppl. 2), 10–14. [Google Scholar]
- Nakazawa, K.R.; Walter, B.A.; Laudier, D.M.; Krishnamoorthy, D.; Mosley, G.E.; Spiller, K.L.; Iatridis, J.C. Accumulation and localization of macrophage phenotypes with human intervertebral disc degeneration. Spine J. Off. J. N. Am. Spine Soc. 2018, 18, 343–356. [Google Scholar] [CrossRef]
- Schubert, A.K.; Smink, J.J.; Arp, M.; Ringe, J.; Hegewald, A.A.; Sittinger, M. uQality Assessment of Surgical Disc Samples Discriminates Human Annulus Fibrosus and Nucleus Pulposus on Tissue and Molecular Level. Int. J. Mol. Sci. 2018, 19, 1761. [Google Scholar] [CrossRef]
- Urban, J.P.; Roberts, S. Degeneration of the intervertebral disc. Arthritis Res. Ther. 2003, 5, 120–130. [Google Scholar] [CrossRef]
- Roszer, T. Understanding the Biology of Self-Renewing Macrophages. Cells 2018, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Klawitter, M.; Hakozaki, M.; Kobayashi, H.; Krupkova, O.; Quero, L.; Ospelt, C.; Gay, S.; Hausmann, O.; Liebscher, T.; Meier, U.; et al. Expression and regulation of toll-like receptors (TLRs) in human intervertebral disc cells. Eur. Spine J. 2014, 23, 1878–1891. [Google Scholar] [CrossRef]
- Klawitter, M.; Quero, L.; Klasen, J.; Gloess, A.N.; Klopprogge, B.; Hausmann, O.; Boos, N.; Wuertz, K. Curcuma DMSO extracts and curcumin exhibit an anti-inflammatory and anti-catabolic effect on human intervertebral disc cells, possibly by influencing TLR2 expression and JNK activity. J. Inflamm. (Lond. Engl.) 2012, 9, 29. [Google Scholar] [CrossRef]
- Kim, J.; Ochoa, M.T.; Krutzik, S.R.; Takeuchi, O.; Uematsu, S.; Legaspi, A.J.; Brightbill, H.D.; Holland, D.; Cunliffe, W.J.; Akira, S.; et al. Activation of toll-like receptor 2 in acne triggers inflammatory cytokine responses. J. Immunol. (Baltim. Md 1950) 2002, 169, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Selway, J.L.; Kurczab, T.; Kealey, T.; Langlands, K. Toll-like receptor 2 activation and comedogenesis: Implications for the pathogenesis of acne. BMC Dermatol. 2013, 13, 10. [Google Scholar] [CrossRef]
- Lin, Y.; Jiao, Y.; Yuan, Y.; Zhou, Z.; Zheng, Y.; Xiao, J.; Li, C.; Chen, Z.; Cao, P. Propionibacterium acnes induces intervertebral disc degeneration by promoting nucleus pulposus cell apoptosis via the TLR2/JNK/mitochondrial-mediated pathway. Emerg. Microbes Infect. 2018, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Brenner, A.K.; Bruserud, O. Functional Toll-Like Receptors (TLRs) Are Expressed by a Majority of Primary Human Acute Myeloid Leukemia Cells and Inducibility of the TLR Signaling Pathway Is Associated with a More Favorable Phenotype. Cancers 2019, 11, 973. [Google Scholar] [CrossRef]
- Achermann, Y.; Tran, B.; Kang, M.; Harro, J.M.; Shirtliff, M.E. Immunoproteomic Identification of In Vivo-Produced Propionibacterium acnes Proteins in a Rabbit Biofilm Infection Model. Clin. Vaccine Immunol. Cvi 2015, 22, 467–476. [Google Scholar] [CrossRef]
