
The Hormetic Adaptative Capacity and Resilience to Oxidative Stress Is Strengthened by Exposome Enrichment with Air Cold Atmospheric Plasma: A Metabolome Targeted Follow-Up Approach


 , , ,
, , ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Air Cold Atmospheric Plasma–Nanoparticle Removal (aCAP-NR) Device
2.3. Sample Preparation and Metabolomic Analysis
2.4. Statistical and Pathway Analysis
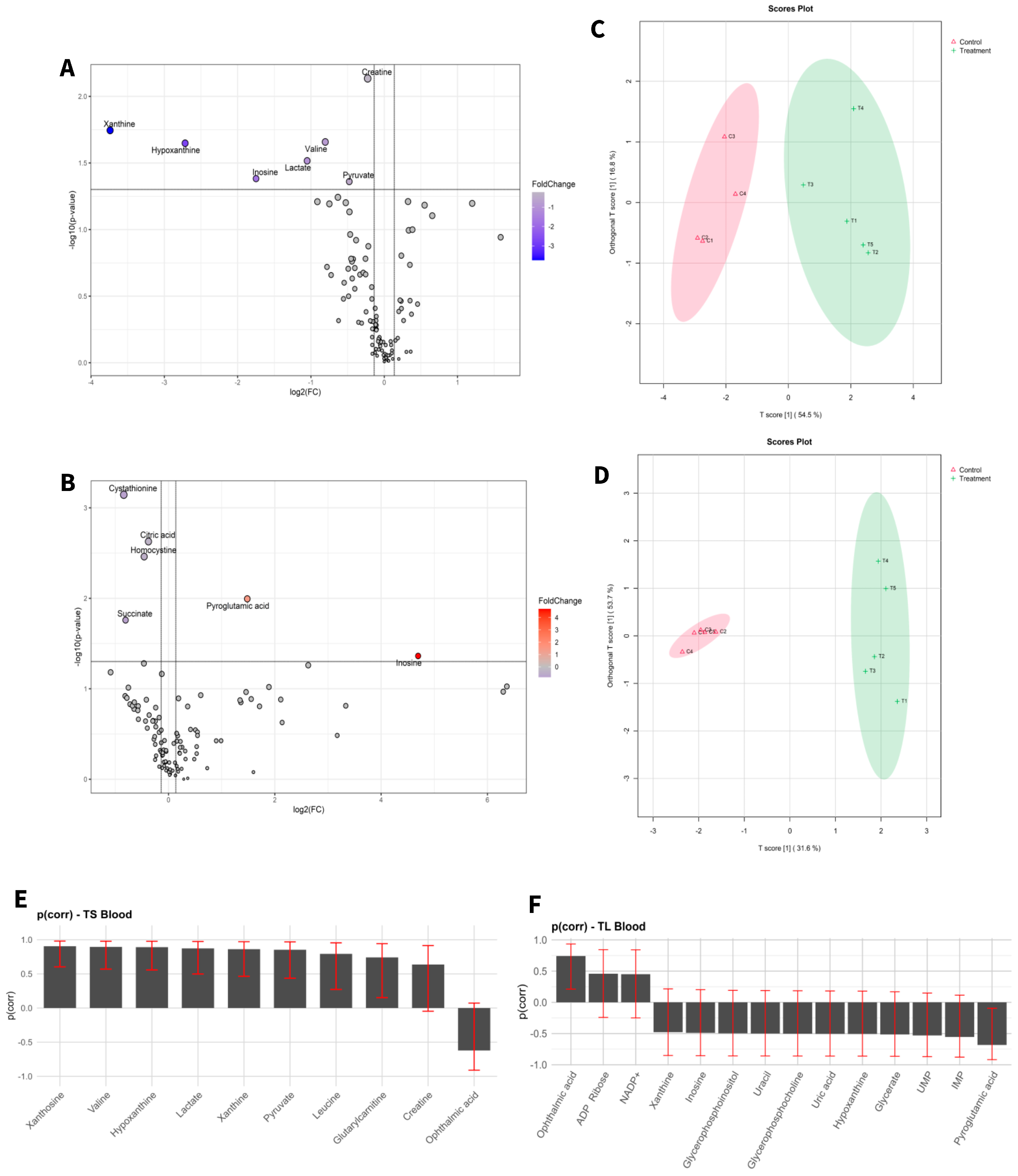
3. Results
3.1. Changes in Metabolome Landscape
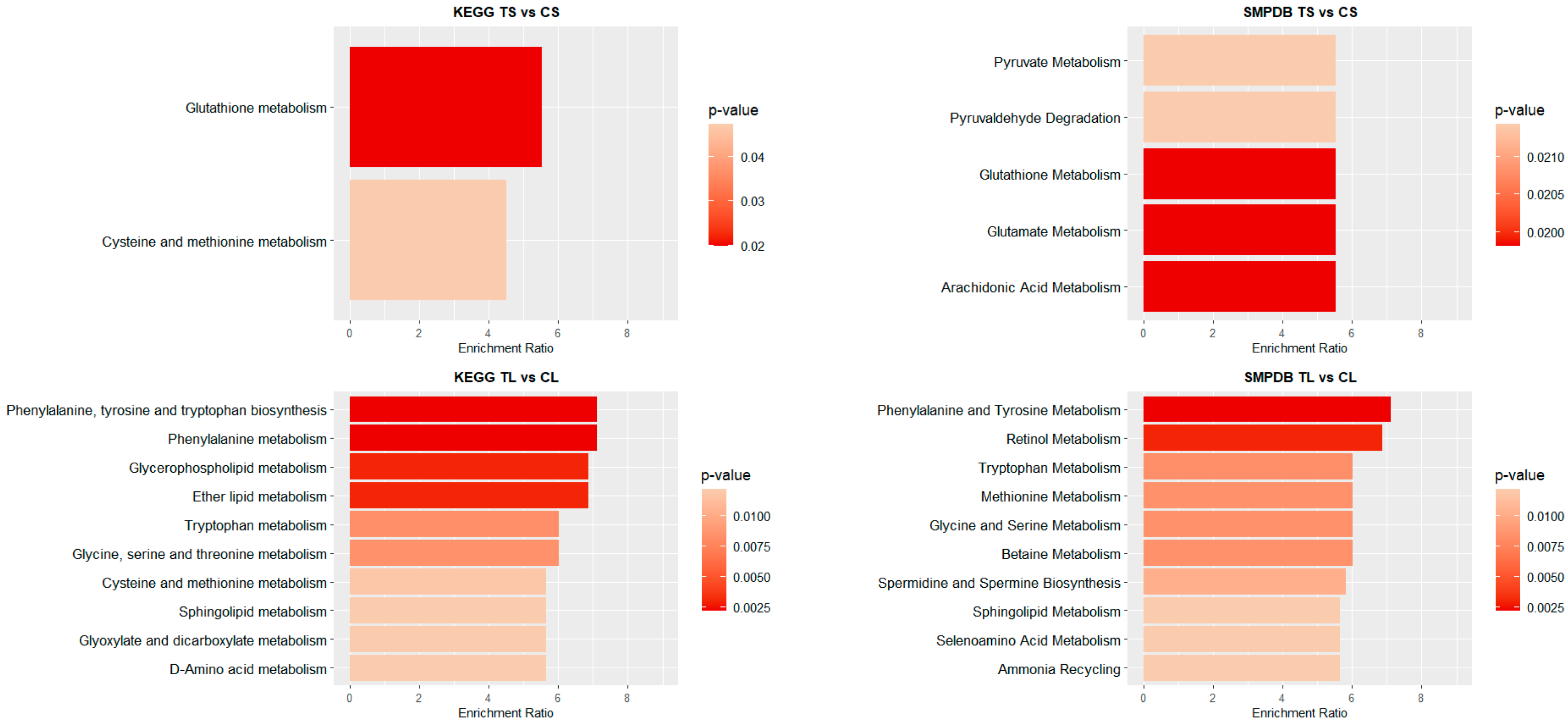
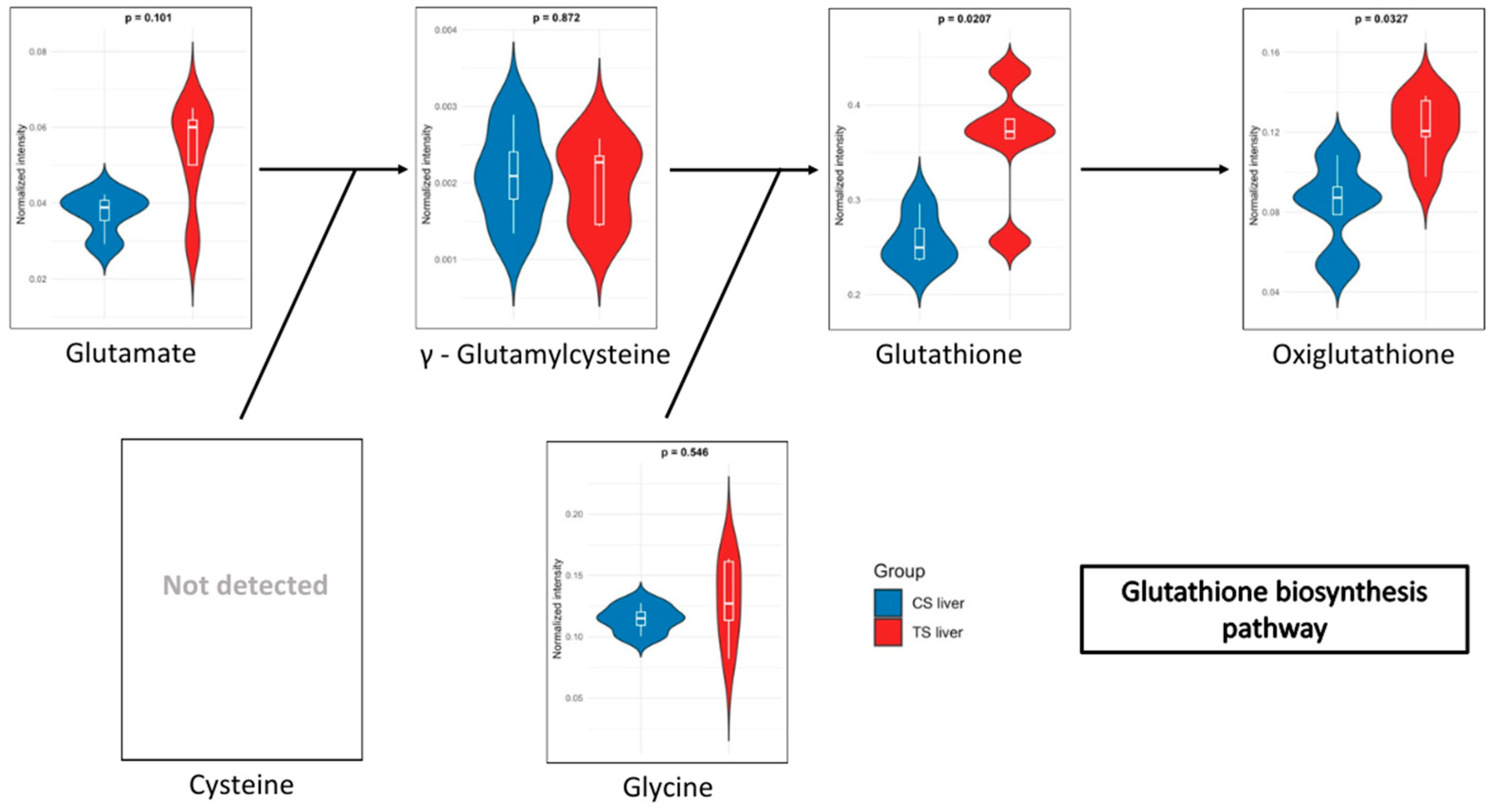
3.2. Metabolome Pathway Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Environmental Health. Available online: https://www.who.int/health-topics/environmental-health (accessed on 24 October 2024).
