Effect of the Most Relevant CYP3A4 and CYP3A5 Polymorphisms on the Pharmacokinetic Parameters of 10 CYP3A Substrates

 ,
,  ,
, 

Abstract
1. Introduction
1.1. CYP3A Subfamily
1.2. CYP3A4 and CYP3A5
2. Materials and Methods
2.1. Study Population and Study Design
2.2. Pharmacokinetic Analysis
2.3. Genotyping
2.4. Statistical Analysis
3. Results and Discussion
Study Limitations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- König, I.R.; Fuchs, O.; Hansen, G.; von Mutius, E.; Kopp, M.V. What is precision medicine? Eur. Respir. J. 2017, 50. [Google Scholar] [CrossRef] [PubMed]
- Weinshilboum, R.; Wang, L. Pharmacogenomics: Bench to bedside. Nat. Rev. Drug Discov. 2004, 3, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Motulsky, A.G. Drug reactions, enzymes, and biochemical genetics. J. Am. Med. Assoc. 1957, 165, 835–837. [Google Scholar] [CrossRef]
- Moderne Probleme der Human Genetik. Ergebn. inn-Google Académico. Available online: https://scholar.google.es/scholar?hl=es&as_sdt=0%2C5&q=Moderne+Probleme+der+Human+genetik.+Ergebn.+inn&btnG= (accessed on 24 February 2020).
- US Food and Drug Administration. Clinical Pharmacogenomics. Premarket Evaluation in Early-Phase Clinical Studies and Recommendations for Labeling; Silver Spring: Rockville, MD, USA, 2013; Volume 26.
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharm. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Feyereisen, R.; Nelson, D.R.; Coon, M.J.; Estabrook, R.W.; Nebert, D.W.; Fujii-kuriyama, Y.; Gonzalez, F.J.; Guengerich, F.P.; Gunsalus, I.C.; Johnson, E.F. The P450 superfamily: Update on new sequences, gene mapping, and recommended nomenclature. DNA Cell Biol. 1991, 10, 1–14. [Google Scholar]
- Ingelman-Sundberg, M.; Rodriguez-Antona, C. Pharmacogenetics of drug-metabolizing enzymes: Implications for a safer and more effective drug therapy. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1563–1570. [Google Scholar] [CrossRef]
- Rostami-Hodjegan, A.; Amin, A.M.; Spencer, E.P.; Lennard, M.S.; Tucker, G.T.; Flanagan, R.J. Influence of dose, cigarette smoking, age, sex, and metabolic activity on plasma clozapine concentrations: A predictive model and nomograms to aid clozapine dose adjustment and to assess compliance in individual patients. J. Clin. Psychopharmacol. 2004, 24, 70–78. [Google Scholar] [CrossRef]
- Domanski, T.L.; Finta, C.; Halpert, J.R.; Zaphiropoulos, P.G. cDNA cloning and initial characterization of CYP3A43, a novel human cytochrome P450. Mol. Pharmacol. 2001, 59, 386–392. [Google Scholar] [CrossRef]
- Shimada, T.; Yamazaki, H.; Mimura, M.; Inui, Y.; Guengerich, F.P. Interindividual variations in human liver cytochrome P-450 enzymes involved in the oxidation of drugs, carcinogens and toxic chemicals: Studies with liver microsomes of 30 Japanese and 30 Caucasians. J. Pharmacol. Exp. Ther. 1994, 270, 414–423. [Google Scholar]
- Brooks, B.A.; McBride, O.W.; Dolphin, C.T.; Farrall, M.; Scambler, P.J.; Gonzalez, F.J.; Idle, J.R. The gene CYP3 encoding P450pcn1 (nifedipine oxidase) is tightly linked to the gene COL1A2 encoding collagen type 1 alpha on 7q21-q22. 1. Am. J. Hum. Genet. 1988, 43, 280. [Google Scholar]
- Finta, C.; Zaphiropoulos, P.G. The human cytochrome P450 3A locus. Gene evolution by capture of downstream exons. Gene 2000, 260, 13–23. [Google Scholar] [CrossRef]
- Hashimoto, H.; Toide, K.; Kitamura, R.; Fujita, M.; Tagawa, S.; Itoh, S.; Kamataki, T. Gene structure of CYP3A4, an adult-specific form of cytochrome P450 in human livers, and its transcriptional control. Eur. J. Bioch. 1993, 218, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Gellner, K.; Eiselt, R.; Hustert, E.; Arnold, H.; Koch, I.; Haberl, M.; Deglmann, C.J.; Burk, O.; Buntefuss, D.; Escher, S.; et al. Genomic organization of the human CYP3A locus: Identification of a new, inducible CYP3A gene. Pharm. Genom. 2001, 11, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Plant, N. The human cytochrome P450 sub-family: Transcriptional regulation, inter-individual variation and interaction networks. Biochim. Biophys. Acta 2007, 1770, 478–488. [Google Scholar] [CrossRef]
- Elens, L.; Van Gelder, T.; Hesselink, D.A.; Haufroid, V.; Van Schaik, R.H. CYP3A4* 22: Promising newly identified CYP3A4 variant allele for personalizing pharmacotherapy. Pharmacogenomics 2013, 14, 47–62. [Google Scholar] [CrossRef]
- Daly, A.K. Significance of the minor cytochrome P450 3A isoforms. Clin. Pharm. 2006, 45, 13–31. [Google Scholar] [CrossRef]
- Shimada, T.; Yamazaki, H.; Mimura, M.; Wakamiya, N.; Ueng, Y.F.; Guengerich, F.P.; Inui, Y. Characterization of microsomal cytochrome P450 enzymes involved in the oxidation of xenobiotic chemicals in human fetal liver and adult lungs. Drug Metab. Dispos. 1996, 24, 515–522. [Google Scholar]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Williams, J.A.; Ring, B.J.; Cantrell, V.E.; Jones, D.R.; Eckstein, J.; Ruterbories, K.; Hamman, M.A.; Hall, S.D.; Wrighton, S.A. Comparative metabolic capabilities of CYP3A4, CYP3A5, and CYP3A7. Drug Metab. Dispos. 2002, 30, 883–891. [Google Scholar] [CrossRef]
- Huang, W.; Lin, Y.S.; McConn, D.J.; Calamia, J.C.; Totah, R.A.; Isoherranen, N.; Glodowski, M.; Thummel, K.E. Evidence of significant contribution from CYP3A5 to hepatic drug metabolism. Drug Metab. Dispos. 2004, 32, 1434–1445. [Google Scholar] [CrossRef]
- Kivistö, K.T.; Bookjans, G.; Fromm, M.F.; Griese, E.-U.; Münzel, P.; KRoemer, H.K. Expression of CYP3A4, CYP3A5 and CYP3A7 in human duodenal tissue. Br. J. Clin. Pharm. 1996, 42, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Kleinbloesem, C.H.; van Brummelen, P.; Faber, H.; Danhof, M.; Vermeulen, N.P.; Breimer, D.D. Variability in nifedipine pharmacokinetics and dynamics: A new oxidation polymorphism in man. Biochem. Pharmacol. 1984, 33, 3721–3724. [Google Scholar] [CrossRef]
- Guengerich, F.P.; Martin, M.V.; Beaune, P.H.; Kremers, P.; Wolff, T.; Waxman, D.J. Characterization of rat and human liver microsomal cytochrome P-450 forms involved in nifedipine oxidation, a prototype for genetic polymorphism in oxidative drug metabolism. J. Biol. Chem. 1986, 261, 5051–5060. [Google Scholar] [PubMed]
- Inoue, K.; Inazawa, J.; Nakagawa, H.; Shimada, T.; Yamazaki, H.; Guengerich, F.P.; Abe, T. Assignment of the human cytochrome P-450 nifedipine oxidase gene (CYP3A4) to chromosome 7 at band q22. 1 by fluorescencein situ hybridization. Jpn. Hum. Genet. 1992, 37, 133–138. [Google Scholar] [CrossRef]
- The Nucleotide Database. Nucleotide–NCBI. Available online: https://www.ncbi.nlm.nih.gov/nuccore/197313719?report=graph (accessed on 24 February 2020).
