

Palm Tocotrienol Preserves Trabecular Osteocyte Indices and Modulates the Expression of Osteocyte Markers in Ovariectomized Rats
 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Consideration
2.2. Source and Composition of Palm Tocotrienol
2.3. Animals and Treatment
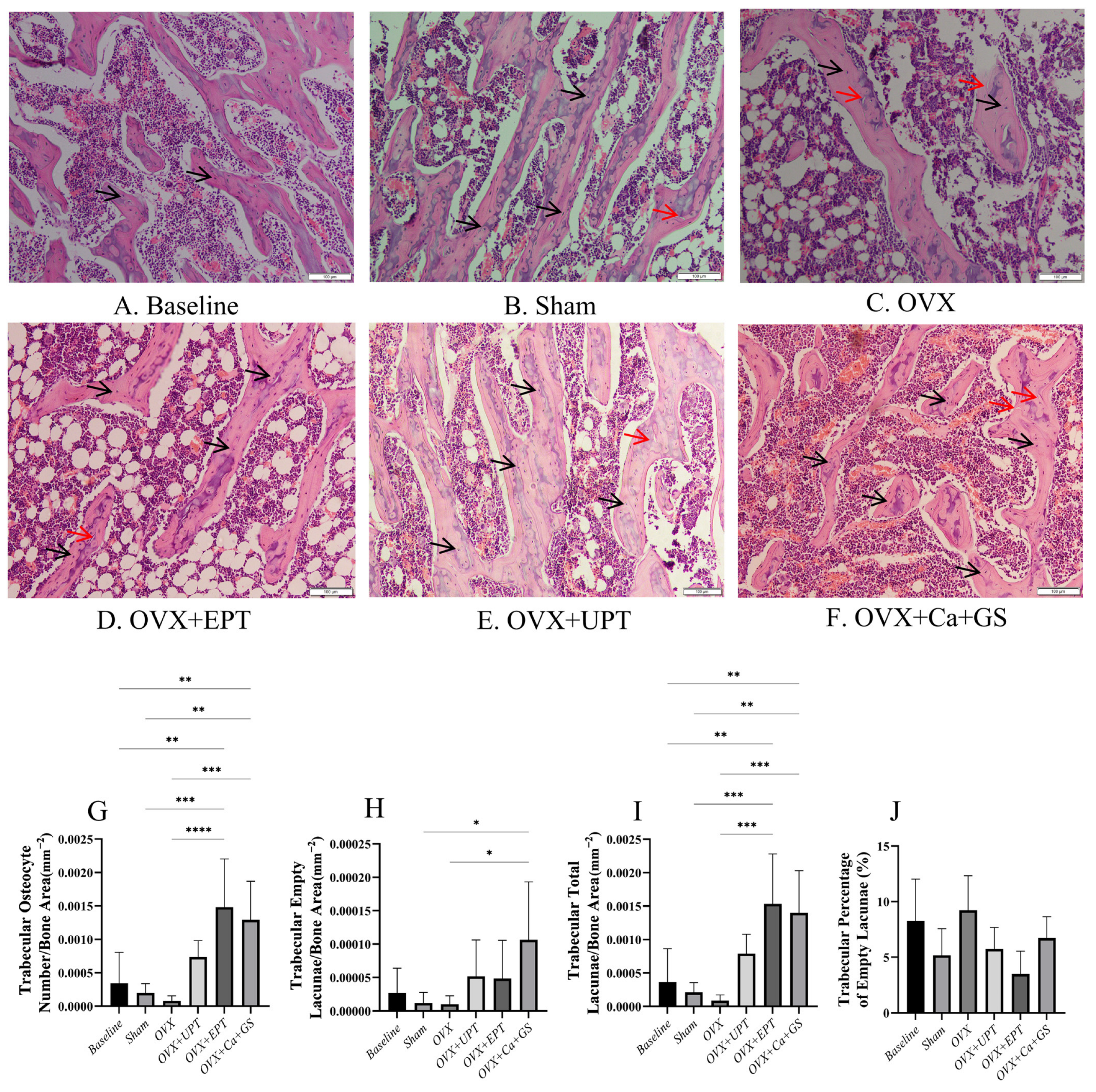
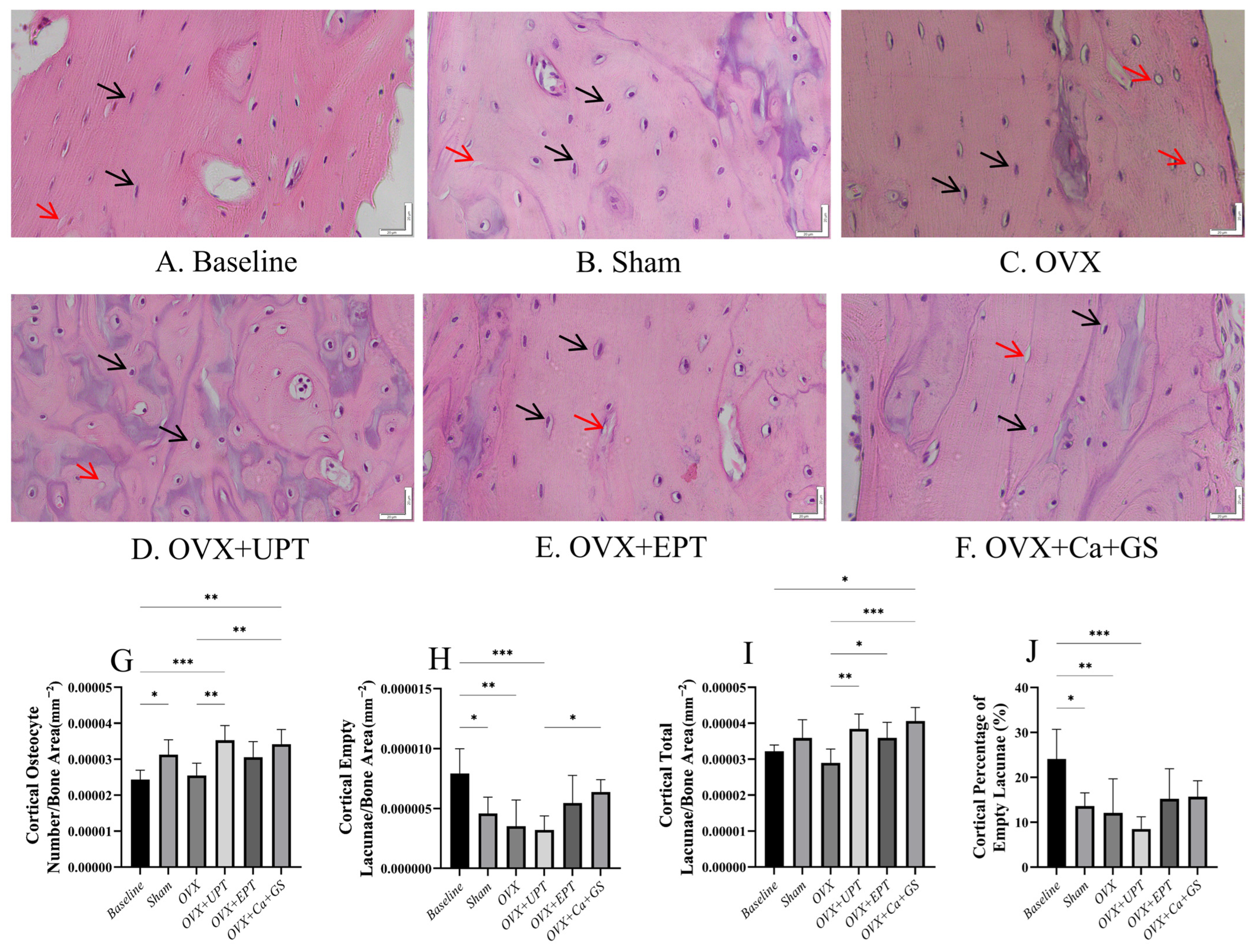
2.4. Measurement of Osteocyte Parameters
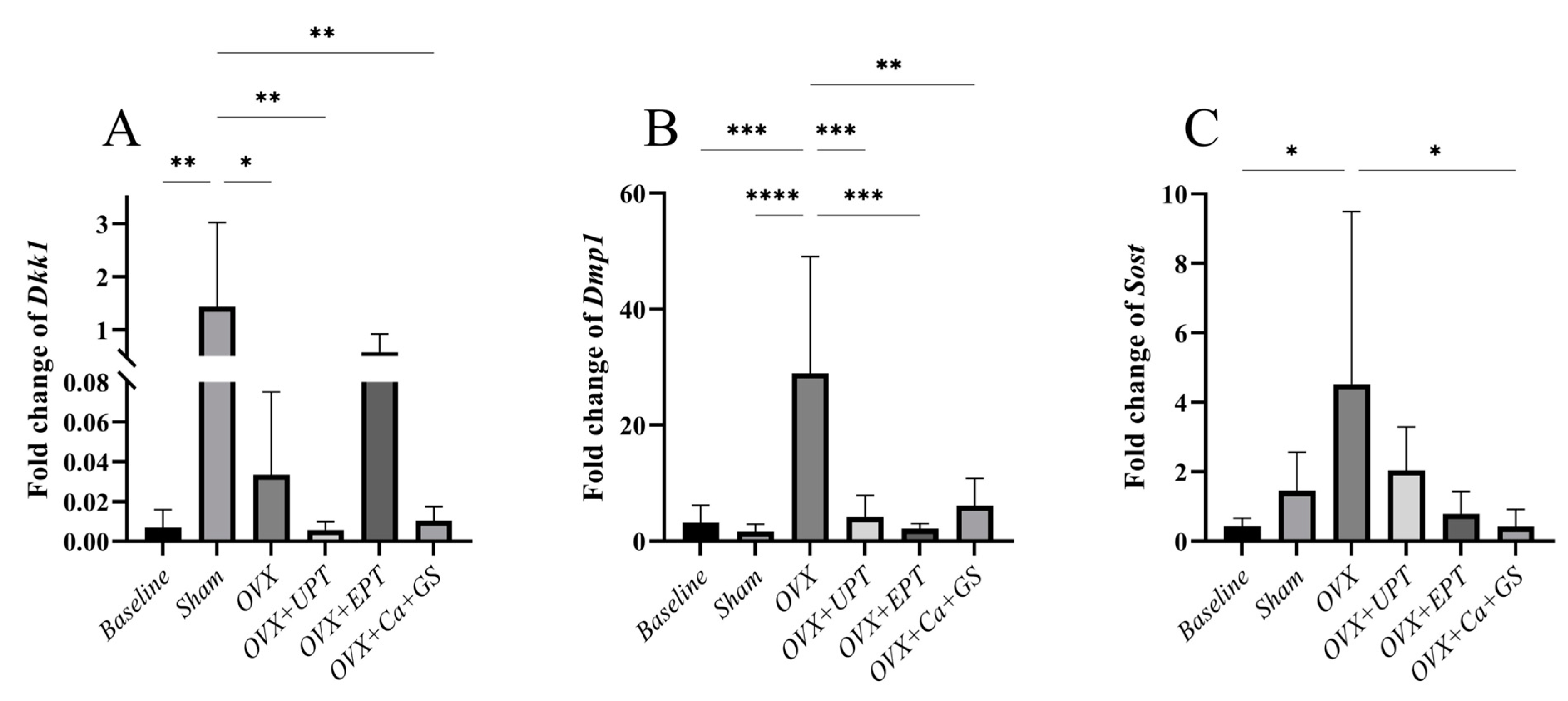
2.5. Gene Expression
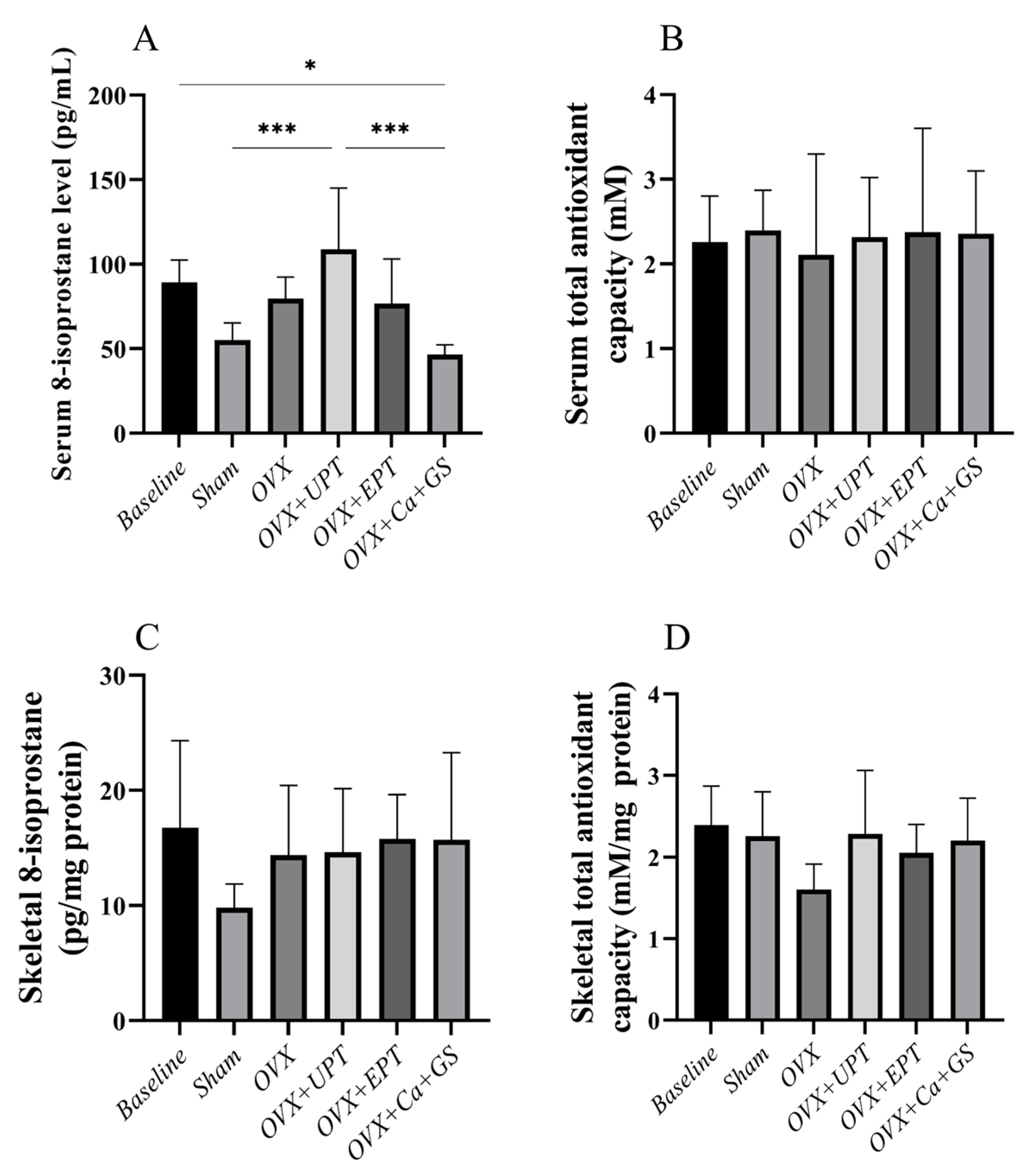
2.6. Measurement of Serum and Skeletal Redox Status
2.7. Statistical Analysis
3. Results
3.1. Osteocyte Parameters
3.2. mRNA Expression Levels of Osteocyte Markers
3.3. Redox Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aibar-Almazan, A.; Voltes-Martinez, A.; Castellote-Caballero, Y.; Afanador-Restrepo, D.F.; Carcelen-Fraile, M.D.C.; Lopez-Ruiz, E. Current Status of the Diagnosis and Management of Osteoporosis. Int. J. Mol. Sci. 2022, 23, 9465. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.L.; Cui, A.Y.; Hsu, C.J.; Peng, R.; Jiang, N.; Xu, X.H.; Ma, Y.G.; Liu, D.; Lu, H.D. Global, regional prevalence, and risk factors of osteoporosis according to the World Health Organization diagnostic criteria: A systematic review and meta-analysis. Osteoporos. Int. 2022, 33, 2137–2153. [Google Scholar] [CrossRef]
- de Villiers, T.J. Bone health and menopause: Osteoporosis prevention and treatment. Best. Pract. Res. Clin. Endocrinol. Metab. 2024, 38, 101782. [Google Scholar] [CrossRef]
- Salari, N.; Darvishi, N.; Bartina, Y.; Larti, M.; Kiaei, A.; Hemmati, M.; Shohaimi, S.; Mohammadi, M. Global prevalence of osteoporosis among the world older adults: A comprehensive systematic review and meta-analysis. J. Orthop. Surg. Res. 2021, 16, 669. [Google Scholar] [CrossRef]
