Apple Supplementation Improves Hemodynamic Parameter and Attenuates Atherosclerosis in High-Fat Diet-Fed Apolipoprotein E-Knockout Mice

 , , ,
, , ,
Abstract
1. Introduction
2. Experimental Section
2.1. Products
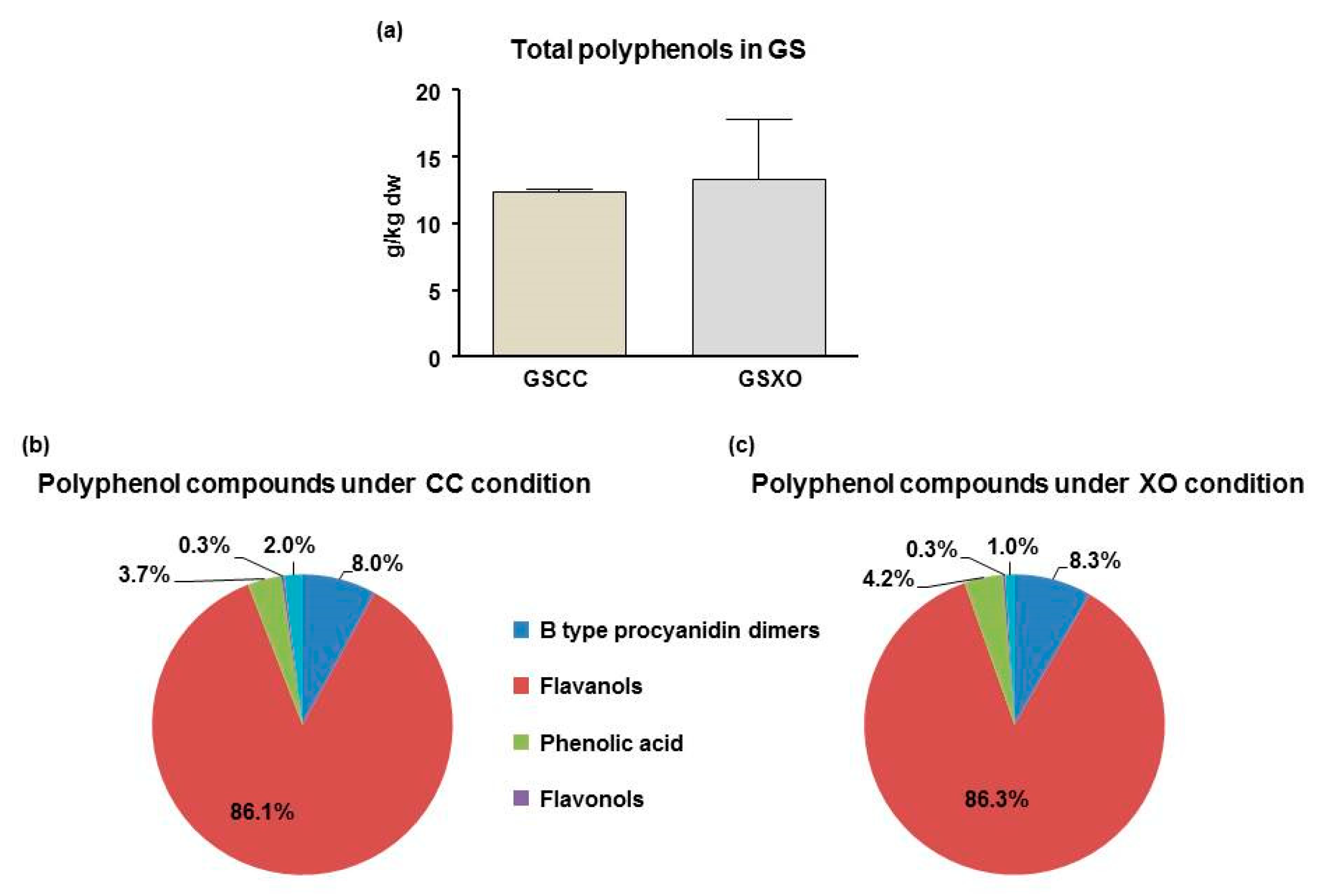
2.2. Determination of Polyphenol Content in GS
2.3. Ethics Statement
2.4. Animals
2.5. Blood Pressure and Heart Rate Measurements
2.6. Echocardiography
2.7. Biochemical Parameters
2.8. Hepatic Secretion of very Low Density Lipoprotein (VLDL) TG
2.9. RNA Extraction and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.10. Atherosclerotic Lesions Analysis
2.11. Atherosclerotic Lesions in Aortic Root
2.12. Liver Histology
2.13. Statistical Analysis
3. Results
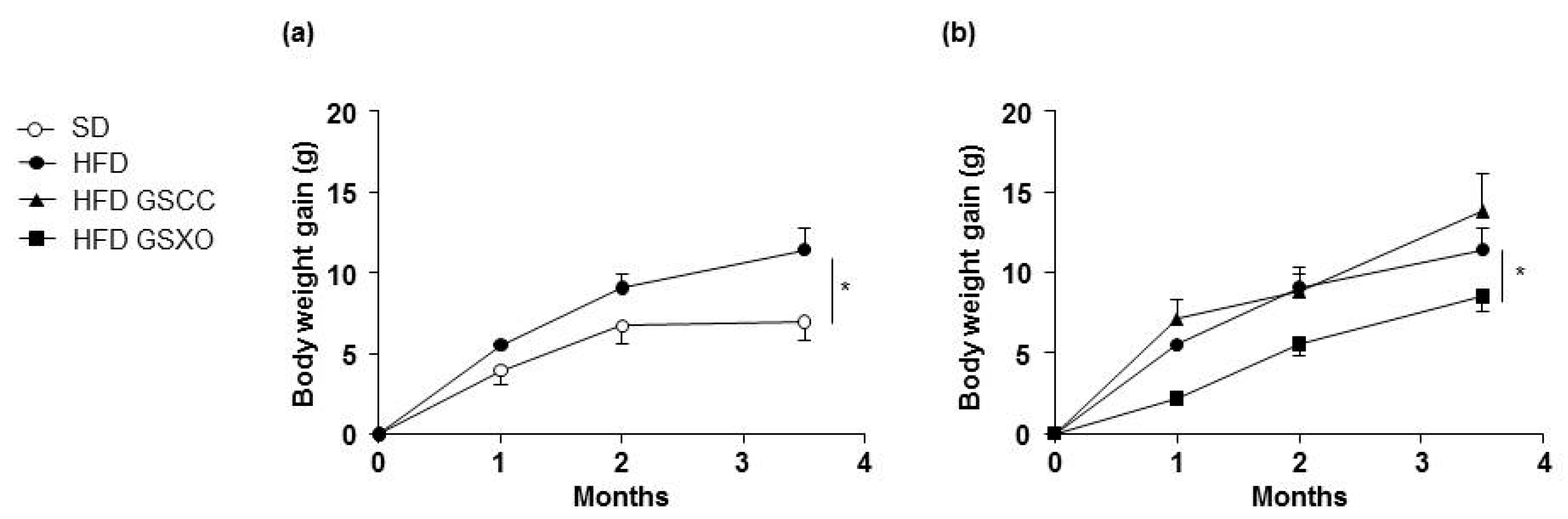
3.1. Effects of Apple Supplementation on Body Weight Gain
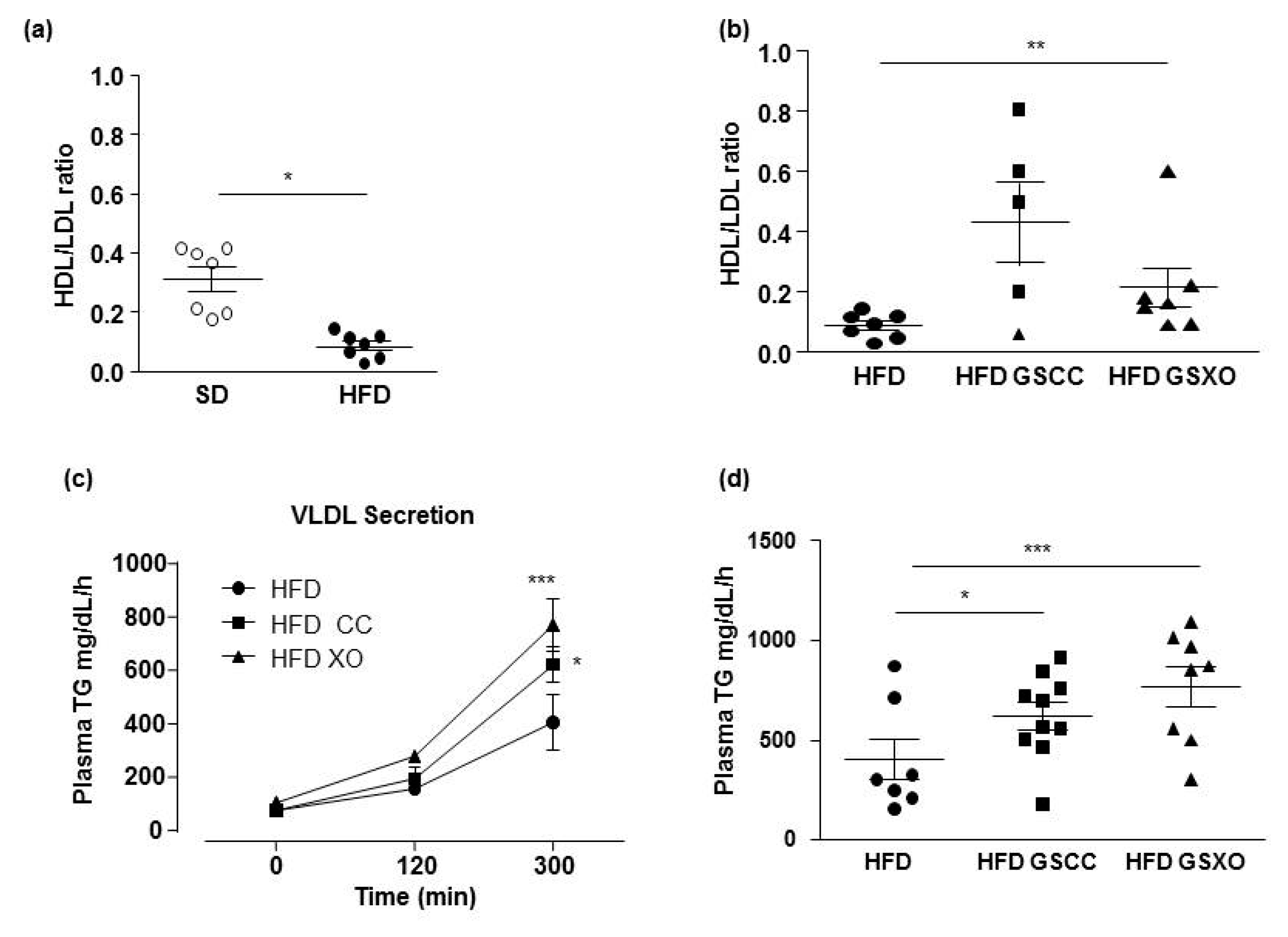
3.2. Effects of Apple Supplementation on Glucose and Lipid Parameters
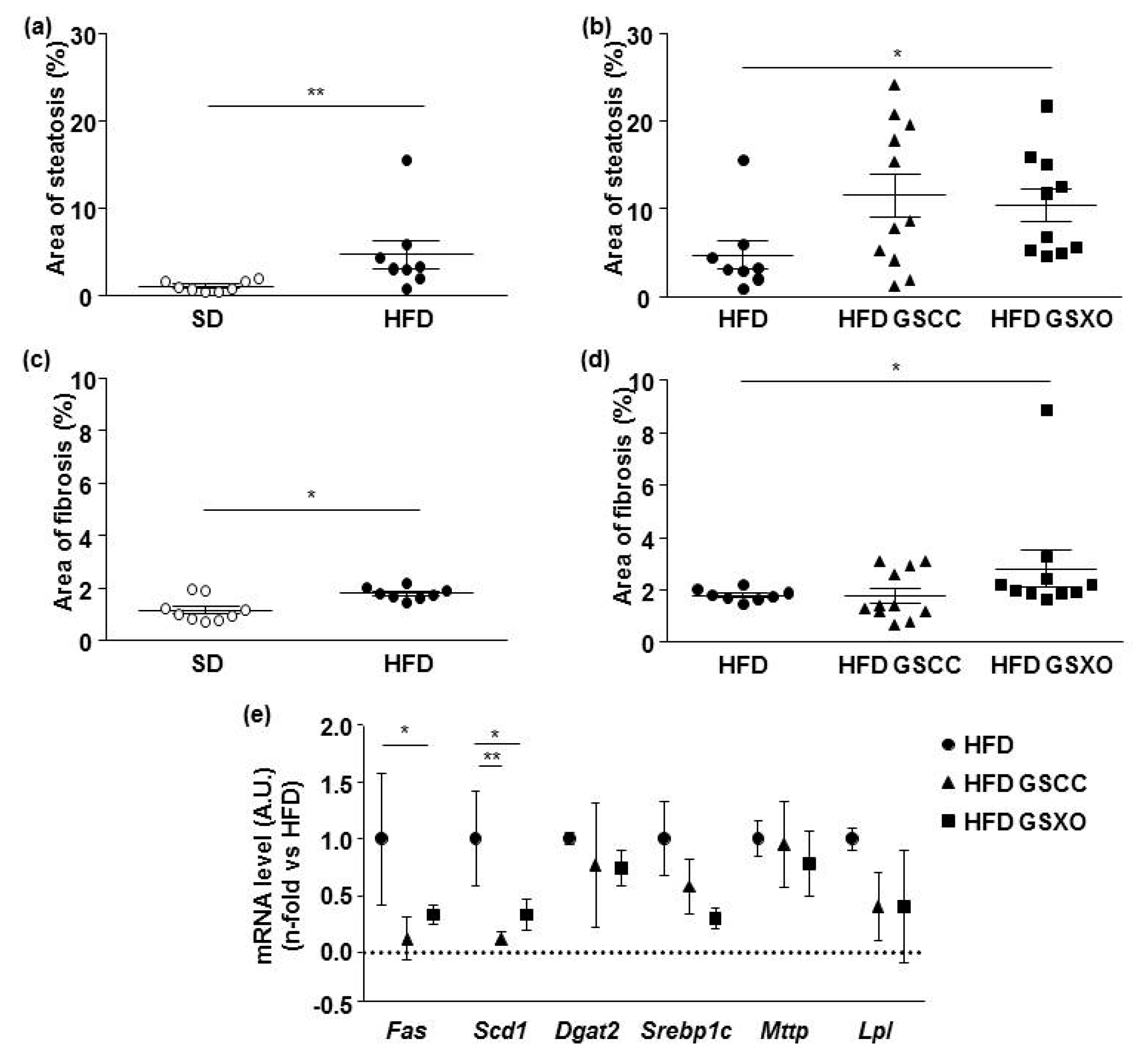
3.3. Effects of Apple Supplementation on Liver
3.4. Effects of Apple on Heart Function
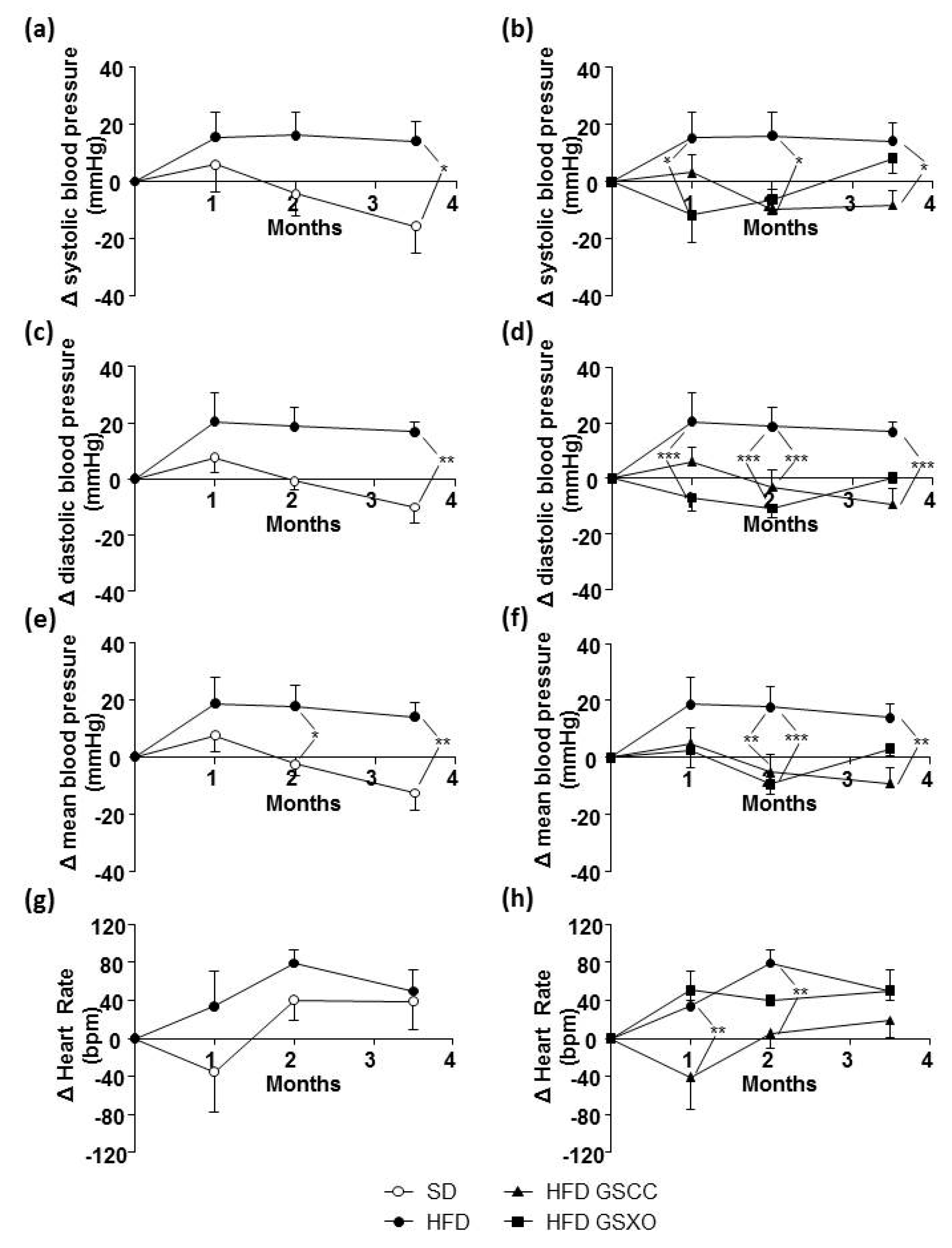
3.5. Effects of Apple Supplementation on Blood Pressure and Heart Rate
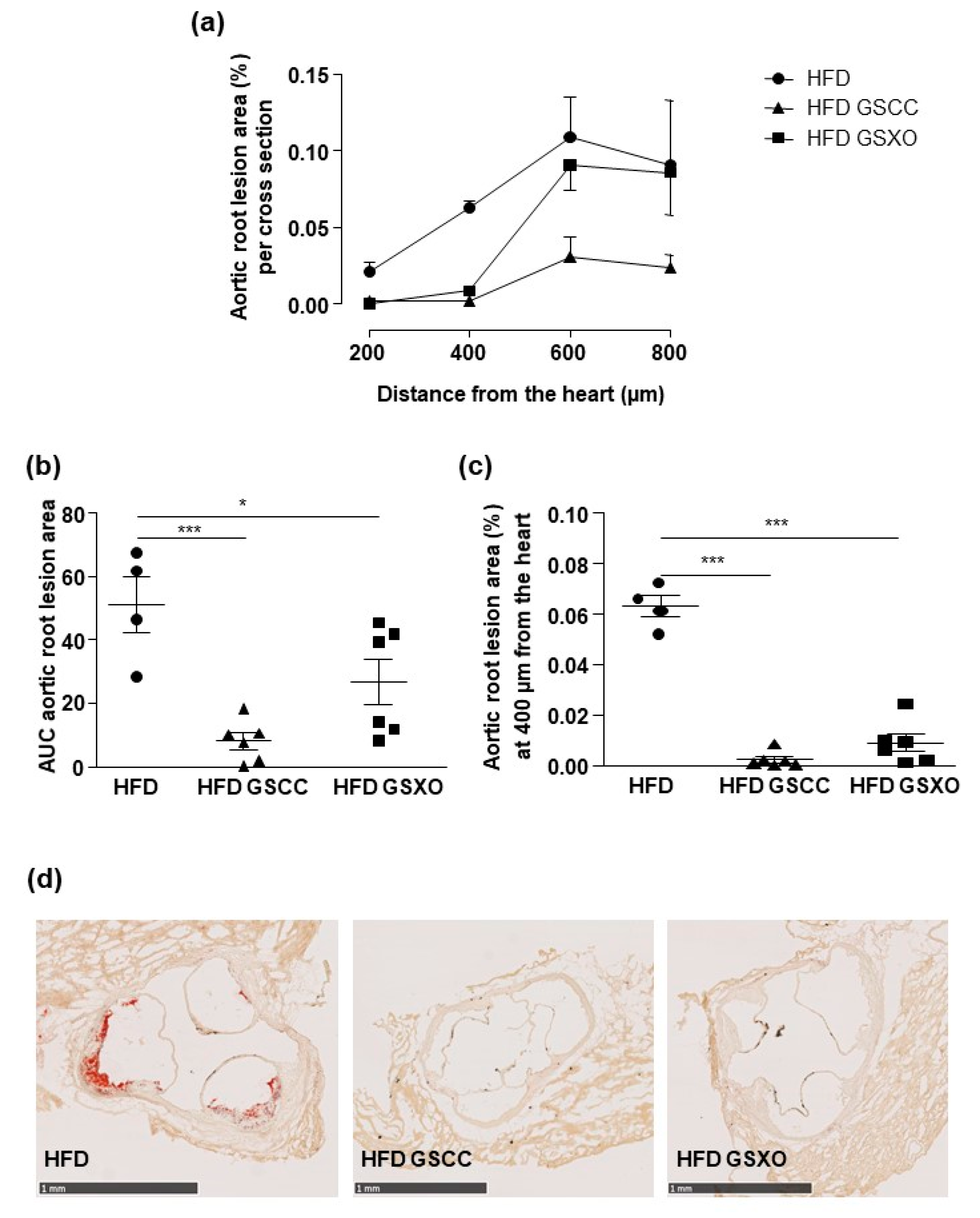
3.6. Effects of Apple Supplementation on Atherosclerotic Lesion
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| ApoE-/- | Apolipoprotein E-deficient mice |
| CC | Classic cold condition |
| DGAT2 | Diacylglycerol O-acyltransferase 2 |
| FAS | Fatty acid synthase |
| GS | Granny Smith |
| GSCC | Granny Smith variety after storage in classic cold condition |
| GSXO | Granny Smith variety after extreme ultra-low oxygen condition |
| HDL | High-density lipoprotein |
| HFD | High fat diet |
| HOMA-IR | Homeostasis model for insulin resistance |
| LDL | Low density lipoprotein |
| LPL | Lipoprotein lipase |
| LVEDD | Left ventricular end-diastolic dimension |
| LVEDV | Left ventricular end-diastolic volume |
| LVESD | Left ventricular end-systolic diameter |
| LVESV | Left ventricular end-systolic volume |
| MTTP | Microsomal triglyceride transfer protein |