- Ensembl Genome Browser. Available online: http://www.ensembl.org/Homo_sapiens/Gene/Summary?db=core;g=ENSG00000160868;r=7:99756960-99784248;t=ENST00000415003 (accessed on 24 February 2020).
- Achour, B.; Barber, J.; Rostami-Hodjegan, A. Expression of hepatic drug-metabolizing cytochrome p450 enzymes and their intercorrelations: A meta-analysis. Drug Metab. Dispos. 2014, 42, 1349–1356. [Google Scholar] [CrossRef]
- Michaels, S.; Wang, M.Z. The Revised Human Liver Cytochrome P450 “Pie”: Absolute Protein Quantification of CYP4F and CYP3A Enzymes Using Targeted Quantitative Proteomics. Drug Metab. Dispos. 2014, 42, 1241–1251. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Dunbar, D.; Ostrowska, A.; Zeisloft, S.; Yang, J.; Kaminsky, L.S. Characterization of human small intestinal cytochromes P-450. Drug Metab. Dispos. 1999, 27, 804–809. [Google Scholar]
- Paine, M.F.; Shen, D.D.; Kunze, K.L.; Perkins, J.D.; Marsh, C.L.; McVicar, J.P.; Barr, D.M.; Gillies, B.S.; Thummel, K.E. First-pass metabolism of midazolam by the human intestine. Clin. Pharm. Ther. 1996, 60, 14–24. [Google Scholar] [CrossRef]
- Kolars, J.C.; Watkins, P.B.; Merion, R.M.; Awni, W.M. First-pass metabolism of cyclosporin by the gut. Lancet 1991, 338, 1488–1490. [Google Scholar] [CrossRef]
- Westlind-Johnsson, A.; Hermann, R.; Huennemeyer, A.; Hauns, B.; Lahu, G.; Nassr, N.; Zech, K.; Ingelman-Sundberg, M.; von Richter, O. Identification and characterization of CYP3A4*20, a novel rare CYP3A4 allele without functional activity. Clin. Pharmacol. Ther. 2006, 79, 339–349. [Google Scholar] [CrossRef]
- Wang, D.; Guo, Y.; Wrighton, S.A.; Cooke, G.E.; Sadee, W. Intronic polymorphism in CYP3A4 affects hepatic expression and response to statin drugs. Pharm. J. 2011, 11, 274–286. [Google Scholar] [CrossRef]
- Klein, K.; Thomas, M.; Winter, S.; Nussler, A.K.; Niemi, M.; Schwab, M.; Zanger, U.M. PPARA: A novel genetic determinant of CYP3A4 in vitro and in vivo. Clin. Pharm. Ther. 2012, 91, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Apellániz-Ruiz, M.; Inglada-Pérez, L.; Naranjo, M.E.G.; Sánchez, L.; Mancikova, V.; Currás-Freixes, M.; de Cubas, A.A.; Comino-Méndez, I.; Triki, S.; Rebai, A.; et al. High frequency and founder effect of the CYP3A4*20 loss-of-function allele in the Spanish population classifies CYP3A4 as a polymorphic enzyme. Pharm. J. 2015, 15, 288–292. [Google Scholar] [CrossRef]
- Tavira, B.; Coto, E.; Diaz-Corte, C.; Alvarez, V.; López-Larrea, C.; Ortega, F. A search for new CYP3A4 variants as determinants of tacrolimus dose requirements in renal-transplanted patients. Pharm. Genom. 2013, 23, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.B.; Struchiner, C.J.; Felipe, C.R.; Tedesco-Silva, H.; Medina-Pestana, J.O.; Suarez-Kurtz, G. CYP3A5 genotype, but not CYP3A4*1b, CYP3A4*22, or hematocrit, predicts tacrolimus dose requirements in Brazilian renal transplant patients. Clin. Pharmacol. Ther. 2013, 94, 201–202. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.; Felipe, C.R.; Tedesco-Silva, H.; Medina-Pestana, J.O.; Struchiner, C.J.; Ojopi, E.B.; Suarez-Kurtz, G. Pharmacogenetics of calcineurin inhibitors in Brazilian renal transplant patients. Pharmacogenomics 2011, 12, 1293–1303. [Google Scholar] [CrossRef]
- Kurzawski, M.; Dąbrowska, J.; Dziewanowski, K.; Domański, L.; Perużyńska, M.; Droździk, M. CYP3A5 and CYP3A4, but not ABCB1 polymorphisms affect tacrolimus dose-adjusted trough concentrations in kidney transplant recipients. Pharmacogenomics 2014, 15, 179–188. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, H.; Elens, L.; de Loor, H.; van Schaik, R.H.; Kuypers, D.R.J. The CYP3A4*22 C>T single nucleotide polymorphism is associated with reduced midazolam and tacrolimus clearance in stable renal allograft recipients. Pharm. J. 2015, 15, 144–152. [Google Scholar] [CrossRef]
- Elens, L.; van Schaik, R.H.; Panin, N.; de Meyer, M.; Wallemacq, P.; Lison, D.; Mourad, M.; Haufroid, V. Effect of a new functional CYP3A4 polymorphism on calcineurin inhibitors’ dose requirements and trough blood levels in stable renal transplant patients. Pharmacogenomics 2011, 12, 1383–1396. [Google Scholar] [CrossRef]
- Elens, L.; Becker, M.L.; Haufroid, V.; Hofman, A.; Visser, L.E.; Uitterlinden, A.G.; Stricker, B.C.; van Schaik, R.H.N. Novel CYP3A4 intron 6 single nucleotide polymorphism is associated with simvastatin-mediated cholesterol reduction in the Rotterdam Study. Pharm. Genom. 2011, 21, 861–866. [Google Scholar] [CrossRef]
- Rebbeck, T.R.; Jaffe, J.M.; Walker, A.H.; Wein, A.J.; Malkowicz, S.B. Modification of clinical presentation of prostate tumors by a novel genetic variant in CYP3A4. J. Natl. Cancer Inst. 1998, 90, 1225–1229. [Google Scholar] [CrossRef] [PubMed]
- Westlind, A.; Löfberg, L.; Tindberg, N.; Andersson, T.B.; Ingelman-Sundberg, M. Interindividual differences in hepatic expression of CYP3A4: Relationship to genetic polymorphism in the 5′-upstream regulatory region. Biochem. Biophys. Res. Commun. 1999, 259, 201–205. [Google Scholar] [CrossRef]
- Rodríguez-Antona, C.; Sayi, J.G.; Gustafsson, L.L.; Bertilsson, L.; Ingelman-Sundberg, M. Phenotype-genotype variability in the human CYP3A locus as assessed by the probe drug quinine and analyses of variant CYP3A4 alleles. Biochem. Biophys. Res. Commun. 2005, 338, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Amirimani, B.; Ning, B.; Deitz, A.C.; Weber, B.L.; Kadlubar, F.F.; Rebbeck, T.R. Increased transcriptional activity of the CYP3A4*1B promoter variant. Environ. Mol. Mutagen. 2003, 42, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Hesselink, D.A.; van Schaik, R.H.N.; van der Heiden, I.P.; van der Werf, M.; Gregoor, P.J.H.S.; Lindemans, J.; Weimar, W.; van Gelder, T. Genetic polymorphisms of the CYP3A4, CYP3A5, and MDR-1 genes and pharmacokinetics of the calcineurin inhibitors cyclosporine and tacrolimus. Clin. Pharmacol. Ther. 2003, 74, 245–254. [Google Scholar] [CrossRef]
- Tavira, B.; Garciá, E.C.; Díaz-Corte, C.; Ortega, F.; Arias, M.; Torres, A.; Díaz, J.M.; Selgas, R.; López-Larrea, C.; Campistol, J.M. Pharmacogenetics of tacrolimus after renal transplantation: Analysis of polymorphisms in genes encoding 16 drug metabolizing enzymes. Clin. Chem. Lab. Med. 2011, 49, 825–833. [Google Scholar] [CrossRef]
- Gervasini, G.; Garcia, M.; Macias, R.M.; Cubero, J.J.; Caravaca, F.; Benitez, J. Impact of genetic polymorphisms on tacrolimus pharmacokinetics and the clinical outcome of renal transplantation. Transpl. Int. 2012, 25, 471–480. [Google Scholar] [CrossRef]
