
Co-Occurrence of Helicobacter pylori and Candida spp. Infections in the Pathogenesis of Gastrointestinal Diseases
 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Methodology
3. Helicobacter pylori
4. Candida albicans and Candida spp.
5. Epidemiology of H. pylori Infections

{kind=link}
| Global Prevalence of H. pylori | |
|---|---|
| Region | Prevalence Estimates, % |
| African region | 56.5 |
| Eastern Mediterranean region | 56.5 |
| European region | 45.7 |
| Region of the Americas | 49.0 |
| Southeast Asia region | 44.2 |
| Western Pacific region | 49.4 |
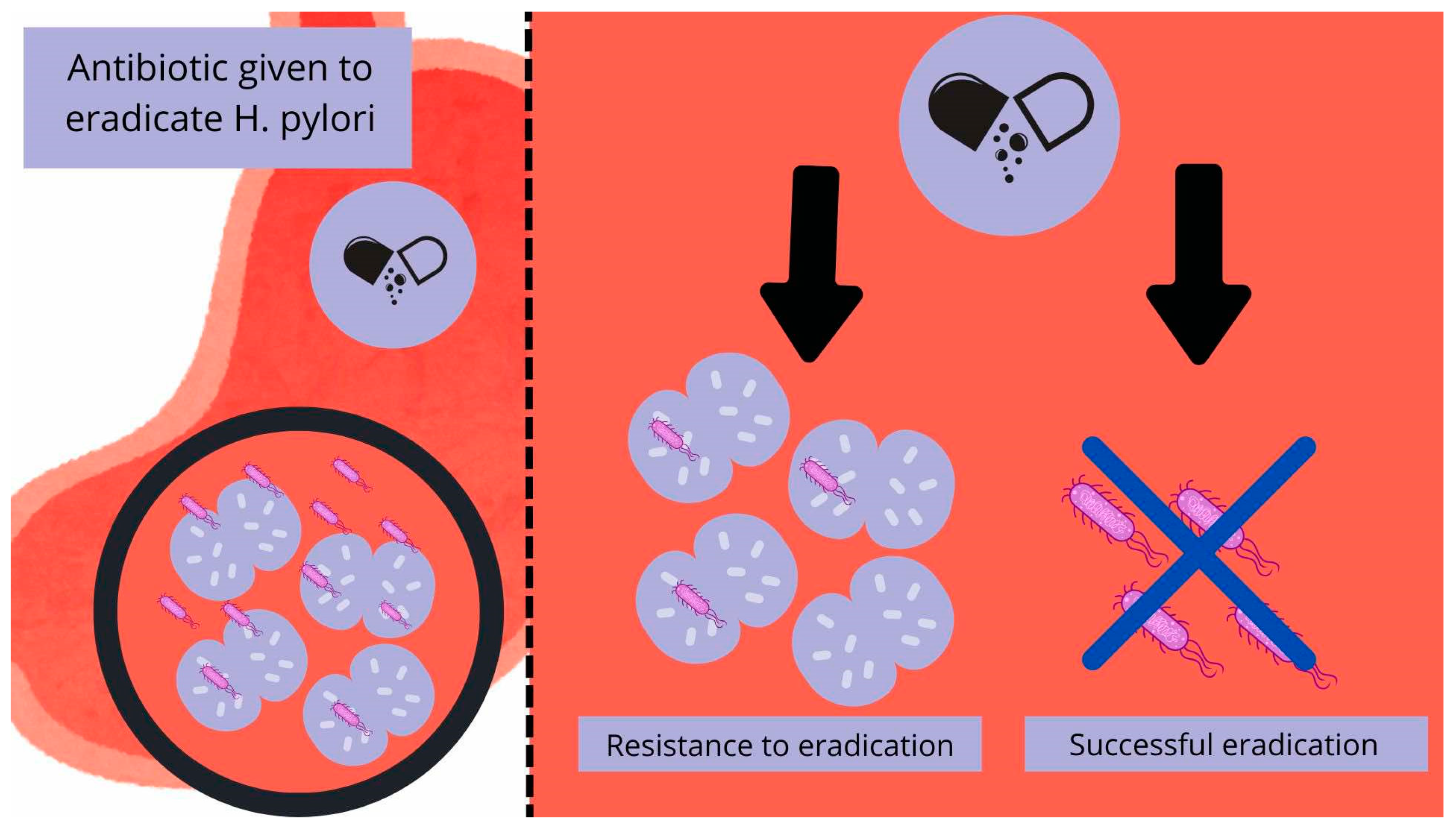
6. H. pylori Eradication and Its Resistance to Antibiotics
7. Interplay Between Candida spp. and H. pylori Infection
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Siavoshi, F.; Saniee, P. Vacuoles of Candida Yeast as a Specialized Niche for Helicobacter pylori. World J. Gastroenterol. 2014, 20, 5263–5273. [Google Scholar] [CrossRef] [PubMed]
- Hui, W.W.; Emerson, L.E.; Clapp, B.; Sheppe, A.E.; Sharma, J.; del Castillo, J.; Ou, M.; Maegawa, G.H.B.; Hoffman, C.; Larkin, J.; et al. The Cross-Kingdom Interaction between Helicobacter pylori and Candida Albicans. PLoS Pathog. 2021, 17, e1009515. [Google Scholar] [CrossRef]
- Ali, A.; AlHussaini, K.I. Helicobacter pylori: A Contemporary Perspective on Pathogenesis, Diagnosis and Treatment Strategies. Microorganisms 2024, 12, 222. [Google Scholar] [CrossRef] [PubMed]
- Kesharwani, A.; Dighe, O.R.; Lamture, Y. Role of Helicobacter pylori in Gastric Carcinoma: A Review. Cureus 2023, 15, e37205. [Google Scholar] [CrossRef]
- Baj, J.; Forma, A.; Sitarz, M.; Portincasa, P.; Garruti, G.; Krasowska, D.; Maciejewski, R. Helicobacter pylori Virulence Factors—Mechanisms of Bacterial Pathogenicity in the Gastric Microenvironment. Cells 2020, 10, 27. [Google Scholar] [CrossRef]
- Azevedo, N.F.; Huntington, J.; Goodman, K.J. The Epidemiology of Helicobacter pylori and Public Health Implications. Helicobacter 2009, 14 (Suppl. 1), 1–7. [Google Scholar] [CrossRef]
- Sachs, G.; Scott, D.R. Helicobacter pylori: Eradication or Preservation. F1000 Med. Rep. 2012, 4, 7. [Google Scholar] [CrossRef]
- Salvatori, S.; Marafini, I.; Laudisi, F.; Monteleone, G.; Stolfi, C. Helicobacter pylori and Gastric Cancer: Pathogenetic Mechanisms. Int. J. Mol. Sci. 2023, 24, 2895. [Google Scholar] [CrossRef]
- Goni, E.; Franceschi, F. Helicobacter pylori and Extragastric Diseases. Helicobacter 2016, 21 (Suppl. 1), 45–48. [Google Scholar] [CrossRef]
- Maiorana, F.; Neschuk, M.; Caronia, M.V.; Elizondo, K.; Robledo, M.L.; Schneider, A.; Veron, G.; Zapata, P.D.; Barreyro, F.J. The Interplay between Helicobacter pylori Infection and Rs738409 PNPLA3 in Metabolic Dysfunction-Associated Steatotic Liver Disease. PLoS ONE 2024, 19, e0310361. [Google Scholar] [CrossRef]