- Tafin, U.F.; Corvec, S.; Betrisey, B.; Zimmerli, W.; Trampuz, A. Role of rifampin against Propionibacterium acnes biofilm in vitro and in an experimental foreign-body infection model. Antimicrob. Agents Chemother. 2012, 56, 1885–1891. [Google Scholar] [CrossRef]
- Vignali, D.A. Multiplexed particle-based flow cytometric assays. J. Immunol. Methods 2000, 243, 243–255. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characterization of Patients | n (%) |
|---|---|
| Age, median (range) | 55 (20–78) |
| Sex, female | 12 (40) |
| Smoker | 10 (33) |
| Previous cortisone infiltration | 8 (26.7) |
| Characterization of Biopsies | |
| Spinal Area | |
| Cervical | 11 (36.7) |
| Lumbar | 19 (63.6) |
| Degree of disc degeneration–Pfirrmann Score 1 | |
| II | 3 (10) |
| III | 17 (56.7) |
| IV | 9 (30) |
| V | 1 (3.3) |
| Modic changes 2 | |
| I | 12 (40) |
| II | 15 (50) |
| Unknown | 3 (10) |
| Tissue | |
| Annulus fibrosus | 6 (20) |
| Nucleus pulposus | 19 (63.3) |
| Mix | 5 (16.7) |
| Disease | |
| Disc herniation | 10 (33.3) |
| Disc degeneration | 20 (66.7) |
| Positive Culture Result | 7 (23.3) |
| Cutibacterium acnes | 3 (10) |
| Coagulase-negative staphylococci 3 | 3 (10) |
| Unknown 4 | 1 (3.3) |
| Characterization of Patients | n (%) |
|---|---|
| Age, median (range) | 60 (25–74) |
| Sex, female | 4 (44.5) |
| Characterization of Biopsies | |
| Spinal Area | |
| Cervical | 1 (11.1) |
| Lumbar | 8 (88.9) |
| Degree of disc degeneration–Pfirrmann Score 1 | |
| II | 2 (22.2) |
| III | 3 (33.3) |
| IV | 4 (44.5) |
| Modic changes 2 | |
| 0 | 2 (22.2) |
| I | 2 (22.2) |
| II | 4 (44.5) |
| Unknown | 1 (11.1) |
| Diagnosis | |
| Disc herniation | 5 (55.6) |
| Disc degeneration 3 | 4 (44.4) |
| Target Gene | Assay Identification Number |
|---|---|
| COX-2 | Hs00153133_m1 |
| GAPDH | Hs02758991_g1 |
| IL-1β | Hs00174097_m1 |
| IL-6 | Hs00174131_m1 |
| IL-8 | Hs00174103_m1 |
| iNOS | Hs01075521_m1 |
| PTGES2 | Hs00228159_m1 |
| TLR2 | Hs00152932_m1 |
| TLR4 | Hs00152939_m1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmid, B.; Hausmann, O.; Hitzl, W.; Achermann, Y.; Wuertz-Kozak, K. The Role of Cutibacterium acnes in Intervertebral Disc Inflammation. Biomedicines 2020, 8, 186. https://doi.org/10.3390/biomedicines8070186
Schmid B, Hausmann O, Hitzl W, Achermann Y, Wuertz-Kozak K. The Role of Cutibacterium acnes in Intervertebral Disc Inflammation. Biomedicines. 2020; 8(7):186. https://doi.org/10.3390/biomedicines8070186
Chicago/Turabian StyleSchmid, Bettina, Oliver Hausmann, Wolfgang Hitzl, Yvonne Achermann, and Karin Wuertz-Kozak. 2020. "The Role of Cutibacterium acnes in Intervertebral Disc Inflammation" Biomedicines 8, no. 7: 186. https://doi.org/10.3390/biomedicines8070186
APA StyleSchmid, B., Hausmann, O., Hitzl, W., Achermann, Y., & Wuertz-Kozak, K. (2020). The Role of Cutibacterium acnes in Intervertebral Disc Inflammation. Biomedicines, 8(7), 186. https://doi.org/10.3390/biomedicines8070186