- Crettol, S.; Venetz, J.-P.; Fontana, M.; Aubert, J.-D.; Pascual, M.; Eap, C.B. CYP3A7, CYP3A5, CYP3A4, and ABCB1 genetic polymorphisms, cyclosporine concentration, and dose requirement in transplant recipients. Ther. Drug Monit. 2008, 30, 689–699. [Google Scholar] [CrossRef]
- Żochowska, D.; Wyzgał, J.; Pączek, L. Impact of CYP3A4*1B and CYP3A5*3 polymorphisms on the pharmacokinetics of cyclosporine and sirolimus in renal transplant recipients. Ann. Transplant. 2012, 17, 36–44. [Google Scholar] [CrossRef]
- Gervasini, G.; García-Pino, G.; Vergara, E.; Mota-Zamorano, S.; García-Cerrada, M.; Luna, E. CYP3A genotypes of donors but not those of the patients increase the risk of acute rejection in renal transplant recipients on calcineurin inhibitors: A pilot study. Eur. J. Clin. Pharmacol. 2018, 74, 53–60. [Google Scholar] [CrossRef]
- Becker, M.L.; Visser, L.E.; van Schaik, R.H.N.; Hofman, A.; Uitterlinden, A.G.; Stricker, B.H.C. Influence of genetic variation in CYP3A4 and ABCB1 on dose decrease or switching during simvastatin and atorvastatin therapy. Pharmacoepidemiol. Drug Saf. 2010, 19, 75–81. [Google Scholar] [CrossRef]
- Birdwell, K.A.; Grady, B.; Choi, L.; Xu, H.; Bian, A.; Denny, J.C.; Jiang, M.; Vranic, G.; Basford, M.; Cowan, J.D.; et al. The use of a DNA biobank linked to electronic medical records to characterize pharmacogenomic predictors of tacrolimus dose requirement in kidney transplant recipients. Pharm. Genom. 2012, 22, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Tang, J.; Rose, R.; Hodgson, E.; Bienstock, R.J.; Mohrenweiser, H.W.; Goldstein, J.A. Identification of variants of CYP3A4 and characterization of their abilities to metabolize testosterone and chlorpyrifos. J. Pharm. Exp. Ther. 2001, 299, 825–831. [Google Scholar]
- Fukushima-Uesaka, H.; Saito, Y.; Watanabe, H.; Shiseki, K.; Saeki, M.; Nakamura, T.; Kurose, K.; Sai, K.; Komamura, K.; Ueno, K.; et al. Haplotypes of CYP3A4 and their close linkage with CYP3A5 haplotypes in a Japanese population. Hum. Mutat. 2004, 23, 100. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.; Ng, C.M.; Barrett, J.S.; Luo, A.; Zhang, B.; Deng, C.; Xi, L.; Cheng, K.; Ming, Y.; Yang, G.; et al. Effects of CYP3A4 and CYP3A5 polymorphisms on tacrolimus pharmacokinetics in Chinese adult renal transplant recipients: A population pharmacokinetic analysis. Pharm. Genom. 2013, 23, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, M.; Hosokawa, M.; Shinke, H.; Hashimoto, E.; Takahashi, T.; Kawai, T.; Matsubara, K.; Ogawa, K.; Fujimoto, Y.; Okamoto, S.; et al. Influence of cytochrome P450 (CYP) 3A4*1G polymorphism on the pharmacokinetics of tacrolimus, probability of acute cellular rejection, and mRNA expression level of CYP3A5 rather than CYP3A4 in living-donor liver transplant patients. Biol. Pharm. Bull. 2013, 36, 1814–1821. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-J.; Li, L.; Lin, L.; Jiang, H.-X.; Zhong, Z.-Y.; Li, W.-M.; Zhang, Y.-J.; Zheng, P.; Tan, X.-H.; Zhou, L. Impact of the CYP3A5, CYP3A4, COMT, IL-10 and POR genetic polymorphisms on tacrolimus metabolism in Chinese renal transplant recipients. PLoS ONE 2014, 9, e86206. [Google Scholar] [CrossRef]
- Li, J.; Liu, S.; Fu, Q.; Zhang, Y.; Wang, X.; Liu, X.; Liu, L.; Wang, C.; Huang, M. Interactive effects of CYP3A4, CYP3A5, MDR1 and NR1I2 polymorphisms on tracrolimus trough concentrations in early postrenal transplant recipients. Pharmacogenomics 2015, 16, 1355–1365. [Google Scholar] [CrossRef]
- Liu, M.; He, H.; Zhang, Y.; Hu, Y.; He, F.; Luo, J.; Luo, Z.; Chen, X.; Liu, Z.; Zhou, H.; et al. IL-3 and CTLA4 gene polymorphisms may influence the tacrolimus dose requirement in Chinese kidney transplant recipients. Acta Pharm. Sin. 2017, 38, 415–423. [Google Scholar] [CrossRef]
- Sun, B.; Guo, Y.; Gao, J.; Shi, W.; Fan, G.; Li, X.; Qiu, J.; Qin, Y.; Liu, G. Influence of CYP3A and ABCB1 polymorphisms on cyclosporine concentrations in renal transplant recipients. Pharmacogenomics 2017, 18, 1503–1513. [Google Scholar] [CrossRef]
- Li, T.-F.; Hu, L.; Ma, X.-L.; Huang, L.; Liu, X.-M.; Luo, X.-X.; Feng, W.-Y.; Wu, C.-F. Population pharmacokinetics of cyclosporine in Chinese children receiving hematopoietic stem cell transplantation. Acta Pharm. Sin. 2019, 40, 1603–1610. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chang, Y.-Z.; Kan, Q.-C.; Zhang, L.-R.; Li, Z.-S.; Lu, H.; Wang, Z.-Y.; Chu, Q.-J.; Zhang, J. CYP3A4*1G genetic polymorphism influences CYP3A activity and response to fentanyl in Chinese gynecologic patients. Eur. J. Clin. Pharm. 2010, 66, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Zhang, X.; Deng, Q.; Wu, Y.; Xiang, G. Impact of CYP3A4*1G polymorphism on metabolism of fentanyl in Chinese patients undergoing lower abdominal surgery. Clin. Chim. Acta 2011, 412, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yuan, J.-J.; Kan, Q.-C.; Zhang, L.-R.; Chang, Y.-Z.; Wang, Z.-Y.; Li, Z.-S. Influence of CYP3A5*3 polymorphism and interaction between CYP3A5*3 and CYP3A4*1G polymorphisms on post-operative fentanyl analgesia in Chinese patients undergoing gynaecological surgery. Eur. J. Anaesthesiol. 2011, 28, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.-L.; Li, H.; Chen, Q.-X.; Hu, Y.; Wu, S.-J.; Tang, L.-Y.; Gong, W.-Y.; Xie, G.-H.; Fang, X.-M. Effect of CYP3A4*1G on the fentanyl consumption for intravenous patient-controlled analgesia after total abdominal hysterectomy in Chinese Han population. J. Clin. Pharm. Ther. 2012, 37, 153–156. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, L.; Zhao, X.; Shen, S.; Luo, X.; Zhang, Y. Association between MDR1/CYP3A4/OPRM1 gene polymorphisms and the post-caesarean fentanyl analgesic effect on Chinese women. Gene 2018, 661, 78–84. [Google Scholar] [CrossRef]
- Sata, F.; Sapone, A.; Elizondo, G.; Stocker, P.; Miller, V.P.; Zheng, W.; Raunio, H.; Crespi, C.L.; Gonzalez, F.J. CYP3A4 allelic variants with amino acid substitutions in exons 7 and 12: Evidence for an allelic variant with altered catalytic activity. Clin. Pharmacol. Ther. 2000, 67, 48–56. [Google Scholar] [CrossRef]
- Thompson, J.F.; Man, M.; Johnson, K.J.; Wood, L.S.; Lira, M.E.; Lloyd, D.B.; Banerjee, P.; Milos, P.M.; Myrand, S.P.; Paulauskis, J.; et al. An association study of 43 SNPs in 16 candidate genes with atorvastatin response. Pharm. J. 2005, 5, 352–358. [Google Scholar] [CrossRef]
- Ensembl Genome Browser. Available online: http://www.ensembl.org/Homo_sapiens/Gene/Summary?db=core;g=ENSG00000106258;r=7:99648194-99679998 (accessed on 24 February 2020).