| SCD1 | Stearoyl-coenzyme A desaturase-1 |
| SD | Standard diet |
| SEM | Standard error of the mean |
| SREBP1C | Sterol regulatory element-binding transcription factor 1 |
| TG | Triglycerides |
| XO | Extreme ultra-low oxygen condition |
Appendix A

References
- Koutsos, A.; Tuohy Kieran, M.; Lovegrove, J.A. Apples and cardiovascular health—is the gut microbiota a core consideration? Nutrients 2015, 7, 3959–3998. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, C.P.; Yang, X.; Croft, K.D.; Considine, M.J.; Ward, N.C.; Rich, L.; Puddey, I.B.; Swinny, E.; Mubarak, A.; Hodgson, J.M. Flavonoid-rich apples and nitrate-rich spinach augment nitric oxide status and improve endothelial function in healthy men and women: A randomized controlled trial. Free Radic. Biol. Med. 2012, 52, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Nagasako-Akazome, Y.; Kanda, T.; Ohtake, Y.; Shimasaki, H.; Kobayashi, T. Apple polyphenols influence cholesterol metabolism in healthy subjects with relatively high body mass index. J. Oleo. Sci. 2007, 56, 417–428. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.C.; Sichieri, R.; Moura, A.S. Weight loss associated with a daily intake of three apples or three pears among overweight women. Nutrition 2003, 19, 253–256. [Google Scholar] [CrossRef]
- Sanchez, D.; Quinones, M.; Moulay, L.; Muguerza, B.; Miguel, M.; Aleixandre, A. Soluble fiber-enriched diets improve inflammation and oxidative stress biomarkers in zucker fatty rats. Pharmacol. Res. 2011, 64, 31–35. [Google Scholar] [CrossRef]
- Setorki, M.; Asgary, S.; Eidi, A.; Rohani, A.H.; Esmaeil, N. Effects of apple juice on risk factors of lipid profile, inflammation and coagulation, endothelial markers and atherosclerotic lesions in high cholesterolemic rabbits. Lipids Health Dis. 2009, 8, 39. [Google Scholar] [CrossRef]
- Song, Y.; Manson, J.E.; Buring, J.E.; Sesso, H.D.; Liu, S. Associations of dietary flavonoids with risk of type 2 diabetes, and markers of insulin resistance and systemic inflammation in women: A prospective study and cross-sectional analysis. J. Am. Coll Nutr. 2005, 24, 376–384. [Google Scholar] [CrossRef]