- Noh, J.Y.; Yang, Y.; Jung, H. Molecular Mechanisms and Emerging Therapeutics for Osteoporosis. Int. J. Mol. Sci. 2020, 21, 7623. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.V.; Lighty, A.M.; Smith, B.J.; Ford Versypt, A.N. A review of mathematical modeling of bone remodeling from a systems biology perspective. Front. Syst. Biol. 2024, 4, 1368555. [Google Scholar] [CrossRef]
- Delgado-Calle, J.; Bellido, T. The osteocyte as a signaling cell. Physiol. Rev. 2022, 102, 379–410. [Google Scholar] [CrossRef]
- Li, X.; Kordsmeier, J.; Xiong, J. New Advances in Osteocyte Mechanotransduction. Curr. Osteoporos. Rep. 2021, 19, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, F.; Hayashi, M.; Ono, T.; Nakashima, T. The regulation of RANKL by mechanical force. J. Bone Min. Metab. 2021, 39, 34–44. [Google Scholar] [CrossRef]
- Ding, P.; Gao, C.; Gao, Y.; Liu, D.; Li, H.; Xu, J.; Chen, X.; Huang, Y.; Zhang, C.; Zheng, M.; et al. Osteocytes regulate senescence of bone and bone marrow. eLife 2022, 11, e81480. [Google Scholar] [CrossRef]
- Kim, J.H.; Liu, X.; Wang, J.; Chen, X.; Zhang, H.; Kim, S.H.; Cui, J.; Li, R.; Zhang, W.; Kong, Y.; et al. Wnt signaling in bone formation and its therapeutic potential for bone diseases. Ther. Adv. Musculoskelet. Dis. 2013, 5, 13–31. [Google Scholar] [CrossRef] [PubMed]
- McNamara, L.M. Osteocytes and Estrogen Deficiency. Curr. Osteoporos. Rep. 2021, 19, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Fu, W.; Xu, H.; Liu, C.J. Pathogenic mechanisms of glucocorticoid-induced osteoporosis. Cytokine Growth Factor. Rev. 2023, 70, 54–66. [Google Scholar] [CrossRef]
- Faienza, M.F.; Ventura, A.; Marzano, F.; Cavallo, L. Postmenopausal osteoporosis: The role of immune system cells. Clin. Dev. Immunol. 2013, 2013, 575936. [Google Scholar] [CrossRef]