- Kuehl, P.; Zhang, J.; Lin, Y.; Lamba, J.; Assem, M.; Schuetz, J.; Watkins, P.B.; Daly, A.; Wrighton, S.A.; Hall, S.D. Sequence diversity in CYP3A promoters and characterization of the genetic basis of polymorphic CYP3A5 expression. Nat. Genet. 2001, 27, 383–391. [Google Scholar] [CrossRef]
- Lamba, J.K.; Lin, Y.S.; Schuetz, E.G.; Thummel, K.E. Genetic contribution to variable human CYP3A-mediated metabolism. Adv. Drug Deliv. Rev. 2002, 54, 1271–1294. [Google Scholar] [CrossRef]
- Miao, J.; Jin, Y.; Marunde, R.L.; Kim, S.; Quinney, S.; Radovich, M.; Li, L.; Hall, S.D. Association of genotypes of the CYP3A cluster with midazolam disposition in vivo. Pharm. J. 2009, 9, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Moes, D.J.A.R.; Swen, J.J.; den Hartigh, J.; van der Straaten, T.; van der Heide, J.J.H.; Sanders, J.S.; Bemelman, F.J.; de Fijter, J.W.; Guchelaar, H.J. Effect of CYP3A4*22, CYP3A5*3, and CYP3A Combined Genotypes on Cyclosporine, Everolimus, and Tacrolimus Pharmacokinetics in Renal Transplantation. CPT Pharm. Syst. Pharmacol. 2014, 3, e100. [Google Scholar] [CrossRef] [PubMed]
- Kitzmiller, J.P.; Sullivan, D.M.; Phelps, M.A.; Wang, D.; Sadee, W. CYP3A4/5 combined genotype analysis for predicting statin dose requirement for optimal lipid control. Drug Metab. Drug Interact. 2013, 28, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Sanchez Spitman, A.B.; Moes, D.J.A.R.; Gelderblom, H.; Dezentje, V.O.; Swen, J.J.; Guchelaar, H.J. Effect of CYP3A4*22, CYP3A5*3, and CYP3A combined genotypes on tamoxifen metabolism. Eur. J. Clin. Pharmacol. 2017, 73, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Hustert, E.; Haberl, M.; Burk, O.; Wolbold, R.; He, Y.Q.; Klein, K.; Nuessler, A.C.; Neuhaus, P.; Klattig, J.; Eiselt, R.; et al. The genetic determinants of the CYP3A5 polymorphism. Pharmacogenetics 2001, 11, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Usmani, K.A.; Chanas, B.; Ghanayem, B.; Xi, T.; Hodgson, E.; Mohrenweiser, H.W.; Goldstein, J.A. Genetic findings and functional studies of human CYP3A5 single nucleotide polymorphisms in different ethnic groups. Pharmacogenetics 2003, 13, 461–472. [Google Scholar] [CrossRef]
- Birdwell, K.A.; Decker, B.; Barbarino, J.M.; Peterson, J.F.; Stein, C.M.; Sadee, W.; Wang, D.; Vinks, A.A.; He, Y.; Swen, J.J. Clinical Pharmacogenetics Implementation Consortium (CPIC) guidelines for CYP3A5 genotype and tacrolimus dosing. Clin. Pharm. Ther. 2015, 98, 19–24. [Google Scholar] [CrossRef]
- Tsamandouras, N.; Dickinson, G.; Guo, Y.; Hall, S.; Rostami-Hodjegan, A.; Galetin, A.; Aarons, L. Identification of the effect of multiple polymorphisms on the pharmacokinetics of simvastatin and simvastatin acid using a population-modeling approach. Clin. Pharmacol. Ther. 2014, 96, 90–100. [Google Scholar] [CrossRef]
- Luzum, J.A.; Theusch, E.; Taylor, K.D.; Wang, A.; Sadee, W.; Binkley, P.F.; Krauss, R.M.; Medina, M.W.; Kitzmiller, J.P. Individual and Combined Associations of Genetic Variants in CYP3A4, CYP3A5, and SLCO1B1 With Simvastatin and Simvastatin Acid Plasma Concentrations. J. Cardiovasc. Pharmacol. 2015, 66, 80–85. [Google Scholar] [CrossRef]
- Prueksaritanont, T.; Gorham, L.M.; Ma, B.; Liu, L.; Yu, X.; Zhao, J.J.; Slaughter, D.E.; Arison, B.H.; Vyas, K.P. In vitro metabolism of simvastatin in humans [SBT] identification of metabolizing enzymes and effect of the drug on hepatic P450s. Drug Metab. Dispos. 1997, 25, 1191–1199. [Google Scholar]
- Westlind-Johnsson, A.; Malmebo, S.; Johansson, A.; Otter, C.; Andersson, T.B.; Johansson, I.; Edwards, R.J.; Boobis, A.R.; Ingelman-Sundberg, M. Comparative analysis of CYP3A expression in human liver suggests only a minor role for CYP3A5 in drug metabolism. Drug Metab. Dispos. 2003, 31, 755–761. [Google Scholar] [CrossRef]
- Wolbold, R.; Klein, K.; Burk, O.; Nüssler, A.K.; Neuhaus, P.; Eichelbaum, M.; Schwab, M.; Zanger, U.M. Sex is a major determinant of CYP3A4 expression in human liver. Hepatology 2003, 38, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Krecic-Shepard, M.E.; Park, K.; Barnas, C.; Slimko, J.; Kerwin, D.R.; Schwartz, J.B. Race and sex influence clearance of nifedipine: Results of a population study. Clin. Pharm. Ther. 2000, 68, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, T.N.; Klepstad, P.; Davies, A.; Bjordal, K.; Lundström, S.; Kaasa, S.; Dale, O. Influences on the pharmacokinetics of oxycodone: A multicentre cross-sectional study in 439 adult cancer patients. Eur. J. Clin. Pharmacol. 2011, 67, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Kahan, B.D.; Kramer, W.G.; Wideman, C.; Flechner, S.M.; Lorber, M.I.; Van, C.B. Demographic factors affecting the pharmacokinetics of cyclosporine estimated by radioimmunoassay. Transplantation 1986, 41, 459–464. [Google Scholar] [CrossRef]
- Zhu, B.; Liu, Z.-Q.; Chen, G.-L.; Chen, X.-P.; Ou-Yang, D.-S.; Wang, L.-S.; Huang, S.-L.; Tan, Z.-R.; Zhou, H.-H. The distribution and gender difference of CYP3A activity in Chinese subjects. Br. J. Clin. Pharm. 2003, 55, 264–269. [Google Scholar] [CrossRef]
- Zhou, S.-F. Drugs behave as substrates, inhibitors and inducers of human cytochrome P450 3A4. Curr. Drug Metab. 2008, 9, 310–322. [Google Scholar] [CrossRef]
- Zhou, S.; Chan, S.Y.; Goh, B.C.; Chan, E.; Duan, W.; Huang, M.; McLeod, H.L. Mechanism-Based Inhibition of Cytochrome P450 3A4 by Therapeutic Drugs. Clin. Pharmacokinet. 2005, 44, 279–304. [Google Scholar] [CrossRef]
- Saiz-Rodríguez, M.; Ochoa, D.; Belmonte, C.; Román, M.; Vieira de Lara, D.; Zubiaur, P.; Koller, D.; Mejía, G.; Abad-Santos, F. Polymorphisms in CYP1A2, CYP2C9 and ABCB1 affect agomelatine pharmacokinetics. J. Psychopharmacol. (Oxf.) 2019, 33, 522–531. [Google Scholar] [CrossRef]
- Ensembl Data. CYP3A4 rs4986910. Available online: https://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=7:99760401-99761401;v=rs4986910;vdb=variation;vf=17958388 (accessed on 25 February 2020).