- Keikha, M.; Karbalaei, M. Potential Association between Bacterial Infections and Ischemic Stroke Based on Fifty Case-Control Studies: A Systematic Review and Meta-Analysis. New Microbes New Infect. 2022, 47, 100980. [Google Scholar] [CrossRef] [PubMed]
- Keikha, M.; Karbalaei, M. A Comprehensive Survey of the Relationship between Helicobacter pylori Infection and Atherosclerosis in the Iranian Population: A Systematic Review and Meta-Analysis. Arch. Iran Med. 2022, 25, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Arellano, L. Helicobacter pylori and Neurological Diseases: Married by the Laws of Inflammation. World J. Gastrointest. Pathophysiol. 2014, 5, 400. [Google Scholar] [CrossRef]
- Kusters, J.G.; Van Vliet, A.H.M.; Kuipers, E.J. Pathogenesis of Helicobacter pylori Infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y.; Sheu, B.S.; Wu, J.J. Helicobacter pylori Infection: An Overview of Bacterial Virulence Factors and Pathogenesis. Biomed. J. 2016, 39, 14. [Google Scholar] [CrossRef]
- Yang, T.; Li, J.; Zhang, Y.; Deng, Z.; Cui, G.; Yuan, J.; Sun, J.; Wu, X.; Hua, D.; Xiang, S.; et al. Intracellular Presence of Helicobacter pylori Antigen and Genes within Gastric and Vaginal Candida. PLoS ONE 2024, 19, e0298442. [Google Scholar] [CrossRef]
- Zou, Y.; Chen, X.; Sun, Y.; Li, P.; Xu, M.; Fang, P.; Zhang, S.; Yuan, G.; Deng, X.; Hu, H. Antibiotics-Free Nanoparticles Eradicate Helicobacter pylori Biofilms and Intracellular Bacteria. J. Control. Release 2022, 348, 370–385. [Google Scholar] [CrossRef]
- Sánchez-Alonzo, K.; Matamala-Valdés, L.; Parra-Sepúlveda, C.; Bernasconi, H.; Campos, V.L.; Smith, C.T.; Sáez, K.; García-Cancino, A. Intracellular Presence of Helicobacter pylori and Its Virulence-Associated Genotypes within the Vaginal Yeast of Term Pregnant Women. Microorganisms 2021, 9, 131. [Google Scholar] [CrossRef]
- Tang, Z.; Fu, L.; Liu, R.; Chen, Y.; Bie, M.; Wang, B. Mechanisms of intracellular Helicobacter pylori infection and clinical considerations. J. Sichuan Univ. 2023, 54, 1300. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, Q.L.; Cheng, D.D.; Xu, W.T.; Lu, N.H. Adhesion and Invasion of Gastric Mucosa Epithelial Cells by Helicobacter pylori. Front. Cell. Infect. Microbiol. 2016, 6, 227385. [Google Scholar] [CrossRef]
- McBain, A.J.; O’Neill, C.A.; Oates, A. Skin Microbiology. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Dowd, F.J. Candida Albicans Infections. In Reference Module in Biomedical Research; Creighton University: Omaha, NE, USA, 2014. [Google Scholar] [CrossRef]
- Wiles, C.M.; Mackenzie, D.W.R. Fungal Diseases of the Central Nervous System. Infect. Nerv. Syst. 1987, 93–117. [Google Scholar] [CrossRef]