- European Commission. Directorate-General for Research and Innovation, Horizon Europe: Strategic Plan 2021–2024; Publications Office of the European Union: Luxembourg, 2021. [Google Scholar]
- Misra, B.B. The Chemical Exposome of Human Aging. Front. Genet. 2020, 11, 574936. [Google Scholar] [CrossRef] [PubMed]
- Antuña, E.; Carlos Bermejo-Millo, J.; Caso-Onzain, E.; Caso-Peláez, E.; Potes, Y.; Coto-Montes, A. Removal of Environmental Nanoparticles Increases Protein Synthesis and Energy Production in Healthy Humans. Front. Bioeng. Biotechnol. 2022, 10, 800011. [Google Scholar] [CrossRef]
- Xiao, S.; Wei, T.; Petersen, J.D.; Zhou, J.; Lu, X. Biological effects of negative air ions on human health and integrated multiomics to identify biomarkers: A literature review. Environ. Sci. Pollut. Res. Int. 2023, 30, 69824–69836. [Google Scholar] [CrossRef]
- Liu, S.; Huang, Q.; Wu, Y.; Song, Y.; Dong, W.; Chu, M.; Yang, D.; Zhang, X.; Zhang, J.; Chen, C.; et al. Metabolic linkages between indoor negative air ions, particulate matter and cardiorespiratory function: A randomized, double-blind crossover study among children. Environ. Int. 2020, 138, 105663, Erratum in Environ. Int. 2020, 143, 106073. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, C.; Kouretas, D. Lipid peroxidation and tissue damage. In Vivo 1999, 13, 295–309. [Google Scholar]
- Ito, F.; Sono, Y.; Ito, T. Measurement and clinical significance of lipid peroxidation as a biomarker of oxidative stress: Oxidative stress in diabetes, atherosclerosis, and chronic inflammation. Antioxidants 2019, 8, 72. [Google Scholar] [CrossRef]
- Bhattacharjee, B.; Bezbaruah, R.; Rynjah, D.; Newar, A.; Sengupta, S.; Pegu, P.; Dey, N.; Bora, S.C.; Barman, D. Cold atmospheric plasma: A noteworthy approach in medical science. Sci. Phar. 2023, 2, 46–76. [Google Scholar] [CrossRef]
- Collin, F. Chemical basis of Reactive Oxygen Species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef]
- Zuo, L.; Wijegunawardana, D. Redox role of ROS and inflammation in pulmonary diseases. Adv. Exp. Med. Biol. 2021, 1304, 187–204. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, X.; Xu, W.; Li, J.; Sun, Y.; Cui, S.; Xu, R.; Li, W.; Jiao, L.; Wang, T. ROS-induced endothelial dysfunction in the pathogenesis of atherosclerosis. Aging Dis. 2024, 16, 250–268. [Google Scholar] [CrossRef] [PubMed]
- Izadjoo, M.; Zack, S.; Kim, H.; Skiba, J. Medical applications of cold atmospheric plasma: State of the science. J. Wound Care 2018, 27 (Suppl. 9), S4–S10. [Google Scholar] [CrossRef] [PubMed]
- Raja, S.J.; Akhildas, S.; Nirmal, M.R.; Rajan, L.; Anand, V. Experimental characterization of atmospheric-pressure plasma using Langmuir probe techniques. In Proceedings of the 2024 IEEE Second International Conference on Emerging Trends in Information Technology and Engineering (ICETITE), Vellore, India, 22–23 February 2024; pp. 1–8. [Google Scholar]
- Cheng, F.; Yan, D.; Chen, J.; Wang, Z.; Horkowitz, A.; Keidar, M.; Sotomayor, E.M. Enhancing innate and adaptive immune systems by cold atmospheric plasma (CAP) and its antitumor immunity. arXiv 2022, arXiv:2201.12737. [Google Scholar]
- Biazar, E.; Aavani, F.; Zeinali, R.; Kheilnezhad, B.; Taheri, K.; Yahyaei, Z. Delivery systems for plasma-reactive species and their applications in the field of biomedicine. Curr. Drug Deliv. 2024, 21, 1497–1514. [Google Scholar] [CrossRef]
- Gross, T.; Ledernez, L.A.; Birrer, L.; Bergmann, M.E.; Altenburger, M.J. Guided plasma application in dentistry-an alternative to antibiotic therapy. Antibiotics 2024, 13, 735. [Google Scholar] [CrossRef]
- Braný, D.; Dvorská, D.; Halašová, E.; Škovierová, H. Cold atmospheric plasma: A powerful tool for modern medicine. Int. J. Mol. Sci. 2020, 21, 2932. [Google Scholar] [CrossRef]
- Menéndez-Coto, N.; Garcia-Gonzalez, C.; Baena-Huerta, F.J.; Zapata-Pérez, R.; Rabadán-Ros, R.; Núñez-Delicado, E.; González-Llorente, L.; Caso-Peláez, E.; Coto-Montes, A. Combining cold Atmospheric Plasma and environmental nanoparticle removal device reduces neurodegenerative markers. Int. J. Mol. Sci. 2024, 25, 12986. [Google Scholar] [CrossRef]
- Saifutdinova, A.A.; Saifutdinov, A.I.; Gainullina, S.V.; Timerkaev, B.A. Modeling the Parameters of an Atmospheric Pressure Dielectric Barrier Discharge Controlled by the Shape of the Applied Voltage. IEEE Trans. Plasma Sci. 2022, 50, 1144–1156. [Google Scholar] [CrossRef]