- Lello, S.; Scambia, G.; Capozzi, A. Treatment options for postmenopausal osteoporosis. Maturitas 2023, 173, 33. [Google Scholar] [CrossRef]
- Skjødt, M.K.; Frost, M.; Abrahamsen, B. Side effects of drugs for osteoporosis and metastatic bone disease. Br. J. Clin. Pharmacol. 2019, 85, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Yeganehjoo, H.; Zamani, N.; Chitkara, S.; Shah, A.K. The Potential Role of Tocotrienols Against Cardiovascular Diseases. In Lipophilic Vitamins in Health and Disease; Tappia, P.S., Shah, A.K., Dhalla, N.S., Eds.; Springer International Publishing: Cham, Switzerland, 2024; pp. 149–161. [Google Scholar] [CrossRef]
- Mohd Zaffarin, A.S.; Ng, S.F.; Ng, M.H.; Hassan, H.; Alias, E. Pharmacology and Pharmacokinetics of Vitamin E: Nanoformulations to Enhance Bioavailability. Int. J. Nanomed. 2020, 15, 9961–9974. [Google Scholar] [CrossRef]
- Meister, M.L.; Mo, H.; Ji, X.; Shen, C.-L. Tocotrienols in Bone Protection: Evidence from Preclinical Studies. eFood 2020, 1, 217–225. [Google Scholar] [CrossRef]
- Mo, H.; Yeganehjoo, H.; Shah, A.; Mo, W.K.; Soelaiman, I.N.; Shen, C.L. Mevalonate-suppressive dietary isoprenoids for bone health. J. Nutr. Biochem. 2012, 23, 1543–1551. [Google Scholar] [CrossRef]
- Kim, K.W.; Kim, B.M.; Won, J.Y.; Min, H.K.; Lee, S.J.; Lee, S.H.; Kim, H.R. Tocotrienol regulates osteoclastogenesis in rheumatoid arthritis. Korean J. Intern. Med. 2021, 36, S273–S282. [Google Scholar] [CrossRef]
- Shah, A.K.; Yeganehjoo, H. The stimulatory impact of d-δ-Tocotrienol on the differentiation of murine MC3T3-E1 preosteoblasts. Mol. Cell Biochem. 2019, 462, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Soelaiman, I.N.; Ming, W.; Abu Bakar, R.; Hashnan, N.A.; Mohd Ali, H.; Mohamed, N.; Muhammad, N.; Shuid, A.N. Palm tocotrienol supplementation enhanced bone formation in oestrogen-deficient rats. Int. J. Endocrinol. 2012, 2012, 532862. [Google Scholar] [CrossRef]