- Ravn-Haren, G.; Dragsted, L.O.; Buch-Andersen, T.; Jensen, E.N.; Jensen, R.I.; Németh-Balogh, M.; Paulovicsová, B.; Bergström, A.; Wilcks, A.; Licht, T.R.; et al. Intake of whole apples or clear apple juice has contrasting effects on plasma lipids in healthy volunteers. Eur. J. Nutr. 2013, 52, 1875–1889. [Google Scholar] [CrossRef]
- Chai, S.C.; Hooshmand, S.; Saadat, R.L.; Payton, M.E.; Brummel-Smith, K.; Arjmandi, B.H. Daily apple versus dried plum: Impact on cardiovascular disease risk factors in postmenopausal women. J. Acad. Nutr. Diet 2012, 112, 1158–1168. [Google Scholar] [CrossRef]
- Soleti, R.; Hilairet, G.; Mallegol, P.; Dourguia, C.; Frifra, M.; Guillou, M.C.; Gacel, A.; Guyot, S.; Pignon, P.; Basset, L.; et al. Screening of ordinary commercial varieties of apple fruits under different storage conditions for their potential vascular and metabolic protective properties. Food Funct. 2018, 9, 5855–5867. [Google Scholar] [CrossRef]
- Boord, J.B.; Maeda, K.; Makowski, L.; Babaev, V.R.; Fazio, S.; Linton, M.F.; Hotamisligil, G.S. Adipocyte fatty acid-binding protein, aP2, alters late atherosclerotic lesion formation in severe hypercholesterolemia. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1686–1691. [Google Scholar] [CrossRef] [PubMed]
- Burger-Kentischer, A.; Göbel, H.; Kleemann, R.; Zernecke, A.; Bucala, R.; Leng, L.; Finkelmeier, D.; Geiger, G.; Schaefer, H.E.; Schober, A.; et al. Reduction of the aortic inflammatory response in spontaneous atherosclerosis by blockade of macrophage migration inhibitory factor (MIF). Atherosclerosis 2006, 184, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Nitiéma, M.; Soleti, R.; Koffi, C.; Belemnaba, L.; Mallegol, P.; Ouédraogo, N.; Kini, F.B.; Ouédraogo, S.; Guissou, I.P.; Andriantsitohaina, R. Ethyl Acetate Fraction of Lannea microcarpa Engl. and K. Krause (Anacardiaceae) Trunk Barks Corrects Angiotensin II-Induced Hypertension and Endothelial Dysfunction in Mice. Oxid Med. Cell Longev. 2019, 2019, 9464608. [Google Scholar] [CrossRef] [PubMed]