- Frolova, A.A.; Piskunov, A.S. Numerical modeling of a cold plasma jet with control of discharge parameters. Processes 2023, 13, 632. [Google Scholar] [CrossRef]
- Brandenburg, R. Dielectric barrier discharges: Progress on plasma sources and on the understanding of regimes and single filaments. Plasma Sources Sci. Technol. 2017, 26, 053001. [Google Scholar] [CrossRef]
- Lin, L.; Keidar, M. A map of control for cold atmospheric plasma jets. APL Rev. 2021, 8, 011306. [Google Scholar] [CrossRef]
- Girard, F.; Peret, M.; Dumont, N.; Badets, V.; Blanc, S.; Gazeli, K.; Noël, C.; Belmonte, T.; Marlin, L.; Cambus, J.-P.; et al. Correlations between gaseous and liquid phase chemistries induced by cold atmospheric plasmas in a physiological buffer. Phys. Chem. Chem. Phys. 2018, 20, 9198–9210. [Google Scholar] [CrossRef]
- Živanić, M.; Espona-Noguera, A.; Lin, A.; Canal, C. Current state of cold atmospheric plasma and cancer-immunity cycle: Therapeutic relevance and overcoming clinical limitations using hydrogels. Adv. Sci. 2023, 10, e2205803. [Google Scholar] [CrossRef] [PubMed]
- Janda, M.; Stancampiano, A.; di Natale, F.; Machala, Z. Short Review on Plasma–Aerosol Interactions. Plasma Process. Polym. 2025, 22, e2400275. [Google Scholar] [CrossRef]
- Meinke, M.C.; Hasse, S.; Schleusener, J.; Hahn, V.; Gerling, T.; Hadian Rasnani, K.; Bernhardt, T.; Ficht, P.-K.; Staffeld, A.; Bekeschus, S.; et al. Radical formation in skin and preclinical characterization of a novel medical plasma device for dermatology after single application. Free Radic. Biol. Med. 2025, 226, 199–215. [Google Scholar] [CrossRef]
- Chauvin, J.; Judée, F.; Yousfi, M.; Vicendo, P.; Merbahi, N. Analysis of reactive oxygen and nitrogen species generated in three liquid media by low temperature helium plasma jet. Sci. Rep. 2017, 7, 4562. [Google Scholar] [CrossRef] [PubMed]
- Girard, P.-M.; Arbabian, A.; Fleury, M.; Bauville, G.; Puech, V.; Dutreix, M.; Sousa, J.S. Synergistic effect of H2O2 and NO2 in cell death induced by cold atmospheric He plasma. Sci. Rep. 2016, 6, 29098. [Google Scholar] [CrossRef]
- Girard-Sahun, F.; Lefrançois, P.; Badets, V.; Arbault, S.; Clement, F. Direct sensing of superoxide and its relatives reactive oxygen and nitrogen species in phosphate buffers during cold atmospheric plasmas exposures. Anal. Chem. 2022, 94, 5555–5565. [Google Scholar] [CrossRef]
- Jang, J.-Y.; Hong, Y.J.; Lim, J.; Choi, J.S.; Choi, E.H.; Kang, S.; Rhim, H. Cold atmospheric plasma (CAP), a novel physicochemical source, induces neural differentiation through cross-talk between the specific RONS cascade and Trk/Ras/ERK signaling pathway. Biomaterials 2018, 156, 258–273. [Google Scholar] [CrossRef]
- Xu, D.; Liu, D.; Wang, B.; Chen, C.; Chen, Z.; Li, D.; Yang, Y.; Chen, H.; Kong, M.G. In situ OH generation from O2− and H2O2 plays a critical role in plasma-induced cell death. PLoS ONE 2015, 10, e0128205. [Google Scholar] [CrossRef]
- Schmidt, A.; Bekeschus, S. Redox for repair: Cold physical plasmas and Nrf2 signaling promoting wound healing. Antioxidants 2018, 7, 146. [Google Scholar] [CrossRef] [PubMed]
- Skaperda, Z.; Tekos, F.; Vardakas, P.; Nepka, C.; Kouretas, D. Reconceptualization of hormetic responses in the frame of redox toxicology. Int. J. Mol. Sci. 2021, 23, 49. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Kozumbo, W.J. The hormetic dose-response mechanism: Nrf2 activation. Pharmacol. Res. 2021, 167, 105526. [Google Scholar] [CrossRef]
- SIST EN 60601-1-2:2015/A1:2021; Medical Electrical Equipment—Part 1–2: General Requirements for Basic Safety and Essential Performance—Collateral Standard: Electromagnetic Disturbances—Requirements and Tests (IEC 60601-1-2:2014/A1:2020). Available online: https://standards.iteh.ai/catalog/standards/sist/49ced698-56ab-4bb8-97e9-a6017a0ad6b4/sist-en-60601-1-2-2015-a1-2021?srsltid=AfmBOooDKvs5PkdRdP47T3SVVbxDWmW1uYsIN4dJUmxk5_0GZpcQnxWZ (accessed on 11 March 2025).
- SIST EN 60601-1:2007/A1:2014/AC:2015; Medical Electrical Equipment—Part 1: General Requirements for Basic Safety and Essential Performance. Available online: https://standards.iteh.ai/catalog/standards/sist/17cf8992-761d-4eca-943e-8eefa3d86fef/sist-en-60601-1-2007-a1-2014-ac-2015?srsltid=AfmBOopw8mgzHWtIAQmADVtQQvIRUnelAKOHDYO6-cKW2NczzUCgO0lK (accessed on 11 March 2025).