- Ibrahim, N.; Mohd Noor, H.; Shuid, A.N.; Mohamad, S.; Abdul Malik, M.M.; Jayusman, P.A.; Shuid, A.N.; Naina Mohamed, I. Osteoprotective Effects in Postmenopausal Osteoporosis Rat Model: Oral Tocotrienol vs. Intraosseous Injection of Tocotrienol-Poly Lactic-Co-Glycolic Acid Combination. Front. Pharmacol. 2021, 12, 706747. [Google Scholar] [CrossRef]
- Abdul-Majeed, S.; Mohamed, N.; Soelaiman, I.N. Effects of tocotrienol and lovastatin combination on osteoblast and osteoclast activity in estrogen-deficient osteoporosis. Evid.-Based Complement. Altern. Med. 2012, 2012, 960742. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.; Khamis, M.F.; Mod Yunoh, M.F.; Abdullah, S.; Mohamed, N.; Shuid, A.N. Targeted delivery of lovastatin and tocotrienol to fracture site promotes fracture healing in osteoporosis model: Micro-computed tomography and biomechanical evaluation. PLoS ONE 2014, 9, e115595. [Google Scholar] [CrossRef]
- Ibrahim, N.; Mohamed, N.; Soelaiman, I.N.; Shuid, A.N. The Effects of Targeted Deliveries of Lovastatin and Tocotrienol on Ossification-Related Gene Expressions in Fracture Healing in an Osteoporosis Rat Model. Int. J. Environ. Res. Public Health 2015, 12, 12958–12976. [Google Scholar] [CrossRef] [PubMed]
- Ekeuku, S.O.; Nor Muhamad, M.L.; Aminuddin, A.A.; Ahmad, F.; Wong, S.K.; Mark-Lee, W.F.; Chin, K.-Y. Effects of emulsified and non-emulsified palm tocotrienol on bone and joint health in ovariectomised rats with monosodium iodoacetate-induced osteoarthritis. Biomed. Pharmacother. 2024, 170, 115998. [Google Scholar] [CrossRef]