- Schotz, M.C.; Scanu, A.; Page, I.H. Effect of triton on lipoprotein lipase of rat plasma. Am. J. Physiol. 1957, 188, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Siri, P.; Candela, N.; Zhang, Y.L.; Ko, C.; Eusufzai, S.; Ginsberg, H.N.; Huang, L.S. Post-transcriptional stimulation of the assembly and secretion of triglyceride-rich apolipoprotein B lipoproteins in a mouse with selective deficiency of brown adipose tissue, obesity, and insulin resistance. J. Biol. Chem. 2001, 276, 46064–46072. [Google Scholar] [CrossRef] [PubMed]
- Trenteseaux, C.; Gaston, A.T.; Aguesse, A.; Poupeau, G.; de Coppet, P.; Andriantsitohaina, R.; Laschet, J.; Amarger, V.; Krempf, M.; Nobecourt-Dupuy, E.; et al. Perinatal Hypercholesterolemia Exacerbates Atherosclerosis Lesions in Offspring by Altering Metabolism of Trimethylamine-N-Oxide and Bile Acids. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2053–2063. [Google Scholar] [CrossRef]
- Boursier, J.; de Ledinghen, V.; Sturm, N.; Amrani, L.; Bacq, Y.; Sandrini, J.; Le Bail, B.; Chaigneau, J.; Zarski, J.P.; Gallois, Y.; et al. Precise evaluation of liver histology by computerized morphometry shows that steatosis influences liver stiffness measured by transient elastography in chronic hepatitis C. J. Gastroenterol. 2014, 49, 527–537. [Google Scholar] [CrossRef]
- Auclair, S.; Mathieu, S.; Elyett, G.; Christine, M.; Andrzej, M.; Dragan, M.; Augustin, S. Apple Polyphenols and Fibers Attenuate Atherosclerosis in Apolipoprotein E-Deficient Mice. J. Agric. Food Chem. 2008, 56, 5558–5563. [Google Scholar] [CrossRef]
- Caro-Gómez, E.; Sierra, J.A.; Escobar, J.S.; Álvarez-Quintero, R.; Naranjo, M.; Medina, S.; Velásquez-Mejía, E.P.; Tabares-Guevara, J.H.; Jaramillo, J.C.; León-Varela, Y.M.; et al. Green Coffee Extract Improves Cardiometabolic Parameters and Modulates Gut Microbiota in High-Fat-Diet-Fed ApoE(-/-) Mice. Nutrients 2019, 11, 497. [Google Scholar] [CrossRef]
- Zheng, F.; Cai, Y. Concurrent exercise improves insulin resistance and nonalcoholic fatty liver disease by upregulating PPAR-γ and genes involved in the beta-oxidation of fatty acids in ApoE-KO mice fed a high-fat diet. Lipids Health Dis. 2019, 18, 6. [Google Scholar] [CrossRef]