- Official Journal of the European Union, L 174, 1 July 2011. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=oj:JOL_2011_174_R_TOC (accessed on 11 March 2025).
- Lundberg, J.O.; Weitzberg, E. Biology of nitrogen oxides in the gastrointestinal tract. Gut 2013, 62, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.K.; Kumar, S.; Choi, E.-H.; Sharma, P.; Misra, S.; Kim, M.-H. Insight into the molecular dynamic simulation studies of reactive oxygen species in native skin membrane. Front. Pharmacol. 2018, 9, 644. [Google Scholar] [CrossRef]
- EN 60601-1; The European Version of the IEC 60601-1 Standard for Medical Electrical Equipment. Available online: https://www.iso.org/standard/59536.html#amendment (accessed on 11 March 2025).
- Ström, M.; Wagner, N.; Kolosenko, I.; Wheelock, Å.M. Enhancing the Robustness of OPLS Modelling in Small Cohorts by Leveraging Permutation Analysis Prior to Variable Selection bioRxiv. 2024. Available online: https://www.biorxiv.org/content/10.1101/2024.03.18.585475v1.full.pdf (accessed on 11 March 2025).
- Chong, J.; Xia, J. MetaboAnalystR: An R package for flexible and reproducible analysis of metabolomics data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef]
- Tian, L.; Li, Z.; Ma, G.; Zhang, X.; Tang, Z.; Wang, S.; Kang, J.; Liang, D.; Yu, T. Metapone: A Bioconductor package for joint pathway testing for untargeted metabolomics data. Bioinformatics 2022, 38, 3662–3664. [Google Scholar] [CrossRef] [PubMed]
- Jewison, T.; Su, Y.; Disfany, F.M.; Liang, Y.; Knox, C.; Maciejewski, A.; Poelzer, J.; Huynh, J.; Zhou, Y.; Arndt, D.; et al. SMPDB 2.0, big improvements to the Small Molecule Pathway Database. Nucleic Acids Res. 2014, 42, D478–D484. [Google Scholar] [CrossRef]
- Zhang, P.; Carlsten, C.; Chaleckis, R.; Hanhineva, K.; Huang, M.; Isobe, T.; Koistinen, V.M.; Meister, I.; Papazian, S.; Sdougkou, K.; et al. Defining the scope of exposome studies and research needs from a multidisciplinary perspective. Environ. Sci. Technol. Lett. 2021, 8, 839–852. [Google Scholar] [CrossRef]
- Hagn, G.; Meier-Menches, S.M.; Plessl-Walder, G.; Mitra, G.; Mohr, T.; Preindl, K.; Schlatter, A.; Schmidl, D.; Gerner, C.; Garhöfer, G.; et al. Plasma instead of serum avoids critical confounding of clinical metabolomics studies by platelets. J. Proteome Res. 2024, 23, 3064–3075. [Google Scholar] [CrossRef]
- Dickson, P.E.; Mittleman, G. Environmental enrichment influences novelty reactivity, novelty preference, and anxiety via distinct genetic mechanisms in C57BL/6J and DBA/2J mice. Sci. Rep. 2021, 11, 3928. [Google Scholar] [CrossRef] [PubMed]
- Argentieri, M.A.; Amin, N.; Nevado-Holgado, A.J.; Sproviero, W.; Collister, J.A.; Keestra, S.M.; Kuilman, M.M.; Ginos, B.N.R.; Ghanbari, M.; Doherty, A.; et al. Integrating the environmental and genetic architectures of aging and mortality. Nat. Med. 2025, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Maitre, L.; Guimbaud, J.-B.; Warembourg, C.; Güil-Oumrait, N.; Petrone, P.M.; Chadeau-Hyam, M.; Vrijheid, M.; Basagaña, X.; Gonzalez, J.R. State-of-the-art methods for exposure-health studies: Results from the exposome data challenge event. Environ. Int. 2022, 168, 107422. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Chen, S.; Jin, X.; Liu, S.; Cao, X.; Liu, B. Metabolomics in aging research: Aging markers from organs. Front. Cell Dev. Biol. 2023, 11, 1198794. [Google Scholar] [CrossRef]
- Zhang, K.; Hast, M.W.; Izumi, S.; Usami, Y.; Shetye, S.; Akabudike, N.; Philp, N.J.; Iwamoto, M.; Nissim, I.; Soslowsky, L.J.; et al. Modulating glucose metabolism and lactate synthesis in injured mouse tendons: Treatment with dichloroacetate, a lactate synthesis inhibitor, improves tendon healing. Am. J. Sports Med. 2018, 46, 2222–2231. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Rossiter, H.B. Each-step activation of oxidative phosphorylation is necessary to explain muscle metabolic kinetic responses to exercise and recovery in humans. J. Physiol. 2015, 593, 5255–5268. [Google Scholar] [CrossRef]
- Grassi, B.; Hogan, M.C.; Gladden, L.B. Microvascular O2 delivery and O2 utilization during metabolic transitions in skeletal muscle. One-hundred years after the pioneering work by August Krogh. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2020, 252, 110842. [Google Scholar] [CrossRef]
