

MiR-27b-3p Reduces the Efficacy of Propranolol in the Treatment of Infantile Hemangioma by Inhibiting the Expression of Apaf-1

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatment
2.2. Cell Transfection
2.3. Cell Counting Kit-8
2.4. Flow Cytometry
2.5. TUNEL Staining
2.6. RNA Extraction and Quantitative RT-PCR
2.7. Western Blotting
2.8. Luciferase Reporter Assay
2.9. Statistical Analysis
3. Results
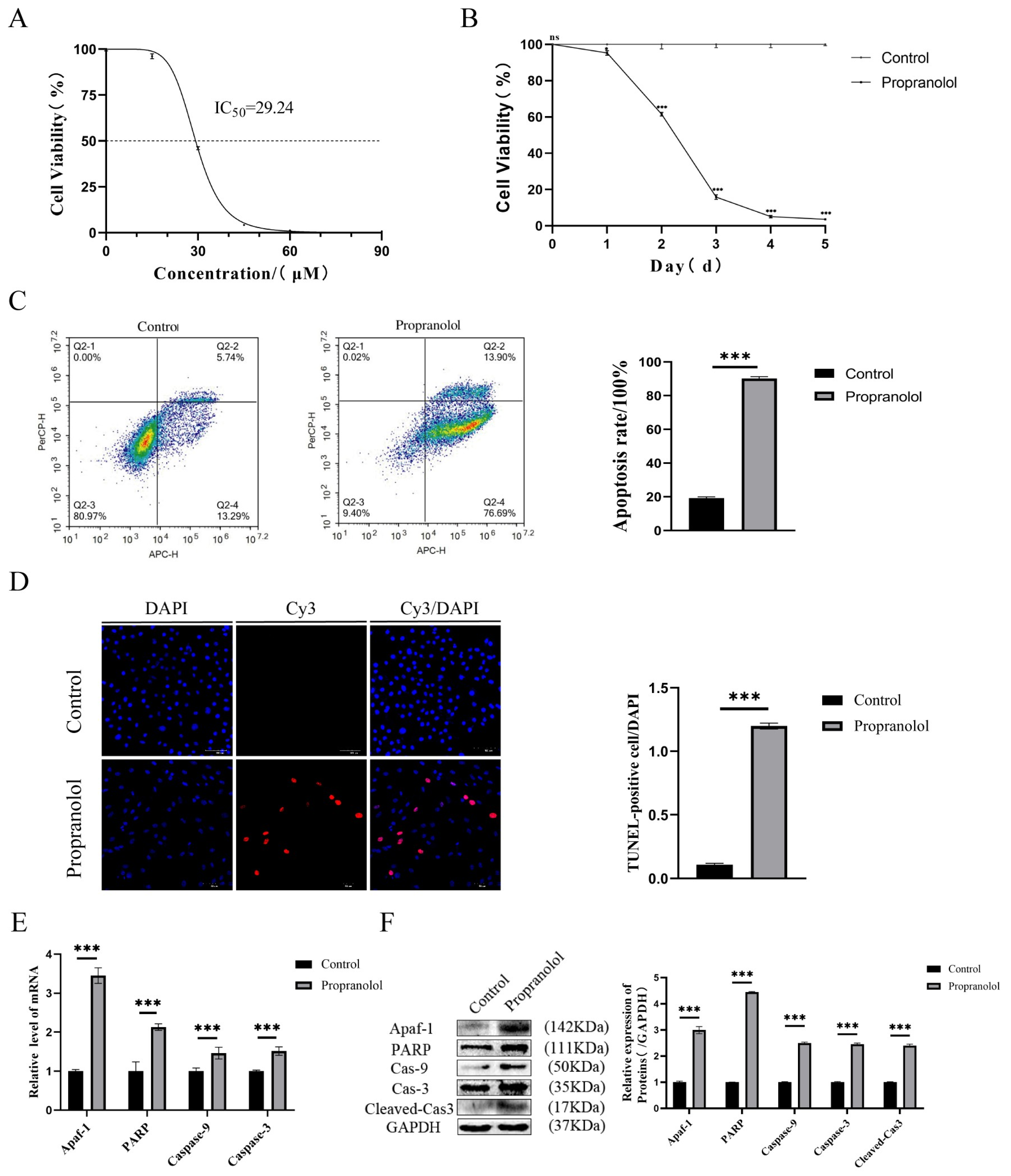
3.1. Propranolol Effectively Inhibited the Activity of HUVECs and Promoted Apoptosis In Vitro
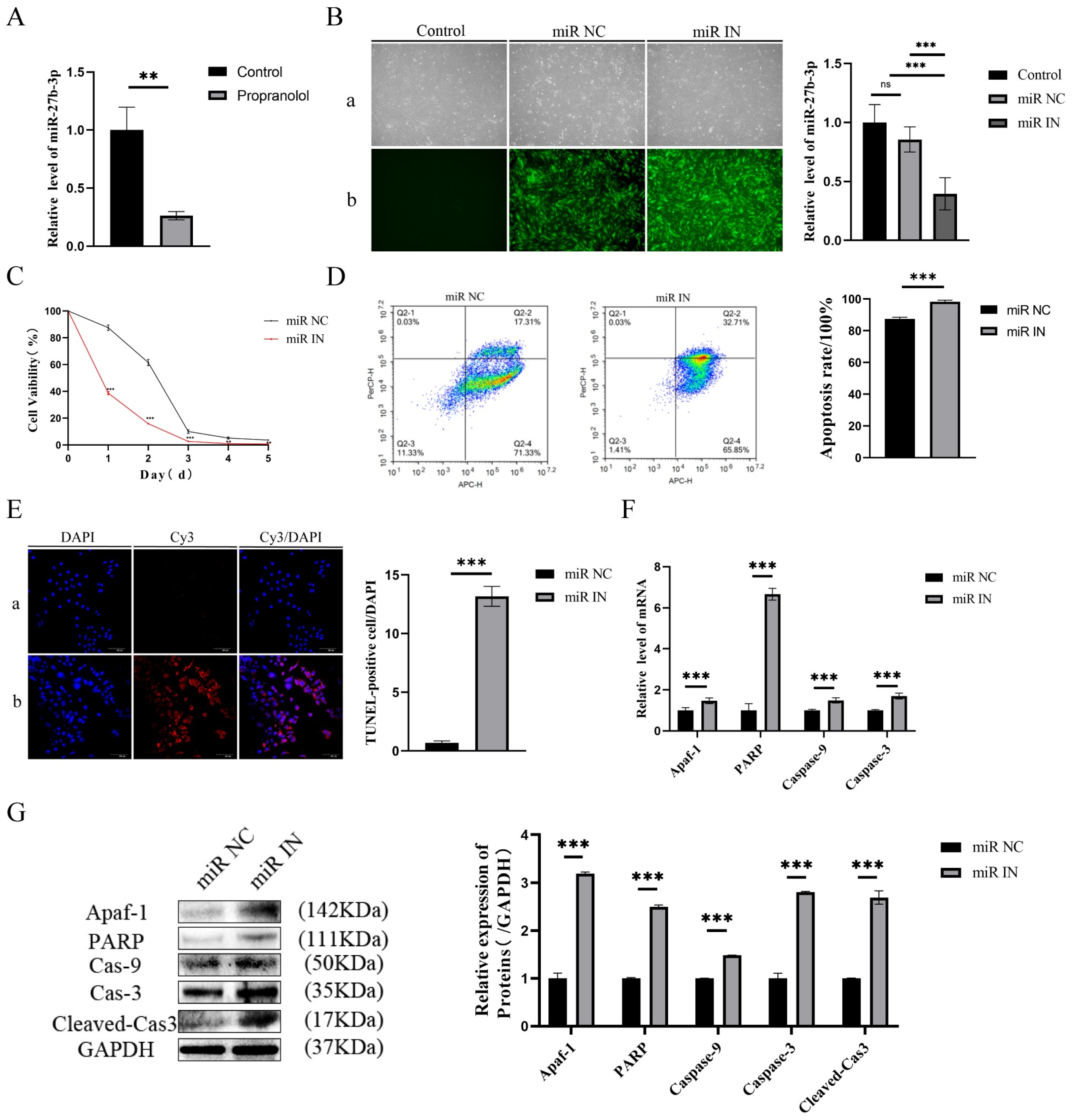
3.2. Propranolol Promotes HUVEC Apoptosis via Downregulating miR-27b-3p
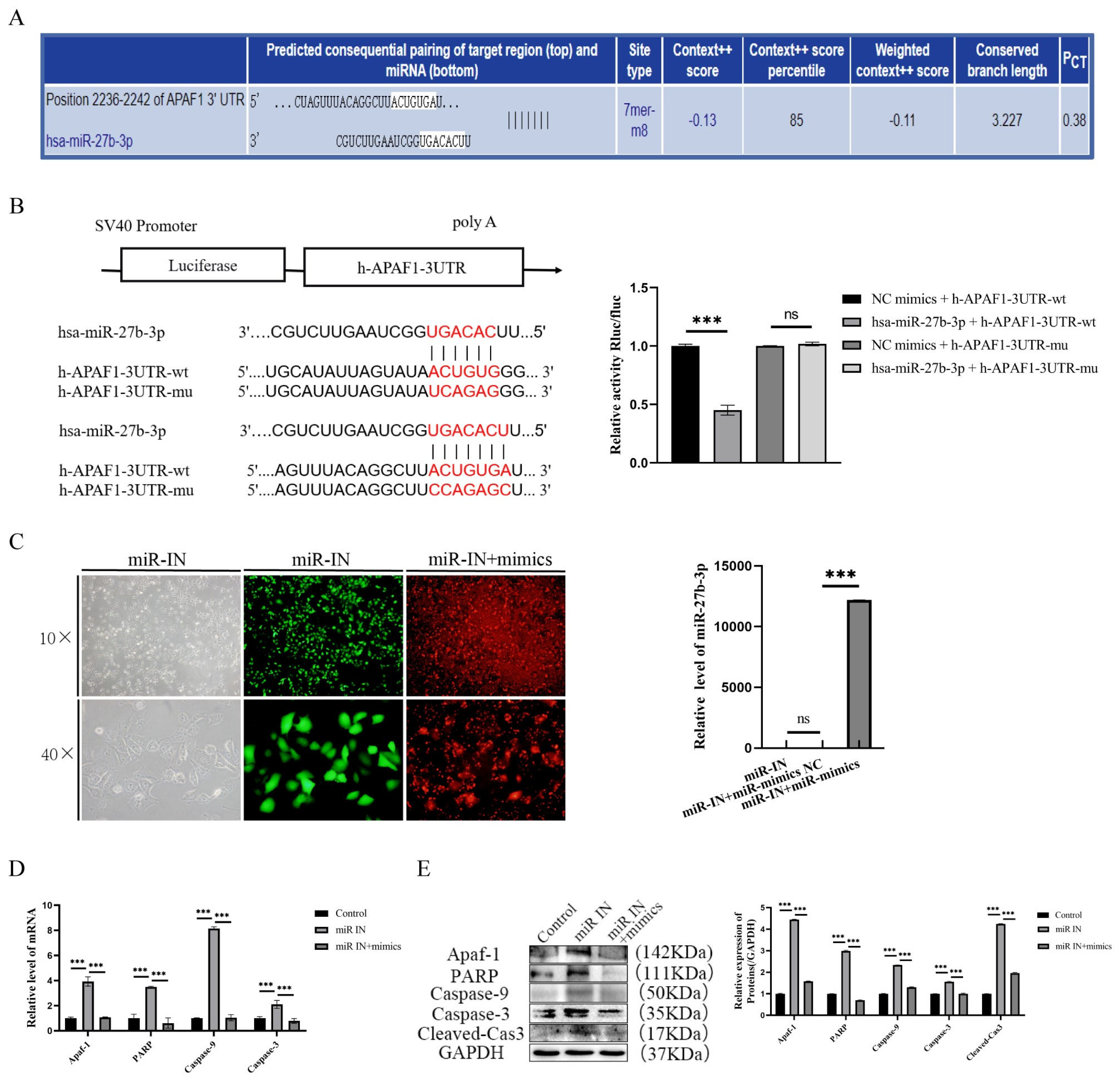
3.3. Targeted Inhibition of Apaf-1 by miR-27b-3p Reduces Propranolol-Induced Apoptosis in HUVECs
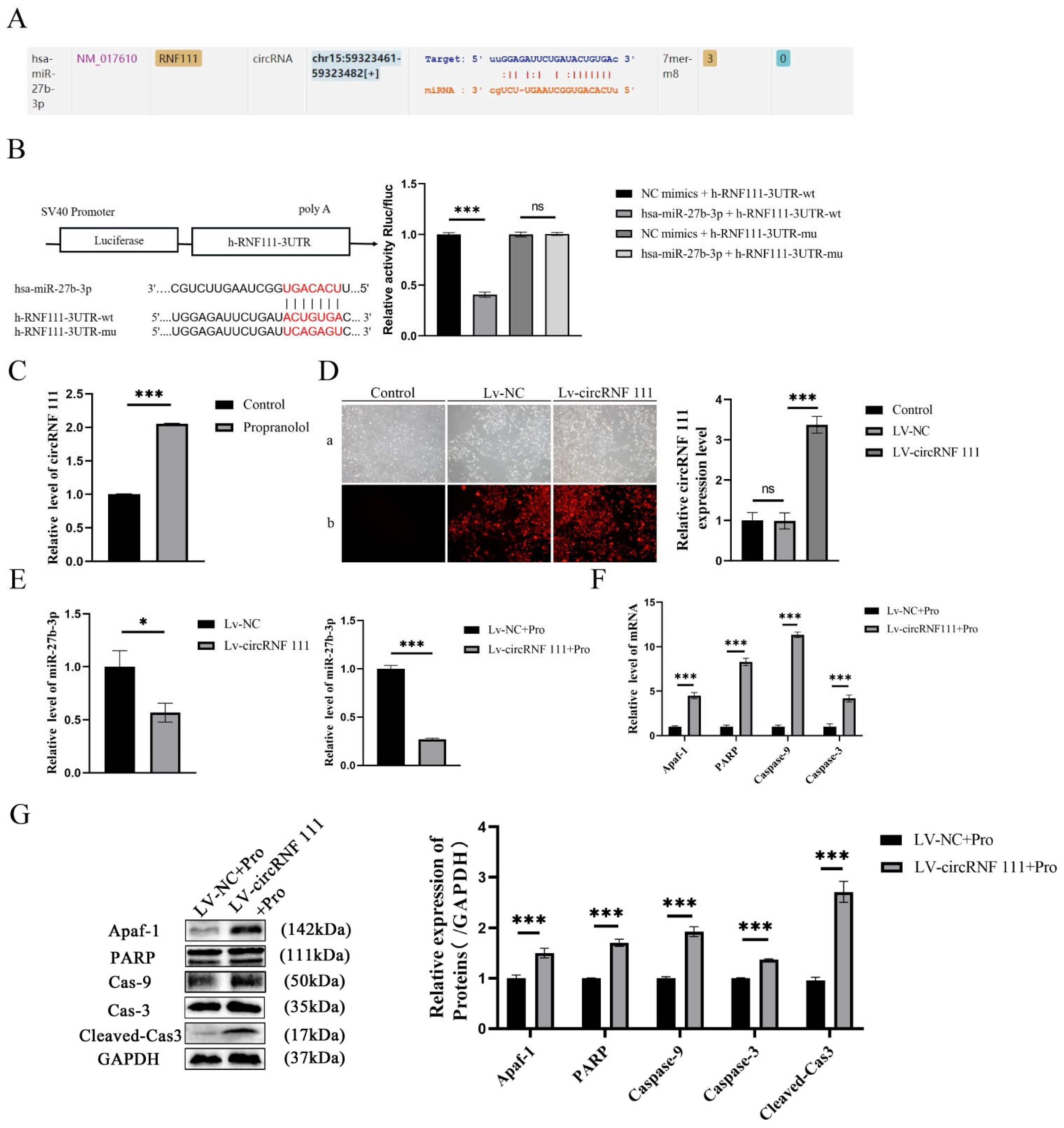
3.4. Propranolol Upregulates circRNF 111 to Promote HUVEC Apoptosis via miR-27b-3p/Apaf-1
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, C.; Zhao, Z.; Ji, Z.; Jiang, Y.; Zheng, J. MiR-187-3p Enhances Propranolol Sensitivity of Hemangioma Stem Cells. Cell Struct. Funct. 2019, 44, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Kum, J.J.; Khan, Z.A. Mechanisms of propranolol action in infantile hemangioma. Dermato-Endocrinology 2014, 6, e979699. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Gong, X.; Qiu, T.; Zhou, J.; Yang, K.; Lan, Y.; Zhang, Z.; Ji, Y. Insights into the mechanisms of angiogenesis in infantile hemangioma. Biomed. Pharmacother. 2024, 178, 117181. [Google Scholar] [CrossRef]