- Plump, A.S.; Smith, J.D.; Hayek, T.; Aalto-Setälä, K.; Walsh, A.; Verstuyft, J.G.; Rubin, E.M.; Breslow, J.L. Severe hypercholesterolemia and atherosclerosis in apolipoprotein E-deficient mice created by homologous recombination in ES cells. Cell 1992, 71, 343–353. [Google Scholar] [CrossRef]
- Silvestre-Roig, C.; Menno, P.; de Winther, C.W.; Weber, M.J.; Daemen, E.L.; Oliver, S. Atherosclerotic plaque destabilization: Mechanisms, models, and therapeutic strategies. Circ. Res. 2014, 114, 214–226. [Google Scholar] [CrossRef]
- Moghadasian, M.H. Pathophysiology of apolipoprotein E deficiency in mice: Relevance to apo E-related disorders in humans. FASEB J. 2001, 15, 2623–2630. [Google Scholar] [CrossRef] [PubMed]
- Lapuente, M.; Estruch, R.; Shahbaz, M.; Casas, R. Relation of fruits and vegetables with major cardiometabolic risk factors, markers of oxidation, and inflammation. Nutrients 2019, 11, 2381. [Google Scholar] [CrossRef] [PubMed]
- Golding, J.B.; McGlasson, W.B.; Wyllie, S.G.; Leach, D.N. Fate of apple peel phenolics during cool storage. J. Agric. Food Chem. 2001, 49, 2283–2289. [Google Scholar] [CrossRef]
- Awad, M.A.; de Jaeger, A. Influences of air and controlled atmosphere storage on the concentration of potentially healthful phenolics in apples and other fruits. Postharvest Biol. Technol. 2003, 7, 53–58. [Google Scholar] [CrossRef]
- Veberic, R.; Schmitzer, V.M.; Petkovsek, M.; Stampar, F. Impact of shelf life on content of primary and secondary metabolites in apple (Malus domestica Borkh.). Food Sci. 2010, 75, S461–S468. [Google Scholar] [CrossRef]
- Omole, J.O.; Ighodaro, O.M. Comparative studies of the effects of egg yolk, oats, apple, and wheat bran on serum lipid profile of wistar rats. ISRN Nutr. 2012, 2013, 730479. [Google Scholar] [CrossRef]
- Samout, N.; Bouzenna, H.; Dhibi, S.; Ncib, S.; ElFeki, A.; Hfaiedh, N. Therapeutic effect of apple pectin in obese rats. Biomed. Pharmacother. 2016, 83, 1233–1238. [Google Scholar] [CrossRef]