- Ensembl Data. CYP3A4 rs67666821. Available online: http://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=7:99757684-99758683;v=rs67666821;vdb=variation;vf=12903884 (accessed on 25 February 2020).
- Ensembl Data. CYP3A4 rs35599367. Available online: https://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=7:99768193-99769193;v=rs35599367;vdb=variation;vf=20923380 (accessed on 25 February 2020).
- Ensembl Data. CYP3A5 rs776746. Available online: http://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=7:99672416-99673416;v=rs776746;vdb=variation;vf=550116 (accessed on 25 February 2020).
- Ensembl Data. CYP3A5 rs10264272. Available online: https://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=7:99664712-99665712;v=rs10264272;vdb=variation;vf=19096090 (accessed on 25 February 2020).
- Ensembl Data. CYP3A5 rs41303343. Available online: https://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=7:99652271-99653271;v=rs41303343;vdb=variation;vf=21091103 (accessed on 25 February 2020).
- Belmonte, C.; Ochoa, D.; Román, M.; Saiz-Rodríguez, M.; Wojnicz, A.; Gómez-Sánchez, C.I.; Martín-Vílchez, S.; Abad-Santos, F. Influence of CYP2D6, CYP3A4, CYP3A5 and ABCB1 Polymorphisms on Pharmacokinetics and Safety of Aripiprazole in Healthy Volunteers. Basic Clin. Pharmacol. Toxicol. 2018, 122, 596–605. [Google Scholar] [CrossRef]
- Saiz-Rodríguez, M.; Ochoa, D.; Herrador, C.; Belmonte, C.; Román, M.; Alday, E.; Koller, D.; Zubiaur, P.; Mejía, G.; Hernández-Martínez, M.; et al. Polymorphisms associated with fentanyl pharmacokinetics, pharmacodynamics and adverse effects. Basic Clin. Pharmacol. Toxicol. 2019, 124, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Kuip, E.J.M.; Zandvliet, M.L.; Koolen, S.L.W.; Mathijssen, R.H.J.; van der Rijt, C.C.D. A review of factors explaining variability in fentanyl pharmacokinetics; focus on implications for cancer patients. Br. J. Clin. Pharmacol. 2017, 83, 294–313. [Google Scholar] [CrossRef] [PubMed]
- Takashina, Y.; Naito, T.; Mino, Y.; Yagi, T.; Ohnishi, K.; Kawakami, J. Impact of CYP3A5 and ABCB1 gene polymorphisms on fentanyl pharmacokinetics and clinical responses in cancer patients undergoing conversion to a transdermal system. Drug Metab. Pharmacokinet. 2012, 27, 414–421. [Google Scholar]
- Drogari, E.; Ragia, G.; Mollaki, V.; Elens, L.; Van Schaik, R.H.; Manolopoulos, V.G. POR*28 SNP is associated with lipid response to atorvastatin in children and adolescents with familial hypercholesterolemia. Pharmacogenomics 2014, 15, 1963–1972. [Google Scholar] [CrossRef] [PubMed]
- Ragia, G.; Kolovou, V.; Tavridou, A.; Elens, L.; Tselepis, A.D.; Elisaf, M.; Van Schaik, R.H.N.; Kolovou, G.; Manolopoulos, V.G. No effect of CYP3A4 intron 6 C>T polymorphism (CYP3A4*22) on lipid-lowering response to statins in Greek patients with primary hypercholesterolemia. Drug Metab. Personal. Ther. 2015, 30. Available online: https://www.degruyter.com/view/j/dmdi.2015.30.issue-1/dmdi-2014-0021/dmdi-2014-0021.xml (accessed on 25 February 2020).
- Rosales, A.; Alvear, M.; Cuevas, A.; Saavedra, N.; Zambrano, T.; Salazar, L.A. Identification of pharmacogenetic predictors of lipid-lowering response to atorvastatin in Chilean subjects with hypercholesterolemia. Clin. Chim. Acta 2012, 413, 495–501. [Google Scholar] [CrossRef]
- Peng, C.; Ding, Y.; Yi, X.; Shen, Y.; Dong, Z.; Cao, L.; Li, Q.; Ren, H.; He, L.; Zhou, D.; et al. Polymorphisms in CYP450 Genes and the Therapeutic Effect of Atorvastatin on Ischemic Stroke: A Retrospective Cohort Study in Chinese Population. Clin. Ther. 2018, 40, 469–477. [Google Scholar] [CrossRef]
- Liu, H.; Xu, Q.; Huang, W.; Zhao, Q.; Jiang, Z.; Kuang, X.; Li, Z.; Sun, H.; Qiu, X. CYP3A5 and CYP3A7 genetic polymorphisms affect tacrolimus concentration in pediatric patients with nephrotic range proteinuria. Eur. J. Clin. Pharmacol. 2019, 75, 1533–1540. [Google Scholar] [CrossRef]
- Qiu, H.; Zhuang, W.; Wang, X.; Huang, M.; Zhou, Z. Association between genetic polymorphisms and variation of imatinib pharmacokinetics in gastrointestinal stromal tumors. Zhonghua Wei Chang Wai Ke Za Zhi 2017, 20, 1031–1034. [Google Scholar]
- Qian, Y.; Sun, L.-N.; Liu, Y.-J.; Zhang, Q.; Xu, J.-H.; Ma, Z.-Q.; Zhang, X.-H.; Xu, H.; Wang, Y.-Q. Genetic Polymorphisms and Adverse Events on Unbound Imatinib and Its Active Metabolite Concentration in Patients With Gastrointestinal Stromal Tumors. Front. Pharmacol. 2019, 10, 854. [Google Scholar] [CrossRef]
- Ensembl Data. CYP3A4 rs755828176. Available online: https://www.ensembl.org/Homo_sapiens/Variation/Explore?r=6:160139351-160140352;v=rs755828176;vdb=variation;vf=213359709 (accessed on 17 February 2020).