- Leaute-Labreze, C.; de la Roque, E.D.; Hubiche, T.; Boralevi, F.; Thambo, J.-B.; Taieb, A. Propranolol for severe hemangiomas of infancy. N. Engl. J. Med. 2008, 358, 2649–2651. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Chen, S.; Xu, C.; Li, L.; Xiang, B. The use of propranolol in the treatment of infantile haemangiomas: An update on potential mechanisms of action. Br. J. Dermatol. 2015, 172, 24–32. [Google Scholar] [CrossRef]
- Holm, A.; Mulliken, J.B.; Bischoff, J. Infantile hemangioma: The common and enigmatic vascular tumor. J. Clin. Investig. 2024, 134, e172836. [Google Scholar] [CrossRef]
- Tu, J.-B.; Ma, R.-Z.; Dong, Q.; Jiang, F.; Hu, X.-Y.; Li, Q.-Y.; Pattar, P.; Zhang, H. Induction of apoptosis in infantile hemangioma endothelial cells by propranolol. Exp. Ther. Med. 2013, 6, 574–578. [Google Scholar] [CrossRef]
- Ji, Y.; Li, K.; Xiao, X.; Zheng, S.; Xu, T.; Chen, S. Effects of propranolol on the proliferation and apoptosis of hemangioma-derived endothelial cells. J. Pediatr. Surg. 2012, 47, 2216–2223. [Google Scholar] [CrossRef]
- Yao, T.-H.; Pataer, P.; Regmi, K.P.; Gu, X.-W.; Li, Q.-Y.; Du, J.-T.; Ge, S.-M.; Tu, J.-B. Propranolol induces hemangioma endothelial cell apoptosis via a p53-BAX mediated pathway. Mol. Med. Rep. 2018, 18, 684–694. [Google Scholar] [CrossRef]
- Noori, A.R.; Hosseini, E.S.; Nikkhah, M.; Hosseinkhani, S. Apoptosome formation upon overexpression of native and truncated Apaf-1 in cell-free and cell-based systems. Arch. Biochem. Biophys. 2018, 642, 46–51. [Google Scholar] [CrossRef]