- Shin, S.K.; Cho, S.J.; Jung, U.J.; Ryu, R.; Choi, M.S. Phlorizin Supplementation Attenuates Obesity, Inflammation, and Hyperglycemia in Diet-Induced Obese Mice Fed a High-Fat Diet. Nutrients 2016, 16, 92. [Google Scholar] [CrossRef]
- Dobrzyn, A.; Ntambi, J.M. The role of stearoyl-CoA desaturase in the control of metabolism. PLEFA 2005, 73, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Magkos, F.; Mohammed, B.S.; Pietka, T.; Abumrad, N.A.; Patterson, B.W.; Okunade, A.; Klein, S. Intrahepatic fat, not visceral fat, is linked with metabolic complications of obesity. Proc. Natl. Acad. Sci. USA 2009, 106, 15430–15435. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Häring, H.U. The metabolically benign and malignant fatty liver. Diabetes 2011, 60, 2011–2017. [Google Scholar] [CrossRef] [PubMed]
- Arconzo, M.; Piccinin, E.; Moschetta, A. Increased risk of acute liver failure by pain killer drugs in NAFLD: Focus on nuclear receptors and their coactivators. Dig. Liver Dis. 2020, 13. [Google Scholar] [CrossRef]
- Wang, L.; Manson, J.E.; Gaziano, J.M.; Buring, J.E.; Sesso, H.D. Fruit and vegetable intake and the risk of hypertension in middle-aged and older women. Am. J. Hypertens 2012, 25, 180–189. [Google Scholar] [CrossRef]
- Perez-Vizcaino, F.; Duarte, J.; Andriantsitohaina, R. Endothelial function and cardiovascular disease: Effects of quercetin and wine polyphenols. Free Radic. Res. 2006, 40, 1054–1065. [Google Scholar] [CrossRef]
- Larson, A.J.; Symons, J.D.; Jalili, T. Therapeutic potential of quercetin to decrease blood pressure: Review of efficacy and mechanisms. Adv. Nutr. 2012, 3, 39–46. [Google Scholar] [CrossRef]
- Gonzalez, J.; Donoso, W.; Sandoval, N.; Reyes, M.; Gonzalez, P.; Gajardo, M.; Morales, E.; Neira, A.; Razmilic, I.; Yuri, J.A.; et al. Apple Peel Supplemented Diet Reduces Parameters of Metabolic Syndrome and Atherogenic Progression in ApoE-/- Mice. ECAM 2015, 2015, 918384. [Google Scholar] [CrossRef]
- Batiha, G.E.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The pharmacological activity, biochemical properties, and pharmacokinetics of the major natural polyphenolic flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef]