- Hiengrach, P.; Panpetch, W.; Chindamporn, A.; Leelahavanichkul, A. Helicobacter pylori, Protected from Antibiotics and Stresses Inside Candida Albicans Vacuoles, Cause Gastritis in Mice. Int. J. Mol. Sci. 2022, 23, 8568. [Google Scholar] [CrossRef] [PubMed]
- The ALS Gene Family of Candida Albicans. Trends Microbiol. 2001, 9, 176–180. [CrossRef] [PubMed]
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida Albicans Secreted Aspartyl Proteinases in Virulence and Pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef]
- Gaffen, S.L.; Hernández-Santos, N.; Peterson, A.C. IL-17 Signaling in Host Defense Against Candida Albicans. Immunol. Res. 2011, 50, 181. [Google Scholar] [CrossRef]
- Borka Balas, R.; Meliț, L.E.; Mărginean, C.O. Worldwide Prevalence and Risk Factors of Helicobacter pylori Infection in Children. Children 2022, 9, 1359. [Google Scholar] [CrossRef]
- Tran, V.; Saad, T.; Tesfaye, M.; Walelign, S.; Wordofa, M.; Abera, D.; Desta, K.; Tsegaye, A.; Ay, A.; Taye, B. Helicobacter pylori (H. pylori) Risk Factor Analysis and Prevalence Prediction: A Machine Learning-Based Approach. BMC Infect. Dis. 2022, 22, 655. [Google Scholar] [CrossRef]
- Smith, S.; Jolaiya, T.; Fowora, M.; Palamides, P.; Ngoka, F.; Bamidele, M.; Lesi, O.; Onyekwere, C.; Ugiagbe, R.; Agbo, I.; et al. Clinical and Socio- Demographic Risk Factors for Acquisition of Helicobacter pylori Infection in Nigeria. Asian Pac. J. Cancer Prev. 2018, 19, 1851–1857. [Google Scholar] [CrossRef]
- Chen, Y.C.; Malfertheiner, P.; Yu, H.T.; Kuo, C.L.; Chang, Y.Y.; Meng, F.T.; Wu, Y.X.; Hsiao, J.L.; Chen, M.J.; Lin, K.P.; et al. Global Prevalence of Helicobacter pylori Infection and Incidence of Gastric Cancer Between 1980 and 2022. Gastroenterology 2024, 166, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Ernst, P.B.; Gold, B.D. The Disease Spectrum of Helicobacter pylori: The Immunopathogenesis of Gastroduodenal Ulcer and Gastric Cancer. Annu. Rev. Microbiol. 2000, 54, 615–640. [Google Scholar] [CrossRef]
- McConaghy, J.R.; Decker, A.; Nair, S. Peptic Ulcer Disease and H. pylori Infection: Common Questions and Answers. Am. Fam. Physician 2023, 107, 165–172. [Google Scholar] [PubMed]
- Chen, T.H.; Cheng, H.T.; Yeh, C.T. Epidemiology Changes in Peptic Ulcer Diseases 18 Years Apart Explored from the Genetic Aspects of Helicobacter pylori. Transl. Res. 2021, 232, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Zheng, M.; Jiang, J.; Cao, D.; Wu, Y.; Zhang, Y.; Fu, Y.; Cao, X. Positive H. pylori Status Predicts Better Prognosis of Non-Cardiac Gastric Cancer Patients: Results from Cohort Study and Meta-Analysis. BMC Cancer 2022, 22, 155. [Google Scholar] [CrossRef]