- Horikoshi, M.; Harada, K.; Tsuno, S.; Kitaguchi, T.; Hirai, M.Y.; Matsumoto, M.; Terada, S.; Tsuboi, T. Distinct lactate metabolism between hepatocytes and myotubes revealed by live cell imaging with genetically encoded indicators. Biochem. Biophys. Res. Commun. 2023, 694, 149416. [Google Scholar] [CrossRef]
- Martino, M.R.; Habibi, M.; Ferguson, D.; Brookheart, R.T.; Thyfault, J.P.; Meyer, G.A.; Lantier, L.; Hughey, C.C.; Finck, B.N. Disruption of hepatic mitochondrial pyruvate and amino acid metabolism impairs gluconeogenesis and endurance exercise capacity in mice. Am. J. Physiol. Endocrinol. Metab. 2024, 326, E515–E527. [Google Scholar] [CrossRef]
- Suhara, T.; Hishiki, T.; Kasahara, M.; Hayakawa, N.; Oyaizu, T.; Nakanishi, T.; Kubo, A.; Morisaki, H.; Kaelin, W.G.; Suematsu, M.; et al. Inhibition of the oxygen sensor PHD2 in the liver improves survival in lactic acidosis by activating the Cori cycle. Proc. Natl. Acad. Sci. USA 2015, 112, 11642–11647. [Google Scholar] [CrossRef]
- Sharer, J.D.; Bodamer, O.; Longo, N.; Tortorelli, S.; Wamelink, M.M.C.; Young, S. Laboratory diagnosis of creatine deficiency syndromes: A technical standard and guideline of the American College of Medical Genetics and Genomics. Genet. Med. 2017, 19, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Wax, B.; Kerksick, C.M.; Jagim, A.R.; Mayo, J.J.; Lyons, B.C.; Kreider, R.B. Creatine for exercise and sports performance, with recovery considerations for healthy populations. Nutrients 2021, 13, 1915. [Google Scholar] [CrossRef] [PubMed]
- Gawedzka, A.; Grandys, M.; Duda, K.; Zapart-Bukowska, J.; Zoladz, J.A.; Majerczak, J. Plasma BCAA concentrations during exercise of varied intensities in young healthy men-the impact of endurance training. PeerJ 2020, 8, e10491. [Google Scholar] [CrossRef] [PubMed]
- Areces, F.; González-Millán, C.; Salinero, J.J.; Abian-Vicen, J.; Lara, B.; Gallo-Salazar, C.; Ruiz-Vicente, D.; Del Coso, J. Changes in serum free amino acids and muscle fatigue experienced during a half-Ironman triathlon. PLoS ONE 2015, 10, e0138376. [Google Scholar] [CrossRef]
- Kamei, Y.; Hatazawa, Y.; Uchitomi, R.; Yoshimura, R.; Miura, S. Regulation of skeletal muscle function by amino acids. Nutrients 2020, 12, 261. [Google Scholar] [CrossRef]
- Kusy, K.; Matysiak, J.; Kokot, Z.J.; Ciekot-Sołtysiak, M.; Klupczyńska-Gabryszak, A.; Zarębska, E.A.; Plewa, S.; Dereziński, P.; Zieliński, J. Exercise-induced response of proteinogenic and non-proteinogenic plasma free amino acids is sport-specific: A comparison of sprint and endurance athletes. PLoS ONE 2024, 19, e0309529. [Google Scholar] [CrossRef]
- Zieliński, J.; Kusy, K. Hypoxanthine: A universal metabolic indicator of training status in competitive sports. Exerc. Sport. Sci. Rev. 2015, 43, 214–221. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Ryu, H.-M.; Choi, J.-Y.; Cho, J.-H.; Kim, C.-D.; Park, S.-H.; Kim, Y.-L. Hypoxanthine causes endothelial dysfunction through oxidative stress-induced apoptosis. Biochem. Biophys. Res. Commun. 2016, 482, 821–827. [Google Scholar] [CrossRef]
- Hira, H.S.; Samal, P.; Kaur, A.; Kapoor, S. Plasma level of hypoxanthine/xanthine as markers of oxidative stress with different stages of obstructive sleep apnea syndrome. Ann. Saudi Med. 2014, 34, 308–313. [Google Scholar] [CrossRef]
- Farthing, D.E.; Farthing, C.A.; Xi, L. Inosine and hypoxanthine as novel biomarkers for cardiac ischemia: From bench to point-of-care. Exp. Biol. Med. 2015, 240, 821–831. [Google Scholar] [CrossRef]
- Wu, G.; Fang, Y.-Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef]
- Uchida, Y.; Ferdousi, F.; Takahashi, S.; Isoda, H. Comprehensive transcriptome profiling of antioxidant activities by glutathione in human HepG2 cells. Molecules 2024, 29, 1090. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Matsuba, Y.; Takahashi, M.; Kamano, N.; Watamura, N.; Sasaguri, H.; Takado, Y.; Yoshihara, Y.; Saito, T.; Saido, T.C. Neuronal glutathione loss leads to neurodegeneration involving gasdermin activation. Sci. Rep. 2023, 13, 1109. [Google Scholar] [CrossRef] [PubMed]
- Averill-Bates, D.A. The antioxidant glutathione. Vitam. Horm. 2023, 121, 109–141. [Google Scholar] [CrossRef]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zheng, Y.; Wang, C.; Liu, Y. Glutathione depletion induces ferroptosis, autophagy, and premature cell senescence in retinal pigment epithelial cells. Cell Death Dis. 2018, 9, 753. [Google Scholar] [CrossRef]
- Chen, F.; Kang, R.; Tang, D.; Liu, J. Ferroptosis: Principles and significance in health and disease. J. Hematol. Oncol. 2024, 17, 41. [Google Scholar] [CrossRef]