- Shakeri, R.; Kheirollahi, A.; Davoodi, J. Contribution of Apaf-1 to the pathogenesis of cancer and neurodegenerative diseases. Biochimie 2021, 190, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Grillone, K.; Carida, G.; Luciano, F.; Cordua, A.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. A systematic review of non-coding RNA therapeutics in early clinical trials: A new perspective against cancer. J. Transl. Med. 2024, 22, 731. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, N.; Pereira, L.M.S.; Severino, F.E.; Moura, R.; Yoshida, W.B.; Reis, P.P. Integrative meta-analysis identifies microRNA-regulated networks in infantile hemangioma. BMC Med. Genet. 2016, 17, 4. [Google Scholar] [CrossRef]
- Fish, J.E.; Srivastava, D. MicroRNAs: Opening a New Vein in Angiogenesis Research. Sci. Signal. 2009, 2, pe1. [Google Scholar] [CrossRef]
- Nakashima, T.; Jinnin, M.; Etoh, T.; Fukushima, S.; Masuguchi, S.; Maruo, K.; Inoue, Y.; Ishihara, T.; Ihn, H. Down-Regulation of mir-424 Contributes to the Abnormal Angiogenesis via MEK1 and Cyclin E1 in Senile Hemangioma: Its Implications to Therapy. PLoS ONE 2010, 5, e14334. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, P.; Liang, P.; Huang, X. The expression of miR-125b regulates angiogenesis during the recovery of heat-denatured HUVECs. Burns 2015, 41, 803–811. [Google Scholar] [CrossRef]