- He, J.J.; Hu, W.C.; Ouyang, Q.; Zhang, S.W.; He, L.J.; Chen, W.Y.; Li, X.Z.; Hu, C.J. Helicobacter pylori Infection Induces Stem Cell-like Properties in Correa Cascade of Gastric Cancer. Cancer Lett. 2022, 542, 215764. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Xu, Y.; Wang, P.; Zhao, Y.; Wan, C. Research Progress on Molecular Mechanism of Pyroptosis Caused by Helicobacter pylori in Gastric Cancer. Ann. Med. Surg. 2024, 86, 2016–2022. [Google Scholar] [CrossRef]
- Kumar, S.; Metz, D.C.; Ellenberg, S.; Kaplan, D.E.; Goldberg, D.S. Risk Factors and Incidence of Gastric Cancer After Detection of Helicobacter pylori Infection: A Large Cohort Study. Gastroenterology 2020, 158, 527–536.e7. [Google Scholar] [CrossRef]
- Watari, J.; Das, K.K.; Amenta, P.S.; Tanabe, H.; Tanaka, A.; Geng, X.; Lin, J.J.C.; Kohgo, Y.; Das, K.M. Effect of Eradication of Helicobacter pylori on the Histology and Cellular Phenotype of Gastric Intestinal Metaplasia. Clin. Gastroenterol. Hepatol. 2008, 6, 409–417. [Google Scholar] [CrossRef]
- Pimanov, S.I.; Makarenko, E.V.; Voropaeva, A.V.; Matveenko, M.E.; Voropaev, E.V. Helicobacter pylori Eradication Improves Gastric Histology and Decreases Serum Gastrin, Pepsinogen I and Pepsinogen II Levels in Patients with Duodenal Ulcer. J. Gastroenterol. Hepatol. 2008, 23, 1666–1671. [Google Scholar] [CrossRef]
- Rokkas, T.; Pistiolas, D.; Sechopoulos, P.; Robotis, I.; Margantinis, G. The Long-Term Impact of Helicobacter pylori Eradication on Gastric Histology: A Systematic Review and Meta-Analysis. Helicobacter 2007, 12 (Suppl. 2), 32–38. [Google Scholar] [CrossRef]
- Li, W.Q.; Ma, J.L.; Zhang, L.; Brown, L.M.; Li, J.Y.; Shen, L.; Pan, K.F.; Liu, W.D.; Hu, Y.; Han, Z.X.; et al. Effects of Helicobacter pylori Treatment on Gastric Cancer Incidence and Mortality in Subgroups. J. Natl. Cancer Inst. 2014, 106, dju116. [Google Scholar] [CrossRef]
- Gisbert, J.P.; McNicholl, A.G. Optimization Strategies Aimed to Increase the Efficacy of H. pylori Eradication Therapies. Helicobacter 2017, 22, e12392. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Megraud, F.; Rokkas, T.; Gisbert, J.P.; Liou, J.-M.; Schulz, C.; Gasbarrini, A.; Hunt, R.H. Management of Helicobacter pylori Infection: The Maastricht VI/Florence Consensus Report. Gut 2022, 71, 1724–1762. [Google Scholar] [CrossRef] [PubMed]
- Salazar, C.O.; Cardenas, V.M.; Reddy, R.K.; Dominguez, D.C.; Snyder, L.K.; Graham, D.Y. Greater than 95% Success with 14-Day Bismuth Quadruple Anti- Helicobacter pylori Therapy: A Pilot Study in US Hispanics. Helicobacter 2012, 17, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Ford, A.C.; Khan, K.J.; Gisbert, J.P.; Forman, D.; Leontiadis, G.I.; Tse, F.; Calvet, X.; Fallone, C.; Fischbach, L.; et al. Optimum Duration of Regimens for Helicobacter pylori Eradication. Cochrane Database Syst. Rev. 2013, 2013, 1–205. [Google Scholar] [CrossRef]