- Kohno, K.; Yamada, W.; Ishitsuka, A.; Sekine, M.; Virgona, N.; Ota, M.; Yano, T. Tocotrienol-rich fraction from annatto ameliorates expression of lysyl oxidase in human osteoblastic MG-63 cells. Biosci. Biotechnol. Biochem. 2020, 84, 526–535. [Google Scholar] [CrossRef]
- Brooks, R.; Kalia, P.; Ireland, D.C.; Beeton, C.; Rushton, N. Direct inhibition of osteoclast formation and activity by the vitamin E isomer gamma-tocotrienol. Int. J. Vitam. Nutr. Res. 2011, 81, 358–367. [Google Scholar] [CrossRef]
- Ha, H.; Lee, J.H.; Kim, H.N.; Lee, Z.H. α-Tocotrienol inhibits osteoclastic bone resorption by suppressing RANKL expression and signaling and bone resorbing activity. Biochem. Biophys. Res. Commun. 2011, 406, 546–551. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Ima-Nirwana, S. The Effects of Tocotrienol on Bone Peptides in a Rat Model of Osteoporosis Induced by Metabolic Syndrome: The Possible Communication between Bone Cells. Int. J. Environ. Res. Public Health 2019, 16, 3313. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Fikri, N.I.A.; Munesveran, K.; Hisham, N.S.N.; Lau, S.H.J.; Chin, K.-Y.; Fahami, N.A.M.; Saad, Q.H.M.; Kamisah, Y.; Abdullah, A.; et al. Effects of tocotrienol on osteocyte-mediated phosphate metabolism in high-carbohydrate high-fat diet-induced osteoporotic rats. J. Funct. Foods 2022, 96, 105213. [Google Scholar] [CrossRef]
- Mohamad, N.-V.; Ima-Nirwana, S.; Chin, K.-Y. Self-emulsified annatto tocotrienol improves bone histomorphometric parameters in a rat model of oestrogen deficiency through suppression of skeletal sclerostin level and RANKL/OPG ratio. Int. J. Med. Sci. 2021, 18, 3665–3673. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, J. The role of Wnt signaling pathway in atherosclerosis and its relationship with angiogenesis. Exp. Ther. Med. 2018, 16, 1975–1981. [Google Scholar] [CrossRef]