- Wang, L.; Fumoto, T.; Masumoto, S.; Shoji, T.; Miura, T.; Naraoka, M.; Matsuda, N.; Imaizumi, T.; Ohkuma, H. Regression of atherosclerosis with apple procyanidins by activating the ATP-binding cassette subfamily A member 1 in a rabbit model. Atherosclerosis 2017, 258, 56–64. [Google Scholar] [CrossRef]
- Xu, Z.R.; Li, J.Y.; Dong, X.W.; Tan, Z.J.; Wu, W.Z.; Xie, Q.M.; Yang, Y.M. Apple polyphenols decrease atherosclerosis and hepatic steatosis in apoe-/- mice through the ROS/MAPK/NF-ΚB pathway. Nutrients 2015, 7, 7085–7105. [Google Scholar] [CrossRef]
- Wu, C.; Luan, H.; Zhang, X.; Wang, S.; Zhang, X.; Sun, X.; Guo, P. Chlorogenic acid protects against atherosclerosis in ApoE-/- mice and promotes cholesterol efflux from RAW264.7 macrophages. PLoS ONE 2014, 9, e95452. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | Standard Diet | High Fat Diet | High Fat Diet+Granny Smith (GS) |
|---|---|---|---|
| Composition, g/kg | |||
| GS CC and XO | absent- | absent | 3.33 |
| Sucrose | 207 | 340 | 340 |
| Dairy butter | 50 | 200 | 200 |
| Casein | 200 | 180.5 | 180.5 |
| Pregelatinized cornstarch | 400 | 145 | 145 |
| Premixture of minerals | 70 | 70 | 70 |
| Crude cellulose | absent | 50 | 46.77 |
| Premixture of vitamins | 10 | 10 | 10 |
| DL-methionine | 3 | 3 | 3 |
| Cholesterol | absent | 1.43 | 1.43 |
| Energy, % | |||
| Protein | 19.3 | 17.7 | 17.7 |
| Fat | 8.4 | 41.7 | 41.7 |
| Carbohydrate | 72.4 | 40.6 | 40.6 |
| SD | HFD | HFD GSCC | HFD GSXO | |
|---|---|---|---|---|
| Glucose (g/L) | 1.62 ± 0.29 | 1.81 ± 0.34 | 1.21 ± 0.15 | 0.92 ± 0.15 |
| Insulin (g/L) | 0.79 ± 0.09 | 0.58 ± 0.03 | 0.89 ± 0.19 | 1.19 ± 0.17 ## |
| HOMA-IR | 0.07 ± 0.02 | 0.05 ± 0.01 | 0.05 ± 0.01 | 0.04 ± 0.01 |
| TG (g/L) | 0.89 ± 0.22 | 1.34 ± 0.17 * | 0.86 ± 0.10 # | 0.56 ± 0.14 ## |
| Cholesterol (g/L) | 7.22 ± 1.54 | 7.70 ± 2.25 | 8.43 ± 1.22 | 6.02 ± 1.00 |
| SD | HFD | HFD GSCC | HFD GSXO | |
|---|---|---|---|---|
| LVESD (mm) | 2.3 ± 0.1 | 2.3 ± 0.1 | 2.6 ± 0.1 | 2.6 ± 0.1 |