- Belohlavkova, P.; Vrbacky, F.; Voglova, J.; Racil, Z.; Zackova, D.; Hrochova, K.; Malakova, J.; Mayer, J.; Zak, P. The significance of enzyme and transporter polymorphisms for imatinib plasma levels and achieving an optimal response in chronic myeloid leukemia patients. Aoms 2018, 14, 1416–1423. [Google Scholar] [CrossRef]
- Seong, S.J.; Lim, M.; Sohn, S.K.; Moon, J.H.; Oh, S.-J.; Kim, B.S.; Ryoo, H.M.; Chung, J.S.; Joo, Y.D.; Bang, S.M.; et al. Influence of enzyme and transporter polymorphisms on trough imatinib concentration and clinical response in chronic myeloid leukemia patients. Ann. Oncol. 2013, 24, 756–760. [Google Scholar] [CrossRef]
- Petain, A.; Kattygnarath, D.; Azard, J.; Chatelut, E.; Delbaldo, C.; Geoerger, B.; Barrois, M.; Séronie-Vivien, S.; LeCesne, A.; Vassal, G.; et al. Population pharmacokinetics and pharmacogenetics of imatinib in children and adults. Clin. Cancer Res. 2008, 14, 7102–7109. [Google Scholar] [CrossRef]
- Adeagbo, B.A.; Bolaji, O.O.; Olugbade, T.A.; Durosinmi, M.A.; Bolarinwa, R.A.; Masimirembwa, C. Influence of CYP3A5*3 and ABCB1 C3435T on clinical outcomes and trough plasma concentrations of imatinib in Nigerians with chronic myeloid leukaemia. J. Clin. Pharm. Ther. 2016, 41, 546–551. [Google Scholar] [CrossRef]
- Maddin, N.; Husin, A.; Gan, S.H.; Aziz, B.A.; Ankathil, R. Impact of CYP3A4*18 and CYP3A5*3 Polymorphisms on Imatinib Mesylate Response Among Chronic Myeloid Leukemia Patients in Malaysia. Oncol. Ther. 2016, 4, 303–314. [Google Scholar] [CrossRef]
- Huang, Y.; Wen, G.; Lu, Y.; Wen, J.; Ji, Y.; Xing, X.; Li, Y.; Wen, J.; Yuan, H. CYP3A4*1G and CYP3A5*3 genetic polymorphisms alter the antihypertensive efficacy of amlodipine in patients with hypertension following renal transplantation. CP 2017, 55, 109–118. [Google Scholar] [CrossRef]
- Kim, K.; Park, P.; Lee, O.; Choi, S.; Min, B.; Shin, K.; Chun, B.; Shin, J.; Park, J. Effect of CYP3A53 genotype on the pharmacokinetics and pharmacodynamics of amlodipine in healthy Korean subjects. Clin. Pharm. Ther. 2006, 80, 646–656. [Google Scholar] [CrossRef]
- Cabaleiro, T.; López-Rodríguez, R.; Román, M.; Ochoa, D.; Novalbos, J.; Borobia, A.; Carcas, A.; Abad-Santos, F. Pharmacogenetics of quetiapine in healthy volunteers: Association with pharmacokinetics, pharmacodynamics, and adverse effects. Int. Clin. Psychopharmacol. 2015, 30, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Shilbayeh, S.A.R.; Sy, S.K.B.; Melhem, M.; Zmeili, R.; Derendorf, H. Quantitation of the impact of CYP3A5 A6986G polymorphism on quetiapine pharmacokinetics by simulation of target attainment: Clinical Pharmacology in Drug Development. Clin. Pharm. Drug Dev. 2015, 4, 387–394. [Google Scholar] [CrossRef]
- Bakken, G.V.; Molden, E.; Hermann, M. Impact of Genetic Variability in CYP2D6, CYP3A5, and ABCB1 on Serum Concentrations of Quetiapine and N-desalkylquetiapine in Psychiatric Patients. Ther. Drug Monit. 2015, 37, 256–261. [Google Scholar] [CrossRef]
- Noetzli, M.; Guidi, M.; Ebbing, K.; Eyer, S.; Wilhelm, L.; Michon, A.; Thomazic, V.; Stancu, I.; Alnawaqil, A.-M.; Bula, C.; et al. Population pharmacokinetic approach to evaluate the effect of CYP2D6, CYP3A, ABCB1, POR and NR1I2 genotypes on donepezil clearance: Pharmacogenetic study on donepezil. Br. J. Clin. Pharmacol. 2014, 78, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Magliulo, L.; Dahl, M.-L.; Lombardi, G.; Fallarini, S.; Villa, L.M.; Biolcati, A.; Scordo, M.G. Do CYP3A and ABCB1 genotypes influence the plasma concentration and clinical outcome of donepezil treatment? Eur. J. Clin. Pharmacol. 2011, 67, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, J.; San, S.N.; Fujiyoshi, M.; Kawauchi, A.; Chiba, N.; Tagai, R.; Sanbe, R.; Yanaka, S.; Sakaue, H.; Kato, Y.; et al. Effect of CYP3A5*3 genetic variant on the metabolism of direct-acting antivirals in vitro: A different effect on asunaprevir versus daclatasvir and beclabuvir. J. Hum. Genet. 2020, 65, 143–153. [Google Scholar] [CrossRef] [PubMed]