- Zeng, Z.; Chen, H.; Cai, J.; Huang, Y.; Yue, J. IL-10 regulates the malignancy of hemangioma-derived endothelial cells via regulation of PCNA. Arch. Biochem. Biophys. 2020, 688, 108404. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Sun, Y.; Shan, H.; Li, X.; Zhang, M.; Zhou, X.; Xing, S.; Sun, H.; Chu, W.; Qiao, G.; et al. Beta-adrenoceptor regulates miRNA expression in rat heart. Med. Sci. Monit. 2012, 18, BR309–BR314. [Google Scholar]
- Varela, M.A.; Roberts, T.C.; Wood, M.J.A. Epigenetics and ncRNAs in Brain Function and Disease: Mechanisms and Prospects for Therapy. Neurotherapeutics 2013, 10, 621–631. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, J.; Li, L.; Li, H.; Mao, S.; Zhang, F.; Zen, K.; Zhang, C.Y.; Zhang, Q. MicroRNA-23a/b and microRNA-27a/b suppress Apaf-1 protein and alleviate hypoxia-induced neuronal apoptosis. Cell Death Dis. 2014, 5, e1132. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jiang, Z.; Zhou, J.; Liu, Z. circRNA RNF111 regulates the growth, migration and invasion of gastric cancer cells by binding to miR-27b-3p. Int. J. Mol. Med. 2020, 46, 1873–1885. [Google Scholar] [CrossRef]
- Albinana, V.; Gallardo-Vara, E.; Casado-Vela, J.; Recio-Poveda, L.; Botella, L.M.; Cuesta, A.M. Propranolol: A “Pick and Roll” Team Player in Benign Tumors and Cancer Therapies. J. Clin. Med. 2022, 11, 4539. [Google Scholar] [CrossRef]
- Sang, Y.; Chen, B.; Song, X.; Li, Y.; Liang, Y.; Han, D.; Zhang, N.; Zhang, H.; Liu, Y.; Chen, T.; et al. circRNA_0025202 Regulates Tamoxifen Sensitivity and Tumor Progression via Regulating the miR-182-5p/FOXO3a Axis in Breast Cancer. Mol. Ther. 2019, 27, 1638–1652. [Google Scholar] [CrossRef]