- Soga, T.; Baran, R.; Suematsu, M.; Ueno, Y.; Ikeda, S.; Sakurakawa, T.; Kakazu, Y.; Ishikawa, T.; Robert, M.; Nishioka, T.; et al. Differential metabolomics reveals ophthalmic acid as an oxidative stress biomarker indicating hepatic glutathione consumption. J. Biol. Chem. 2006, 281, 16768–16776. [Google Scholar] [CrossRef]
- Kaur, G.; Leslie, E.M.; Tillman, H.; Lee, W.M.; Swanlund, D.P.; Karvellas, C.J.; US Acute Liver Failure Study Group. Detection of ophthalmic acid in serum from acetaminophen-induced acute liver failure patients is more frequent in non-survivors. PLoS ONE 2015, 10, e0139299. [Google Scholar] [CrossRef]
- Stewart, G.W. Pyroglutamate acidosis 2023. A review of 100 cases. Clin. Med. 2024, 24, 100030. [Google Scholar] [CrossRef]
- Koga, M.; Serritella, A.V.; Messmer, M.M.; Hayashi-Takagi, A.; Hester, L.D.; Snyder, S.H.; Sawa, A.; Sedlak, T.W. Glutathione is a physiologic reservoir of neuronal glutamate. Biochem. Biophys. Res. Commun. 2011, 409, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Hirai, S.; Horii, S.; Matsuzaki, Y.; Ono, S.; Shimmura, Y.; Sato, K.; Egashira, Y. Anti-inflammatory effect of pyroglutamyl-leucine on lipopolysaccharide-stimulated RAW 264.7 macrophages. Life Sci. 2014, 117, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Torres, A.G.; Ribas de Pouplana, L. Inosine in biology and disease. Genes 2021, 12, 600. [Google Scholar] [CrossRef] [PubMed]
- Basile, M.S.; Bramanti, P.; Mazzon, E. Inosine in neurodegenerative diseases: From the bench to the bedside. Molecules 2022, 27, 4644. [Google Scholar] [CrossRef]
- Módis, K.; Gerő, D.; Stangl, R.; Rosero, O.; Szijártó, A.; Lotz, G.; Mohácsik, P.; Szoleczky, P.; Coletta, C.; Szabó, C. Adenosine and inosine exert cytoprotective effects in an in vitro model of liver ischemia-reperfusion injury. Int. J. Mol. Med. 2013, 31, 437–446. [Google Scholar] [CrossRef]
- Haskó, G.; Kuhel, D.G.; Németh, Z.H.; Mabley, J.G.; Stachlewitz, R.F.; Virág, L.; Lohinai, Z.; Southan, G.J.; Salzman, A.L.; Szabó, C. Inosine inhibits inflammatory cytokine production by a posttranscriptional mechanism and protects against endotoxin-induced shock. J. Immunol. 2000, 164, 1013–1019. [Google Scholar] [CrossRef]
- Mao, B.; Guo, W.; Tang, X.; Zhang, Q.; Yang, B.; Zhao, J.; Cui, S.; Zhang, H. Inosine pretreatment attenuates LPS-induced lung injury through regulating the TLR4/MyD88/NF-κB signaling pathway in vivo. Nutrients 2022, 14, 2830. [Google Scholar] [CrossRef]
- Liang, H.; Song, K. Comprehensive metabolomics and transcriptomics analysis reveals protein and amino acid metabolic characteristics in liver tissue under chronic hypoxia. PLoS ONE 2023, 18, e0291798. [Google Scholar] [CrossRef]
- Hou, Y.; Hu, S.; Li, X.; He, W.; Wu, G. Amino acid metabolism in the liver: Nutritional and physiological significance. Adv. Exp. Med. Biol. 2020, 1265, 21–37. [Google Scholar] [CrossRef]
- Paulusma, C.C.; Lamers, W.H.; Broer, S.; van de Graaf, S.F.J. Amino acid metabolism, transport and signalling in the liver revisited. Biochem. Pharmacol. 2022, 201, 115074. [Google Scholar] [CrossRef]
- Owen, J.B.; Butterfield, D.A. Measurement of oxidized/reduced glutathione ratio. Methods Mol. Biol. 2010, 648, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.A.; Jinnah, H.A.; Kamatani, N. Shortage of cellular ATP as a cause of diseases and strategies to enhance ATP. Front. Pharmacol. 2019, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Smith, Z.H.; Hayden, C.M.T.; Hayes, K.L.; Kent, J.A. Skeletal muscle inosine monophosphate formation preserves ΔGATP during incremental step contractions in vivo. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2025, 328, R195–R205. [Google Scholar] [CrossRef] [PubMed]
- Prokesch, A.; Graef, F.A.; Madl, T.; Kahlhofer, J.; Heidenreich, S.; Schumann, A.; Moyschewitz, E.; Pristoynik, P.; Blaschitz, A.; Knauer, M.; et al. Liver p53 is stabilized upon starvation and required for amino acid catabolism and gluconeogenesis. FASEB J. 2017, 31, 732–742. [Google Scholar] [CrossRef]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Internal Standards Compound | nmol | Internal Standards Compound | nmol |
|---|---|---|---|
| Adenosine-15N5-monophosphate | 5.0 | D5-glutathione | 1.0 |
| Adenosine-15N5-triphosphate | 5.0 | 13C6-isoleucine | 0.5 |
| D4-alanine | 0.5 | D3-lactic acid | 1.0 |
| D7-arginine | 0.5 | D3-leucine | 0.5 |