- Thung, I.; Aramin, H.; Vavinskaya, V.; Gupta, S.; Park, J.Y.; Crowe, S.E.; Valasek, M.A. Review Article: The Global Emergence of Helicobacter pylori Antibiotic Resistance. Aliment. Pharmacol. Ther. 2016, 43, 514–533. [Google Scholar] [CrossRef]
- ACG. American College of Gastroenterology, Guideline on Treatment of Helicobacter pylori: New Recommendations… Will Practice Change? Available online: https://gi.org/journals-publications/ebgi/schoenfeld_sep2024/ (accessed on 22 April 2025).
- McNicholl, A.G.; Linares, P.M.; Nyssen, O.P.; Calvet, X.; Gisbert, J.P. Meta-Analysis: Esomeprazole or Rabeprazole vs. First-Generation Pump Inhibitors in the Treatment of Helicobacter pylori Infection. Aliment. Pharmacol. Ther. 2012, 36, 414–425. [Google Scholar] [CrossRef]
- Gong, Y.; Yuan, Y. Resistance Mechanisms of Helicobacter pylori and Its Dual Target Precise Therapy. Crit. Rev. Microbiol. 2018, 44, 371–392. [Google Scholar] [CrossRef]
- Tshibangu-Kabamba, E.; Yamaoka, Y. Helicobacter pylori Infection and Antibiotic Resistance—from Biology to Clinical Implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 613–629. [Google Scholar] [CrossRef]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of Antibiotic Resistance in Helicobacter pylori: A Systematic Review and Meta-Analysis in World Health Organization Regions. Gastroenterology 2018, 155, 1372–1382.e17. [Google Scholar] [CrossRef]
- Salahi-Niri, A.; Nabavi-Rad, A.; Monaghan, T.M.; Rokkas, T.; Doulberis, M.; Sadeghi, A.; Zali, M.R.; Yamaoka, Y.; Tacconelli, E.; Yadegar, A. Global Prevalence of Helicobacter pylori Antibiotic Resistance among Children in the World Health Organization Regions between 2000 and 2023: A Systematic Review and Meta-Analysis. BMC Med. 2024, 22, 1–17. [Google Scholar] [CrossRef]
- Katzenstein, A.L.A.; Maksem, J. Candidal Infection of Gastric Ulcers. Histology, Incidence, and Clinical Significance. Am. J. Clin. Pathol. 1979, 71, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.B.; Jenkins, D. Gastro-Oesophageal Candidiasis. Gut 1982, 23, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Robin Warren, J.; Marshall, B. Unidentified Curved Bacilli On Gastric Epithelium In Active Chronic Gastritis. Lancet 1983, 321, 1273–1275. [Google Scholar] [CrossRef]
- Zwolińska-Wcisło, M.; Budak, A.; Trojanowska, D.; Bogdał, J.; Stachura, J. Fungal Colonization of the Stomach and Its Clinical Relevance. Mycoses 1998, 41, 327–334. [Google Scholar] [CrossRef]
- Medical Science Monitor, Effect of Fungal Colonization of Gastric Mucosa on the Course of Gastric Ulcers Healing. Article Abstract #421150. Available online: https://medscimonit.com/abstract/index/idArt/421150 (accessed on 23 March 2025).
