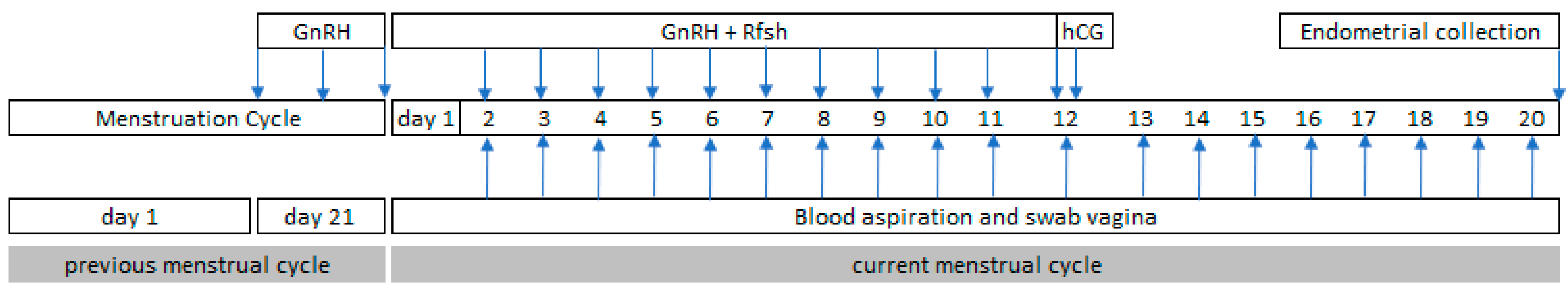
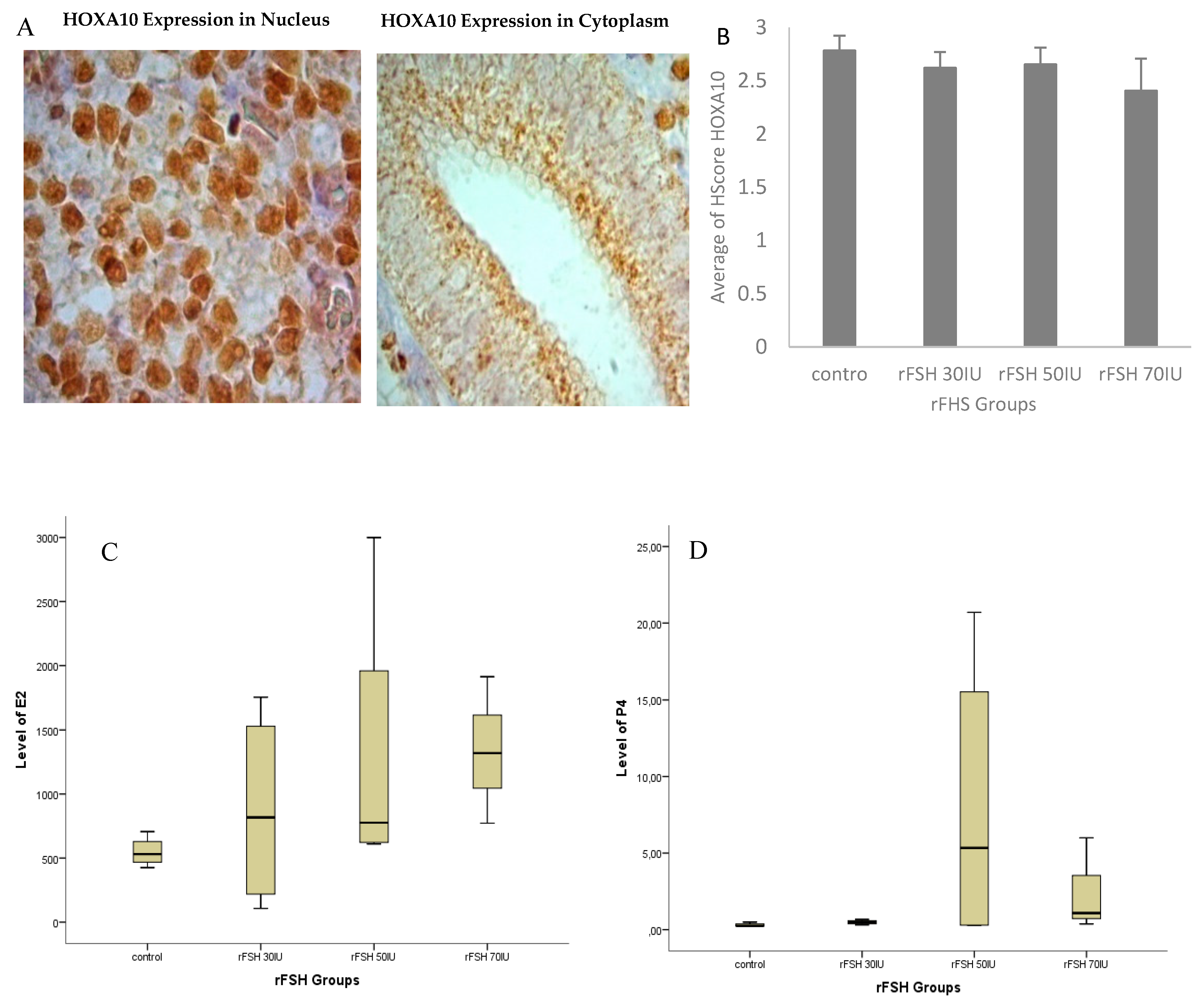
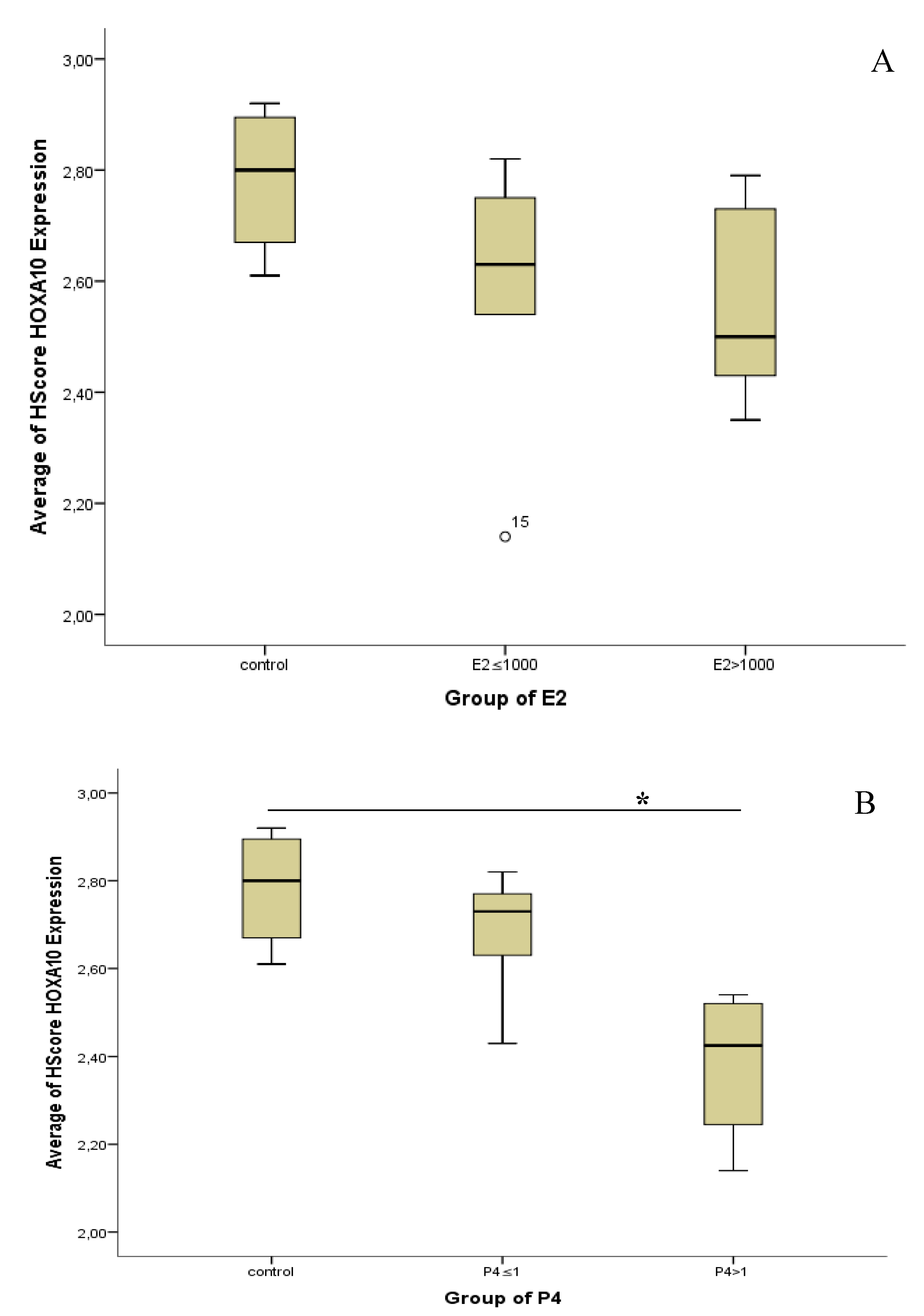
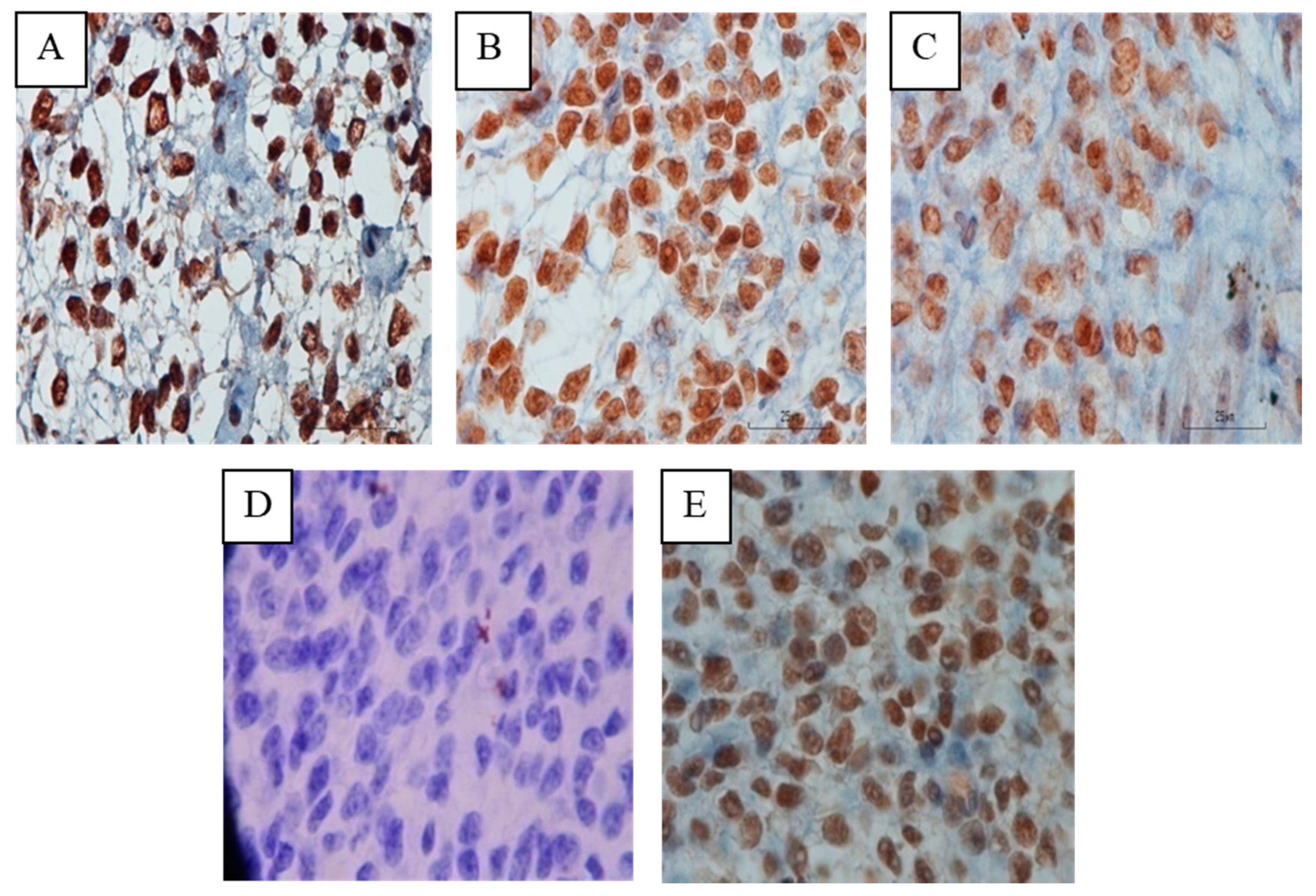
Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Sahar, N.; Mujihartini, N.; Pudjianto, D.A.; Pradhita, A.D.; Thuffi, R.; Kusmardi, K. Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity. Biomedicines 2019, 7, 83. https://doi.org/10.3390/biomedicines7040083
Sahar N, Mujihartini N, Pudjianto DA, Pradhita AD, Thuffi R, Kusmardi K. Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity. Biomedicines. 2019; 7(4):83. https://doi.org/10.3390/biomedicines7040083
Chicago/Turabian StyleSahar, Nurhuda, Ninik Mujihartini, Dwi Ari Pudjianto, Adhea Debby Pradhita, Rosalina Thuffi, and Kusmardi Kusmardi. 2019. "Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity" Biomedicines 7, no. 4: 83. https://doi.org/10.3390/biomedicines7040083
APA StyleSahar, N., Mujihartini, N., Pudjianto, D. A., Pradhita, A. D., Thuffi, R., & Kusmardi, K. (2019). Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity. Biomedicines, 7(4), 83. https://doi.org/10.3390/biomedicines7040083