- Tang, Q.; Chen, Z.; Zhao, L.; Xu, H. Circular RNA hsa_circ_0000515 acts as a miR-326 sponge to promote cervical cancer progression through up-regulation of ELK1. Aging 2019, 11, 9982–9999. [Google Scholar] [CrossRef]
- Wu, G.; Zhang, A.; Yang, Y.; Wu, D. Circ-RNF111 aggravates the malignancy of gastric cancer through miR-876-3p-dependent regulation of KLF12. World J. Surg. Oncol. 2021, 19, 259. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Xu, Y.; Wei, Z.; Ding, Q. CircAP2A2 acts as a ceRNA to participate in infantile hemangiomas progression by sponging miR-382-5p via regulating the expression of VEGFA. J. Clin. Lab. Anal. 2020, 34, e23258. [Google Scholar] [CrossRef]
- Sun, X.; Liu, B.; Zhao, X.-D.; Wang, L.-Y.; Ji, W.-Y. MicroRNA-221 accelerates the proliferation of laryngeal cancer cell line Hep-2 by suppressing Apaf-1. Oncol. Rep. 2015, 33, 1221–1226. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Fisslthaler, B.; Zippel, N.; Froemel, T.; Hu, J.; Elgheznawy, A.; Heide, H.; Popp, R.; Fleming, I. MicroRNA-223 Antagonizes Angiogenesis by Targeting β1 Integrin and Preventing Growth Factor Signaling in Endothelial Cells. Circ. Res. 2013, 113, 1320–1330. [Google Scholar] [CrossRef]
- Shen, S.-J.; Song, Y.; Ren, X.-Y.; Xu, Y.-L.; Zhou, Y.-D.; Liang, Z.-Y.; Sun, Q. MicroRNA-27b-3p Promotes Tumor Progression and Metastasis by Inhibiting Peroxisome Proliferator-Activated Receptor Gamma in Triple-Negative Breast Cancer. Front. Oncol. 2020, 10, 1371. [Google Scholar] [CrossRef]