- PubMed, Assessment of Co-Existence of Helicobacter pylori and Candida Fungi in Diseases of the Upper Gastrointestinal Tract. Available online: https://pubmed.ncbi.nlm.nih.gov/20224149/ (accessed on 9 March 2025).
- Wang, Y.H.; Lv, Z.F.; Zhong, Y.; Liu, D.S.; Chen, S.P.; Xie, Y. The Internalization of Helicobacter pylori Plays a Role in the Failure of H. pylori Eradication. Helicobacter 2017, 22, e12324. [Google Scholar] [CrossRef]
- D’Enfert, C.; Kaune, A.K.; Alaban, L.R.; Chakraborty, S.; Cole, N.; Delavy, M.; Kosmala, D.; Marsaux, B.; Fróis-Martins, R.; Morelli, M.; et al. The Impact of the Fungus-Host-Microbiota Interplay upon Candida Albicans Infections: Current Knowledge and New Perspectives. FEMS Microbiol. Rev. 2021, 45, 14. [Google Scholar] [CrossRef]
- Heydari, S.; Siavoshi, F.; Jazayeri, M.H.; Sarrafnejad, A.; Saniee, P. Helicobacter pylori Release from Yeast as a Vesicle-Encased or Free Bacterium. Helicobacter 2020, 25, e12725. [Google Scholar] [CrossRef]
- Plant cell biology, Volume 1, PWN. Available online: https://ksiegarnia.pwn.pl/Biologia-komorki-roslinnej-Tom-1-Struktura,68710268,p.html?srsltid=AfmBOopJzuyr-7vIiLAziGblmX-O8gomgJuV5XVfUzCd4WosnmEx6E7J (accessed on 9 March 2025).
- Siavoshi, F.; Heydari, S.; Shafiee, M.; Ahmadi, S.; Saniee, P.; Sarrafnejad, A.; Kolahdoozan, S. Sequestration inside the Yeast Vacuole May Enhance Helicobacter pylori Survival against Stressful Condition. Infect. Genet. Evol. 2019, 69, 127–133. [Google Scholar] [CrossRef]
- Scott, D.R.; Sachs, G.; Marcus, E.A. The Role of Acid Inhibition in Helicobacter pylori Eradication. F1000Research 2016, 5, 1747. [Google Scholar] [CrossRef]
- Yang, J.C.; Lu, C.W.; Lin, C.J. Treatment of Helicobacter pylori Infection: Current Status and Future Concepts. World J. Gastroenterol. 2014, 20, 5283–5293. [Google Scholar] [CrossRef]
- Siavoshi, F.; Sahraee, M.; Ebrahimi, H.; Sarrafnejad, A.; Saniee, P. Natural Fruits, Flowers, Honey, and Honeybees Harbor Helicobacter pylori-Positive Yeasts. Helicobacter 2018, 23, e12471. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Alonzo, K.; Silva-Mieres, F.; Arellano-Arriagada, L.; Parra-Sepúlveda, C.; Bernasconi, H.; Smith, C.T.; Campos, V.L.; García-Cancino, A. Nutrient Deficiency Promotes the Entry of Helicobacter pylori Cells into Candida Yeast Cells. Biology 2021, 10, 426. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Alonzo, K.; Parra-Sepúlveda, C.; Vega, S.; Bernasconi, H.; Campos, V.L.; Smith, C.T.; Sáez, K.; García-Cancino, A. In Vitro Incorporation of Helicobacter pylori into Candida Albicans Caused by Acidic PH Stress. Pathogens 2020, 9, 489. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Alonzo, K.; Belmar, L.; Parra-Sepúlveda, C.; Bernasconi, H.; Campos, V.L.; Smith, C.T.; Sáez, K.; García-Cancino, A. Antibiotics as a Stressing Factor Triggering the Harboring of Helicobacter pylori J99 within Candida Albicans ATCC10231. Pathogens 2021, 10, 382. [Google Scholar] [CrossRef]
- Hildebrandt, E.; McGee, D.J. Helicobacter pylori Lipopolysaccharide Modification, Lewis Antigen Expression, and Gastric Colonization Are Cholesterol-Dependent. BMC Microbiol. 2009, 9, 258. [Google Scholar] [CrossRef]