| D3-aspartic acid | 0.5 | D4-lysine | 0.5 |
| D3-carnitine | 0.5 | D3-methionine | 0.5 |
| D4-citric acid | 0.5 | D6-ornithine | 0.5 |
| 13C1-citrulline | 0.5 | D5-phenylalanine | 0.5 |
| 13C6-fructose-1,6-diphosphate | 1.0 | D7-proline | 0.5 |
| Guanosine-15N5-monophosphate | 5.0 | 13C3-pyruvate | 0.5 |
| Guanosine-15N5-triphosphate | 5.0 | D3-serine | 0.5 |
| 13C6-glucose | 10.0 | D6-succinic acid | 0.5 |
| 13C6-glucose-6-phosphate | 1.0 | D5-tryptophan | 0.5 |
| D3-glutamic acid | 0.5 | D4-tyrosine | 0.5 |
| D5-glutamine | 0.5 | D8-valine | 0.5 |
| TS vs. CS | ||||
| Metabolites | TS (Mean ± SD) | CS (Mean ± SD) | p Value | Log Fold Change |
| Glutathione | 13.195 ± 1.704 | 11.78 ± 2.115 | 0.02065847 | 0.49196 |
| Ophthalmic acid | 0.079 ± 0.038 | 0.035 ± 0.009 | 0.02743137 | 1.3896 |
| Oxiglutathione | 4.451 ± 0.266 | 3.709 ± 0.24 | 0.03274709 | 0.53576 |
| TL vs. CL | ||||
| Metabolites | TL (Mean ± SD) | CL (Mean ± SD) | p Value | Log Fold Change |
| Phenylalanine | 28.285 ± 7.546 | 34.82 ± 9.783 | 0.00198513 | −0.41637 |
| Glycerophosphocholine | 0.716 ± 0.11 | 0.901 ± 0.213 | 0.00386516 | −0.63543 |
| Serine | 88.144 ± 25.328 | 112.205 ± 47.507 | 0.00494991 | −0.70711 |
| Betaine | 0.117 ± 0.021 | 0.123 ± 0.025 | 0.00563138 | −0.6689 |
| Tryptophan | 0.76 ± 0.095 | 0.85 ± 0.159 | 0.00712724 | −0.41976 |
| Leucine | 0.165 ± 0.033 | 0.172 ± 0.036 | 0.00872041 | −0.51078 |
| Methionine | 0.133 ± 0.036 | 0.15 ± 0.041 | 0.0099779 | −0.41337 |
| Citrulline | 0.375 ± 0.048 | 0.439 ± 0.073 | 0.01278198 | −0.51846 |
| Isoleucine | 0.585 ± 0.09 | 0.707 ± 0.141 | 0.01392599 | −0.58514 |
| Valine | 28.285 ± 7.546 | 34.82 ± 9.783 | 0.0183183 | −0.63669 |
| Oxiglutathione | 0.716 ± 0.11 | 0.901 ± 0.213 | 0.02235191 | −0.34569 |
| TS vs. CS | ||||
| Metabolites | TS (Mean ± SD) | CS (Mean ± SD) | p Value | Log Fold Change |
| Creatine | 0.068 ± 0.004 | 0.079 ± 0.005 | 0.00733447 | −0.22613 |
| Xanthine | 0.016 ± 0.005 | 0.208 ± 0.181 | 0.01799637 | −3.7395 |
| Valine | 0.588 ± 0.151 | 1.027 ± 0.243 | 0.021987 | −0.8043 |
| Hypoxanthine | 0.005 ± 0.001 | 0.033 ± 0.02 | 0.02248012 | −2.7161 |
| Lactate | 4.899 ± 1.743 | 10.156 ± 3.972 | 0.03046626 | −1.0516 |
| Pyruvate | 0.42 ± 0.052 | 0.584 ± 0.135 | 0.04361697 | −0.47708 |
| TL vs. CL | ||||
| Metabolites | TL (Mean ± SD) | CL (Mean ± SD) | p Value | Log Fold Change |
| Pyroglutamic acid | 0.641 ± 0.341 | 0.229 ± 0.073 | 0.0101355 | 1.4824 |
| Inosine | 0.032 ± 0.045 | 0.001 ± 0.001 | 0.043386 | 4.6955 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Llorente, L.; Andrés-Gasco, M.; Gil Aranda, M.A.; Rabadán-Ros, R.; Zapata-Pérez, R.; Núñez-Delicado, E.; Menéndez-Coto, N.; García-González, C.; Baena-Huerta, F.J.; Coto-Montes, A.; et al. The Hormetic Adaptative Capacity and Resilience to Oxidative Stress Is Strengthened by Exposome Enrichment with Air Cold Atmospheric Plasma: A Metabolome Targeted Follow-Up Approach. Biomedicines 2025, 13, 949. https://doi.org/10.3390/biomedicines13040949
González-Llorente L, Andrés-Gasco M, Gil Aranda MA, Rabadán-Ros R, Zapata-Pérez R, Núñez-Delicado E, Menéndez-Coto N, García-González C, Baena-Huerta FJ, Coto-Montes A, et al. The Hormetic Adaptative Capacity and Resilience to Oxidative Stress Is Strengthened by Exposome Enrichment with Air Cold Atmospheric Plasma: A Metabolome Targeted Follow-Up Approach. Biomedicines. 2025; 13(4):949. https://doi.org/10.3390/biomedicines13040949
Chicago/Turabian StyleGonzález-Llorente, Lucía, Miguel Andrés-Gasco, Macarena Alba Gil Aranda, Rubén Rabadán-Ros, Rubén Zapata-Pérez, Estrella Núñez-Delicado, Nerea Menéndez-Coto, Claudia García-González, Francisco Javier Baena-Huerta, Ana Coto-Montes, and et al. 2025. "The Hormetic Adaptative Capacity and Resilience to Oxidative Stress Is Strengthened by Exposome Enrichment with Air Cold Atmospheric Plasma: A Metabolome Targeted Follow-Up Approach" Biomedicines 13, no. 4: 949. https://doi.org/10.3390/biomedicines13040949
APA StyleGonzález-Llorente, L., Andrés-Gasco, M., Gil Aranda, M. A., Rabadán-Ros, R., Zapata-Pérez, R., Núñez-Delicado, E., Menéndez-Coto, N., García-González, C., Baena-Huerta, F. J., Coto-Montes, A., & Caso-Peláez, E. (2025). The Hormetic Adaptative Capacity and Resilience to Oxidative Stress Is Strengthened by Exposome Enrichment with Air Cold Atmospheric Plasma: A Metabolome Targeted Follow-Up Approach. Biomedicines, 13(4), 949. https://doi.org/10.3390/biomedicines13040949