- de Farias Junior, G.C.; de Sousa Neto, I.V.; Guzzoni, V.; Pisani, G.D.; Royer, C.; de Lima, C.L.; de Assis Rocha Neves, F.; Bogni, F.H.; Nonaka, K.O.; Durigan, J.L.Q.; et al. Remodeling process in bone of aged rats in response to resistance training. Life Sci. 2020, 256, 118008. [Google Scholar] [CrossRef]
- Rezai Rad, M.; Liu, D.; He, H.; Brooks, H.; Xiao, M.; Wise, G.E.; Yao, S. The role of dentin matrix protein 1 (DMP1) in regulation of osteogenic differentiation of rat dental follicle stem cells (DFSCs). Arch. Oral Biol. 2015, 60, 546–556. [Google Scholar] [CrossRef]
- Allison, H.; O’Sullivan, L.M.; McNamara, L.M. Temporal changes in cortical microporosity during estrogen deficiency associated with perilacunar resorption and osteocyte apoptosis: A pilot study. Bone Rep. 2022, 16, 101590. [Google Scholar] [CrossRef]
- Tiede-Lewis, L.M.; Xie, Y.; Hulbert, M.A.; Campos, R.; Dallas, M.R.; Dusevich, V.; Bonewald, L.F.; Dallas, S.L. Degeneration of the osteocyte network in the C57BL/6 mouse model of aging. Aging 2017, 9, 2190–2208. [Google Scholar] [CrossRef]
- Martin, A.; Kentrup, D. The Role of DMP1 in CKD-MBD. Curr. Osteoporos. Rep. 2021, 19, 500–509. [Google Scholar] [CrossRef]
- Gao, Y.; Chen, N.; Fu, Z.; Zhang, Q. Progress of Wnt Signaling Pathway in Osteoporosis. Biomolecules 2023, 13, 483. [Google Scholar] [CrossRef]
- Vasiliadis, E.S.; Evangelopoulos, D.-S.; Kaspiris, A.; Benetos, I.S.; Vlachos, C.; Pneumaticos, S.G. The Role of Sclerostin in Bone Diseases. J. Clin. Med. 2022, 11, 806. [Google Scholar] [CrossRef] [PubMed]
- Starlinger, J.; Santol, J.; Kaiser, G.; Sarahrudi, K. Close negative correlation of local and circulating Dickkopf-1 and Sclerostin levels during human fracture healing. Sci. Rep. 2024, 14, 6524. [Google Scholar] [CrossRef] [PubMed]