- Ribeiro, F.C.; Rossoni, R.D.; de Barros, P.P.; Santos, J.D.; Fugisaki, L.R.O.; Leão, M.P.V.; Junqueira, J.C. Action Mechanisms of Probiotics on Candida Spp. and Candidiasis Prevention: An Update. J. Appl. Microbiol. 2020, 129, 175–185. [Google Scholar] [CrossRef]
- Fan, Y.; Chen, X.; Shan, T.; Wang, N.; Han, Q.; Ren, B.; Cheng, L. Polymicrobial Interactions of Helicobacter pylori and Its Role in the Process of Oral Diseases. J. Oral Microbiol. 2025, 17, 2469896. [Google Scholar] [CrossRef]
- Gareayaghi, N.; Akkus, S.; Saribas, S.; Demiryas, S.; Ozbey, D.; Kepil, N.; Demirci, M.; Dinc, H.O.; Akcin, R.; Uysal, O.; et al. Epstein-Barr Virus and Helicobacter pylori Co-Infection in Patients with Gastric Cancer and Duodenale Ulcer. New Microbiol. 2021, 44, 217–226. [Google Scholar]
- Shukla, S.K.; Prasad, K.N.; Tripathi, A.; Singh, A.; Saxena, A.; Chand Ghoshal, U.; Krishnani, N.; Husain, N.; Prasad, K.N. Epstein-Barr Virus DNA Load and Its Association with Helicobacter pylori Infection in Gastroduodenal Diseases. Braz. J. Infect. Dis. 2011, 15, 583–590. [Google Scholar] [CrossRef]
- Akkus, S.; Gareayaghi, N.; Saribas, S.; Demiryas, S.; Ozbey, D.; Kepil, N.; Demirci, M.; Ziver Sarp, T.; Oyku Dinc, H.; Akcin, R.; et al. Co-Infection Relationship with Epstein-Barr Virus in Gastroduodenal Diseases with Helicobacter pylori. Quantitative PCR and EBNA-1 Gene-Based Approach. Acta Gastroenterol. Belg. 2022, 85, 301–308. [Google Scholar] [CrossRef]
| Region | Worldwide Prevalence of H. pylori Antibiotic Resistance (%) | ||||
|---|---|---|---|---|---|
| Clarithromycin | Metronidazole | Levofloxacin | Amoxicillin | Tetracycline | |
| Africa | 15 | 91 | 14 | 38 | 13 |
| Americas | 14 | 27 | 14 | 8 | 4 |
| Eastern Mediterranean region | 29 | 61 | 23 | 14 | 10 |
| European region | 32 | 38 | 14 | 0 | 0 |
| Southeast Asia | 17 | 59 | 25 | 12 | 0 |
| Western Pacific region | 34 | 55 | 24 | 1 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braksator, J.; Kofla-Dłubacz, A.; Antosz-Popiołek, K.; Szyller, H.; Koga-Batko, J.; Wrześniewska, M.; Dyda, M.; Pytrus, T. Co-Occurrence of Helicobacter pylori and Candida spp. Infections in the Pathogenesis of Gastrointestinal Diseases. Biomedicines 2025, 13, 1172. https://doi.org/10.3390/biomedicines13051172
Braksator J, Kofla-Dłubacz A, Antosz-Popiołek K, Szyller H, Koga-Batko J, Wrześniewska M, Dyda M, Pytrus T. Co-Occurrence of Helicobacter pylori and Candida spp. Infections in the Pathogenesis of Gastrointestinal Diseases. Biomedicines. 2025; 13(5):1172. https://doi.org/10.3390/biomedicines13051172
Chicago/Turabian StyleBraksator, Joanna, Anna Kofla-Dłubacz, Katarzyna Antosz-Popiołek, Hubert Szyller, Joanna Koga-Batko, Martyna Wrześniewska, Maciej Dyda, and Tomasz Pytrus. 2025. "Co-Occurrence of Helicobacter pylori and Candida spp. Infections in the Pathogenesis of Gastrointestinal Diseases" Biomedicines 13, no. 5: 1172. https://doi.org/10.3390/biomedicines13051172
APA StyleBraksator, J., Kofla-Dłubacz, A., Antosz-Popiołek, K., Szyller, H., Koga-Batko, J., Wrześniewska, M., Dyda, M., & Pytrus, T. (2025). Co-Occurrence of Helicobacter pylori and Candida spp. Infections in the Pathogenesis of Gastrointestinal Diseases. Biomedicines, 13(5), 1172. https://doi.org/10.3390/biomedicines13051172