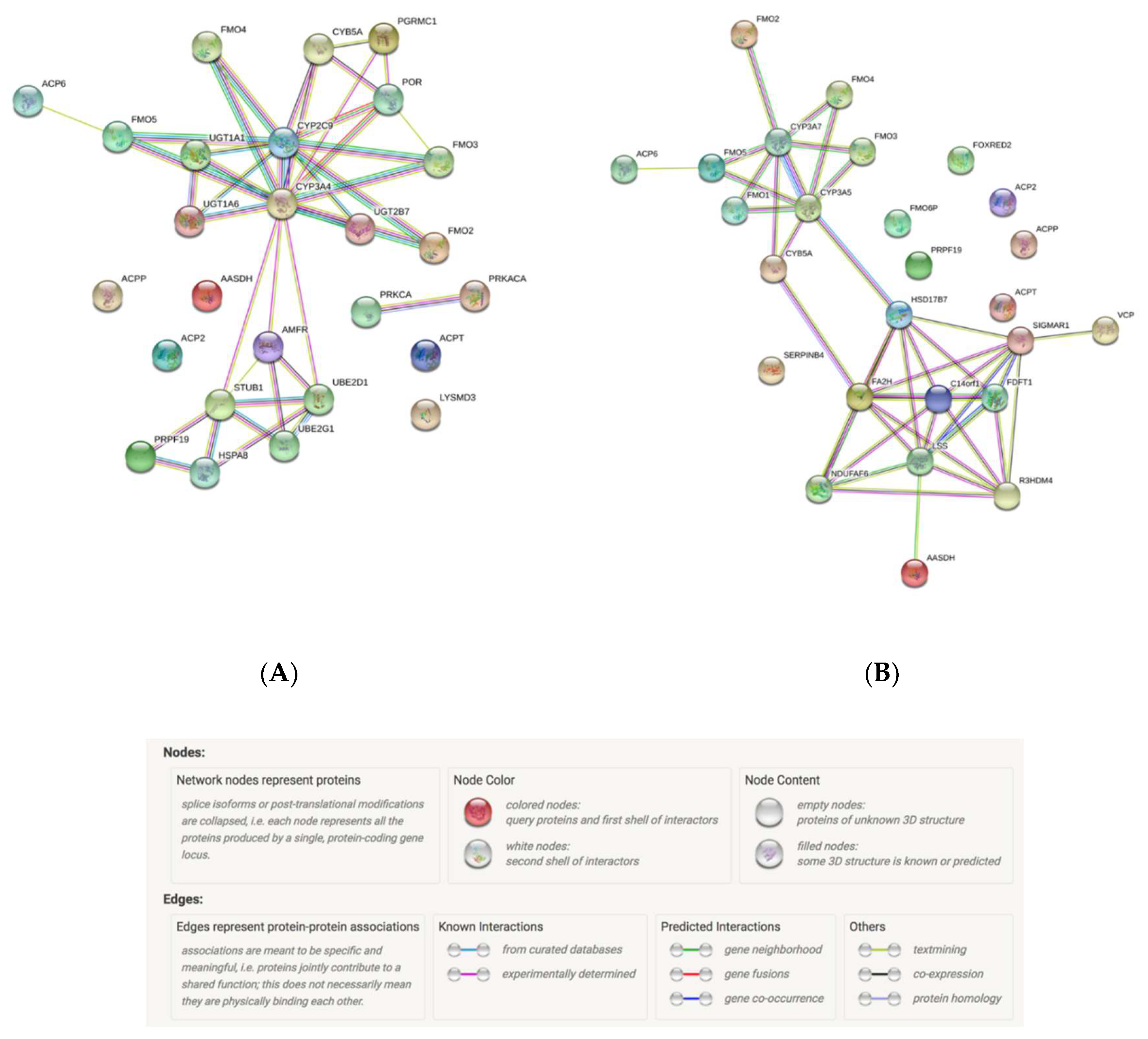{kind=link}
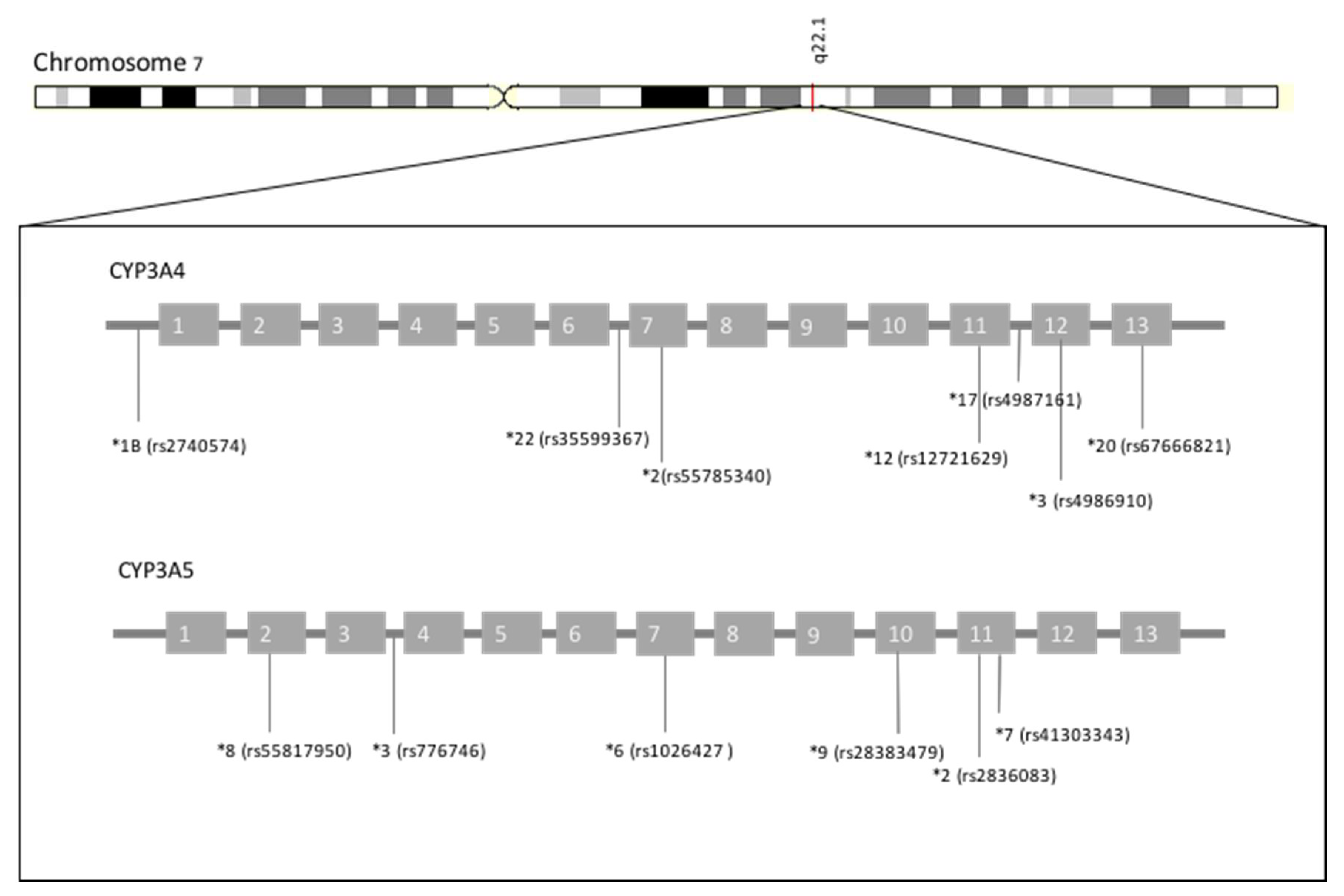{kind=link}
| CYP3A Allele | Reference SNP Identifier | In Vitro Effect | PK Effect in CYP3A Metabolism | Minor Allele Frequencies | |||
|---|---|---|---|---|---|---|---|
| Europeans | Latin-Americans | Africans | East Asians | ||||
| CYP3A4*22 | rs35599367 | ↓ [35,36] | ↓ | 50/1006 (4.97%) | 18/694 (2.59%) | 1/1322 (0.08%) | 0/1008 (0%) |
| CYP3A4*1B | rs2740574 | (= [46], ↓ [47]?, ↑ [48]?) | (↑?) [38], [41,42,49,50,51,52,53,54] | 28/1006 (2.78%) | 73/694 (10.52%) | 1012/1322 (76.55%) | 4/1008 (0.4%) |
| CYP3A4*1G | rs2242480 | (↑?) [59], [60,61,62,63,64,65] | 82/1006 (8.15%) | 273/694 (39.34%) | 1124/1322 (85.02%) | 270/1008 (26.79%) | |
| CYP3A4*3 | rs4986910 | (↓?) [72] | 7/1006 (0.70%) | 5/694 (0.72%) | 1/1322 (0.08%) | 0/1008 (0%) | |
| CYP3A4*20 | rs67666821 | None | ↓ | 26/64600 (0.04%)~ | 22/13290 (0.17%)~ | 3/42021 (0.01%)~ | 0/3134 (0.00%)~ |
| CYP3A5*3 | rs776746 | ↓ | ↓ | 949/1006 (94.33%) | 553/694 (79.68%) | 238/1322 (18.00%) | 719/1008 (71.33%) |
| CYP3A5*6 | rs10264272 | (↓?) [74] | (↓?) [39], [40] | 3/1006 (0.30%) | 16/694 (2.31%) | 204/1322 (15.43%) | 0/1008 (0.00%) |
| CYP3A5*7 | rs41303343 | =/↓ | (↓?) [39], [40] | 0/942 (0.00%)^ | 27/1090 (2.48%)^ | 174/2014 (8.64%)^ | 0/480 (0.00%)^ |
| CYP3A5*8 | rs55817950 | ↓ | 0/ 113648 (0.00%) | 0/ 34590 (0.00%) | 0/ 16216 (0.00%) | 0/ 18391 (0.00%) | |
| CYP3A5*9 | rs28383479 | ↓ | 0/113434(0.00%) | 0/34340(0.00%) | 0/16226(0.00%) | 2/18358 (0.01%) | |
| Gene | Genotype | N | % |
| CYP3A4 | *1/*1 | 233 | 92.8 |
| *1/*20 | 5 | 2.0 | |
| *1/*22 | 11 | 4.4 | |
| *1/*3 | 1 | 0.4 | |
| *3/*22 | 1 | 0.4 | |
| CYP3A5 | *1/*1 | 5 | 2.0 |
| *1/*3 | 45 | 17.9 | |
| *3/*3 | 192 | 76.5 | |
| *3/*6 | 8 | 3.2 | |
| *3/*7 | 1 | 0.4 | |
| Phenotype | N | % | |
| CYP3A | PM | 17 | 6.8 |
| IM | 184 | 73.3 | |
| EM | 50 | 19.9 |
| Gene | Genotype/Phenotype | Pharmacokinetic Parameter | |||||
|---|---|---|---|---|---|---|---|
| Normalized AUC | Normalized Cmax | Normalized T1/2 | Normalized Tmax | Normalized Cl | Normalized Vd | ||
| CYP3A4 | *1/*1 (n = 233) | 0.99 (0.35) | 0.99 (0.42) | 0.99 (0.32) | 1.00 (0.56) | 0.99 (0.38) | 1.00 (0.37) |
| *1/*20 (n = 5) | 1.12 (0.24) | 0.96 (0.33) | 1.18 (0.24) | 0.66 (0.27) | 0.73 (0.18) | 0.90 (0.10) | |
| *1/*22 (n = 11) | 1.06 (0.46) | 0.98 (0.38) | 0.99 (0.18) | 1.00 (0.71) | 0.96 (0.43) | 0.98 (0.43) | |
| *1/*3 + *3/*22 (n = 2) | 1.24 (0.40) | 1.25 (0.10) | 1.07 (0.28) | 0.93 (0.03) | 0.75 (0.39) | 0.83 (0.18) | |
| p-value | 0.523 | 0.738 | 0.566 | 0.482 | 0.338 | 0.874 | |
| CYP3A4 | Wild-type (n = 233) | 0.99 (0.35) | 0.99 (0.42) | 0.99 (0.32) | 1.00 (0.56) | 0.99 (0.37) | 1.00 (0.37) |
| Mutated (n = 18) | 1.10 (0.39) | 1.00 (0.34) | 1.05 (0.20) | 0.90 (0.58) | 0.87 (0.37) | 0.94 (0.34) | |
| p-value | 0.240 | 0.853 | 0.276 | 0.351 | 0.146 | 0.428 | |
| CYP3A5 | *1/*1 (n = 5) | 0.90 (0.30) | 1.13 (0.48) | 0.88 (0.19) | 0.73 (0.26) | 1.20 (0.51) | 1.05 (0.27) |
| *1/*3 (n = 45) | 1.02 (0.32) | 1.02 (0.44) | 1.04 (0.36) | 0.90 (0.52) | 0.94 (0.38) | 0.96 (0.23) | |
| *3/*3 + *3/*6 + *3/*7 (n = 201) | 0.99 (0.37) | 0.99 (0.40) | 0.99 (0.31) | 1.03 (0.57) | 0.98 (0.38) | 1.00 (0.39) | |
| p-value | 0.710 | 0.626 | 0.731 | 0.257 | 0.486 | 0.833 | |
| CYP3A | EM (n = 50) | 1.01 (0.31) | 1.03 (0.44) | 1.02 (0.35) | 0.88 (0.50) | 0.97 (0.39) | 0.97 (0.23) |
| IM (n = 183) | 0.99 (0.37) | 0.99 (0.41) | 0.98 (0.31) | 1.03 (0.57) | 0.99 (0.37) | 1.01 (0.40) | |
| PM (n = 18) | 1.09 (0.39) | 1.00 (0.34) | 1.05 (0.21) | 0.93 (0.59) | 0.87 (0.37) | 0.94 (0.34) | |