| LVEDD (mm) | 3.4 ± 0.1 | 3.6 ± 0.1 | 3.9 ± 0.1 | 3.7 ± 0.1 |
| LVESV (mL) | 19.7 ± 2.2 | 19.4 ± 2.2 | 25.6 ± 4.1 | 26.4 ± 2.7 |
| LVEDV (mL) | 60.6 ± 4.8 | 55.3 ± 2.4 | 68 ± 5 | 60.5 ± 4.2 |
| Stroke volume (mL) | 40.9 ± 3.6 | 35.9 ± 2.4 | 42.3 ± 3.4 | 34.1 ± 2 |
| Ejection fraction (%) | 67.2 ± 3.0 | 65.1 ± 3.6 | 62.9 ± 3.7 | 57 ± 2.3 |
| Shortening fraction (%) | 37.1 ± 2.2 | 35.5 ± 2.7 | 34 ± 2.6 | 29.5 ± 1.6 |
| Cardiac output (mL/min) | 21.3 ± 4.1 | 17.4 ± 2.5 | 27.7 ± 3.6 ## | 23 ± 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soleti, R.; Trenteseaux, C.; Fizanne, L.; Coué, M.; Hilairet, G.; Kasbi-Chadli, F.; Mallegol, P.; Chaigneau, J.; Boursier, J.; Krempf, M.; et al. Apple Supplementation Improves Hemodynamic Parameter and Attenuates Atherosclerosis in High-Fat Diet-Fed Apolipoprotein E-Knockout Mice. Biomedicines 2020, 8, 495. https://doi.org/10.3390/biomedicines8110495
Soleti R, Trenteseaux C, Fizanne L, Coué M, Hilairet G, Kasbi-Chadli F, Mallegol P, Chaigneau J, Boursier J, Krempf M, et al. Apple Supplementation Improves Hemodynamic Parameter and Attenuates Atherosclerosis in High-Fat Diet-Fed Apolipoprotein E-Knockout Mice. Biomedicines. 2020; 8(11):495. https://doi.org/10.3390/biomedicines8110495
Chicago/Turabian StyleSoleti, Raffaella, Charlotte Trenteseaux, Lionel Fizanne, Marine Coué, Gregory Hilairet, Fatima Kasbi-Chadli, Patricia Mallegol, Julien Chaigneau, Jerome Boursier, Michel Krempf, and et al. 2020. "Apple Supplementation Improves Hemodynamic Parameter and Attenuates Atherosclerosis in High-Fat Diet-Fed Apolipoprotein E-Knockout Mice" Biomedicines 8, no. 11: 495. https://doi.org/10.3390/biomedicines8110495
APA StyleSoleti, R., Trenteseaux, C., Fizanne, L., Coué, M., Hilairet, G., Kasbi-Chadli, F., Mallegol, P., Chaigneau, J., Boursier, J., Krempf, M., Orsel, M., Ouguerram, K., & Andriantsitohaina, R. (2020). Apple Supplementation Improves Hemodynamic Parameter and Attenuates Atherosclerosis in High-Fat Diet-Fed Apolipoprotein E-Knockout Mice. Biomedicines, 8(11), 495. https://doi.org/10.3390/biomedicines8110495