- Nur Farhana, M.F.; James Jam, J.; Chua, K.H.; Ekram, A.; Chin, K.Y.; Ima Nirwana, S. Comparing the Effects of Alpha-Tocopherol and Tocotrienol Isomers on Osteoblasts hFOB 1.19 Cultured on Bovine Bone Scaffold. Sains Malays. 2021, 50, 2319–2328. [Google Scholar] [CrossRef]
- Signorini, C.; Moretti, E.; Collodel, G. Role of isoprostanes in human male infertility. Syst. Biol. Reprod. Med. 2020, 66, 291–299. [Google Scholar] [CrossRef]
- Rubio, C.P.; Hernández-Ruiz, J.; Martinez-Subiela, S.; Tvarijonaviciute, A.; Ceron, J.J. Spectrophotometric assays for total antioxidant capacity (TAC) in dog serum: An update. BMC Vet. Res. 2016, 12, 166. [Google Scholar] [CrossRef]
- Mohamad, N.V.; Ima-Nirwana, S.; Chin, K.Y. Therapeutic potential of annatto tocotrienol with self-emulsifying drug delivery system in a rat model of postmenopausal bone loss. Biomed. Pharmacother. 2021, 137, 111368. [Google Scholar] [CrossRef]
- Vakili, S.; Zal, F.; Mostafavipour, Z.; Savardashtaki, A.; Jafari, K.M.; Hassanpour, A. Effects of quercetin and vitamin E on ovariectomy-induced oxidative stress in rat serum and tibia. Arch. Biol. Sci. 2020, 72, 95–104. [Google Scholar] [CrossRef]
- Maniam, S.; Mohamed, N.; Shuid, A.N.; Soelaiman, I.N. Palm Tocotrienol Exerted Better Antioxidant Activities in Bone than α-Tocopherol. Basic Clin. Pharmacol. Toxicol. 2008, 103, 55–60. [Google Scholar] [CrossRef]
- Weivoda, M.M.; Youssef, S.J.; Oursler, M.J. Sclerostin expression and functions beyond the osteocyte. Bone 2017, 96, 45–50. [Google Scholar] [CrossRef]
- Terasawa, M.; Shimokawa, R.; Terashima, T.; Ohya, K.; Takagi, Y.; Shimokawa, H. Expression of dentin matrix protein 1 (DMP1) in nonmineralized tissues. J. Bone Min. Metab. 2004, 22, 430–438. [Google Scholar] [CrossRef]