- Zhang, Q.; Shao, W.; Li, Y.; Liu, L.; Chen, W.; Wang, C.; Li, B. Long non-coding RNA LINC01128 affects proliferation, migration, and invasion of glioma cells by regulating miR-27b-3p. Folia Neuropathol. 2022, 60, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Huang, S.; Cao, J.; Ye, J.; Huang, H.; Liao, D.; Yang, Y.; Chen, W.; Pu, R. Combined targeting of vascular endothelial growth factor C (VEGFC) and P65 using miR-27b-3p agomir and lipoteichoic acid in the treatment of gastric cancer. J. Gastrointest. Oncol. 2021, 12, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Liu, Z.; Xia, J. New insights into circRNA and its mechanisms in angiogenesis regulation in ischemic stroke: A biomarker and therapeutic target. Mol. Biol. Rep. 2023, 50, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhao, Y.; Chen, Q.; Zhu, S.; Niu, Y.; Ye, Z.; Hu, P.; Chen, D.; Xu, P.; Chen, J.; et al. Hypoxic exosomal HIF-1α-stabilizing circZNF91 promotes chemoresistance of normoxic pancreatic cancer cells via enhancing glycolysis. Oncogene 2021, 40, 5505–5517. [Google Scholar] [CrossRef]
- Gomez-Acevedo, H.; Dai, Y.; Strub, G.; Shawber, C.; Wu, J.K.; Richter, G.T. Identification of putative biomarkers for Infantile Hemangiomas and Propranolol treatment via data integration. Sci. Rep. 2020, 10, 3261. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Name of the Gene | Primer Sequence |
|---|---|
| miRNA-27b-3P circRNF 111 | 5′-AGTCCCGTGTCTGTAATGCC-3′ F: 5′-TAGCAGTTCCCCAATCCTTG-3′ R: 5′-CACAAATTCCCATCATTCCC-3′ |
| Apaf-1 | F: 5′-CGGCCCTGCGCATCTGATTCAT-3′ R: 5′-GGGCGAACGACTAAGCGGGACAG-3′ |
| PARP | F: 5′-GGTCTTCCCCTACCCTCTCAA-3′ R: 5′-CGTTGTGTGTTCGCCTCT-3′ |