| p-value | 0.408 | 0.692 | 0.501 | 0.155 | 0.324 | 0.720 | |
| Sex | Men (n = 149) | 1.00 (0.37) | 1.00 (0.43) | 0.96 (0.30) | 0.99 (0.58) | 0.99 (0.38) | 0.96 (0.32) |
| Women (n = 102) | 0.99 (0.35) | 1.00 (0.39) | 1.05 (0.34) | 1.01 (0.54) | 0.97 (0.37) | 1.06 (0.41) | |
| p-value | 0.765 | 0.832 | 0.030 | 0.426 | 0.889 | 0.057 | |
| Gene | Genotype/Phenotype | Pharmacokinetic Parameter | |||||
|---|---|---|---|---|---|---|---|
| Normalized AUC | Normalized Cmax | Normalized T1/2 | Normalized Tmax | Normalized Cl | Normalized Vd | ||
| CYP3A4 | *1/*1 (n = 150) | 0.98 (0.41) | 1.00 (0.49) | 1.00 (0.31) | 1.00 (0.56) | 1.00 (0.40) | 1.00 (0.41) |
| *1/*20 (n = 4) | 1.08 (0.26) | 0.91 (0.36) | 1.13 (0.25) | 0.74 (0.24) | 0.78 (0.16) | 0.89 (0.12) | |
| *1/*22 (n = 9) | 1.14 (0.46) | 1.02 (0.39) | 1.01 (0.19) | 1.01 (0.80) | 0.85 (0.39) | 0.91 (0.45) | |
| *3/*22 (n = 1) | 1.53 | 1.32 | 1.27 | 0.96 | 0.47 | 0.71 | |
| p-value | 0.317 | 0.796 | 0.626 | 0.822 | 0.194 | 0.673 | |
| CYP3A4 | Wild-type (n = 150) | 0.98 (0.41) | 1.00 (0.49) | 0.99 (0.31) | 1.00 (0.56) | 1.00 (0.40) | 1.00 (0.41) |
| Mutated (n = 14) | 1.15 (0.40) | 1.01 (0.37) | 1.06 (0.20) | 0.93 (0.64) | 0.81 (0.33) | 0.89 (0.36) | |
| p-value | 0.099 | 0.723 | 0.274 | 0.459 | 0.069 | 0.264 | |
| CYP3A5 | *1/*1 (n = 2) | 0.96 (0.49) | 1.24 (0.72) | 0.83 (0.19) | 0.75 (0.48) | 1.25 (0.81) | 0.99 (0.47) |
| *1/*3 (n = 23) | 1.02 (0.42) | 1.01 (0.59) | 1.03 (0.40) | 0.94 (0.63) | 0.97 (0.48) | 0.92 (0.28) | |
| *3/*3 + *3/*6 + *3/*7 (n = 139) | 1.00 (0.41) | 0.99 (0.45) | 1.00 (0.29) | 1.01 (0.56) | 0.98 (0.38) | 1.01 (0.42) | |
| p-value | 0.987 | 0.760 | 0.834 | 0.538 | 0.731 | 0.818 | |
| CYP3A | EM (n = 25) | 1.01 (0.42) | 1.03 (0.59) | 1.01 (0.39) | 0.92 (0.62) | 0.99 (0.49) | 0.93 (0.29) |
| IM (n = 126) | 0.98 (0.41) | 1.00 (0.47) | 0.99 (0.30) | 1.01 (0.55) | 0.99 (0.38) | 1.02 (0.43) | |
| PM (n = 13) | 1.14 (0.41) | 0.94 (0.28) | 1.06 (0.21) | 0.97 (0.65) | 0.82 (0.34) | 0.91 (0.37) | |
| p-value | 0.342 | 0.988 | 0.566 | 0.560 | 0.280 | 0.526 | |
| Sex | Men (n = 95) | 0.99 (0.43) | 0.98 (0.50) | 0.96 (0.27) | 0.96 (0.57) | 1.00 (0.41) | 0.96 (0.34) |
| Women (n = 69) | 1.01 (0.39) | 1.03 (0.44) | 1.05 (0.34) | 1.04 (0.56) | 0.96 (0.38) | 1.05 (0.48) | |
| p-value | 0.747 | 0.278 | 0.093 | 0.209 | 0.618 | 0.304 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saiz-Rodríguez, M.; Almenara, S.; Navares-Gómez, M.; Ochoa, D.; Román, M.; Zubiaur, P.; Koller, D.; Santos, M.; Mejía, G.; Borobia, A.M.; et al. Effect of the Most Relevant CYP3A4 and CYP3A5 Polymorphisms on the Pharmacokinetic Parameters of 10 CYP3A Substrates. Biomedicines 2020, 8, 94. https://doi.org/10.3390/biomedicines8040094
Saiz-Rodríguez M, Almenara S, Navares-Gómez M, Ochoa D, Román M, Zubiaur P, Koller D, Santos M, Mejía G, Borobia AM, et al. Effect of the Most Relevant CYP3A4 and CYP3A5 Polymorphisms on the Pharmacokinetic Parameters of 10 CYP3A Substrates. Biomedicines. 2020; 8(4):94. https://doi.org/10.3390/biomedicines8040094
Chicago/Turabian StyleSaiz-Rodríguez, Miriam, Susana Almenara, Marcos Navares-Gómez, Dolores Ochoa, Manuel Román, Pablo Zubiaur, Dora Koller, María Santos, Gina Mejía, Alberto M. Borobia, and et al. 2020. "Effect of the Most Relevant CYP3A4 and CYP3A5 Polymorphisms on the Pharmacokinetic Parameters of 10 CYP3A Substrates" Biomedicines 8, no. 4: 94. https://doi.org/10.3390/biomedicines8040094
APA StyleSaiz-Rodríguez, M., Almenara, S., Navares-Gómez, M., Ochoa, D., Román, M., Zubiaur, P., Koller, D., Santos, M., Mejía, G., Borobia, A. M., Rodríguez-Antona, C., & Abad-Santos, F. (2020). Effect of the Most Relevant CYP3A4 and CYP3A5 Polymorphisms on the Pharmacokinetic Parameters of 10 CYP3A Substrates. Biomedicines, 8(4), 94. https://doi.org/10.3390/biomedicines8040094