- Roach, H.I.; Mehta, G.; Oreffo, R.O.C.; Clarke, N.M.P.; Cooper, C. Temporal Analysis of Rat Growth Plates: Cessation of Growth with Age Despite Presence of a Physis. J. Histochem. Cytochem. 2003, 51, 373–383. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ekeuku, S.O.; Zarir, S.Z.; Razali, A.N.; Mohamad Zaidi, S.; Mohamed Ali Jinnah, N.H.; Nor Muhamad, M.L.; Wong, S.K.; Chin, K.-Y. Palm Tocotrienol Preserves Trabecular Osteocyte Indices and Modulates the Expression of Osteocyte Markers in Ovariectomized Rats. Biomedicines 2025, 13, 1220. https://doi.org/10.3390/biomedicines13051220
Ekeuku SO, Zarir SZ, Razali AN, Mohamad Zaidi S, Mohamed Ali Jinnah NH, Nor Muhamad ML, Wong SK, Chin K-Y. Palm Tocotrienol Preserves Trabecular Osteocyte Indices and Modulates the Expression of Osteocyte Markers in Ovariectomized Rats. Biomedicines. 2025; 13(5):1220. https://doi.org/10.3390/biomedicines13051220
Chicago/Turabian StyleEkeuku, Sophia Ogechi, Shafiq Zikry Zarir, Anis Nazira Razali, Syamima Mohamad Zaidi, Noor Halinah Mohamed Ali Jinnah, Muhamed Lahtif Nor Muhamad, Sok Kuan Wong, and Kok-Yong Chin. 2025. "Palm Tocotrienol Preserves Trabecular Osteocyte Indices and Modulates the Expression of Osteocyte Markers in Ovariectomized Rats" Biomedicines 13, no. 5: 1220. https://doi.org/10.3390/biomedicines13051220
APA StyleEkeuku, S. O., Zarir, S. Z., Razali, A. N., Mohamad Zaidi, S., Mohamed Ali Jinnah, N. H., Nor Muhamad, M. L., Wong, S. K., & Chin, K.-Y. (2025). Palm Tocotrienol Preserves Trabecular Osteocyte Indices and Modulates the Expression of Osteocyte Markers in Ovariectomized Rats. Biomedicines, 13(5), 1220. https://doi.org/10.3390/biomedicines13051220