| Caspase-9 | F: 5′-CTGAGCCAGATGCTGTCCCATA-3′ R: 5′-GACACCATCCAAGGTCTCGATGTA-3′ |
| Caspase-3 U6 | F: 5′-GACTGCGGTATTGAGACAGA-3′ R: 5′-CGAGTGAGGATGTGCATGAA-3′ F: 5′-CTCGCTTCGGCAGCACATATACTA-3′ R: 5′-ACGAATTTGCGTGTCATCCTTGCG-3′ |
| GAPDH | F: 5′-TGTTCGTCATGGGTGTGAACC-3′ R: 5′-ATGGACTGTGGTCATGAGTCC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhu, Y.; Xie, L.; Ahmadi, S.; Yao, C.; Cui, H.; Kang, X.; Tu, J.; Na, S. MiR-27b-3p Reduces the Efficacy of Propranolol in the Treatment of Infantile Hemangioma by Inhibiting the Expression of Apaf-1. Biomedicines 2025, 13, 1092. https://doi.org/10.3390/biomedicines13051092
Li J, Zhu Y, Xie L, Ahmadi S, Yao C, Cui H, Kang X, Tu J, Na S. MiR-27b-3p Reduces the Efficacy of Propranolol in the Treatment of Infantile Hemangioma by Inhibiting the Expression of Apaf-1. Biomedicines. 2025; 13(5):1092. https://doi.org/10.3390/biomedicines13051092
Chicago/Turabian StyleLi, Jing, Yifei Zhu, Linyang Xie, Sina Ahmadi, Chonghao Yao, Hao Cui, Xuteng Kang, Junbo Tu, and Sijia Na. 2025. "MiR-27b-3p Reduces the Efficacy of Propranolol in the Treatment of Infantile Hemangioma by Inhibiting the Expression of Apaf-1" Biomedicines 13, no. 5: 1092. https://doi.org/10.3390/biomedicines13051092
APA StyleLi, J., Zhu, Y., Xie, L., Ahmadi, S., Yao, C., Cui, H., Kang, X., Tu, J., & Na, S. (2025). MiR-27b-3p Reduces the Efficacy of Propranolol in the Treatment of Infantile Hemangioma by Inhibiting the Expression of Apaf-1. Biomedicines, 13(5), 1092. https://doi.org/10.3390/biomedicines13051092