
Nonlinear Dynamics of TNFR1 and TNFR2 Expression on Immune Cells: Genetic and Age-Related Aspects of Inflamm-Aging Mechanisms

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Peripheral Blood Mononuclear Cell (PBMC) Isolation and Cultivation
2.2. Assessment of TNF Receptor Expression on PBMC Subpopulations
2.3. Genotyping of TNFR1 and TNFR2 Polymorphisms
2.4. Statistics
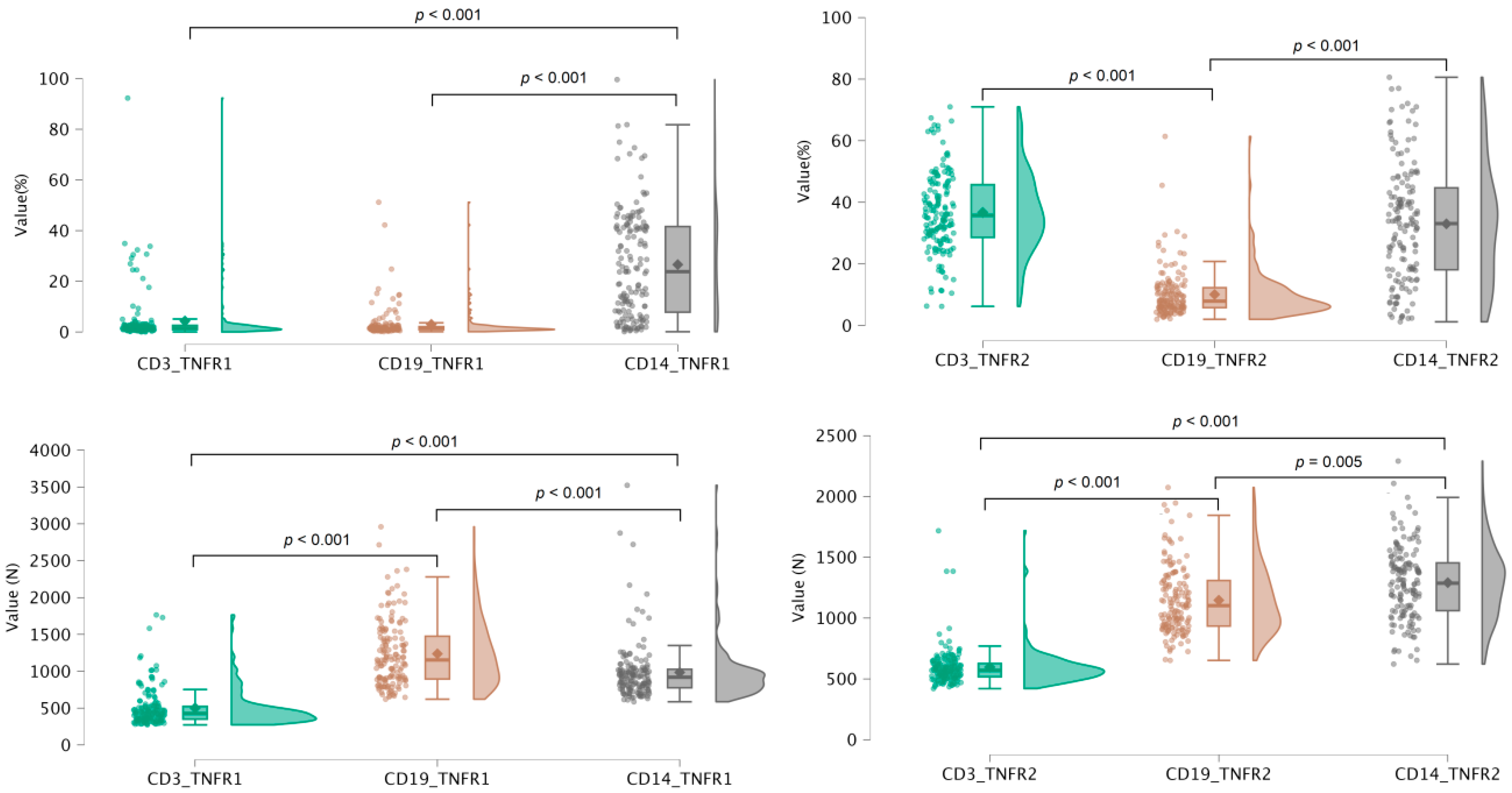
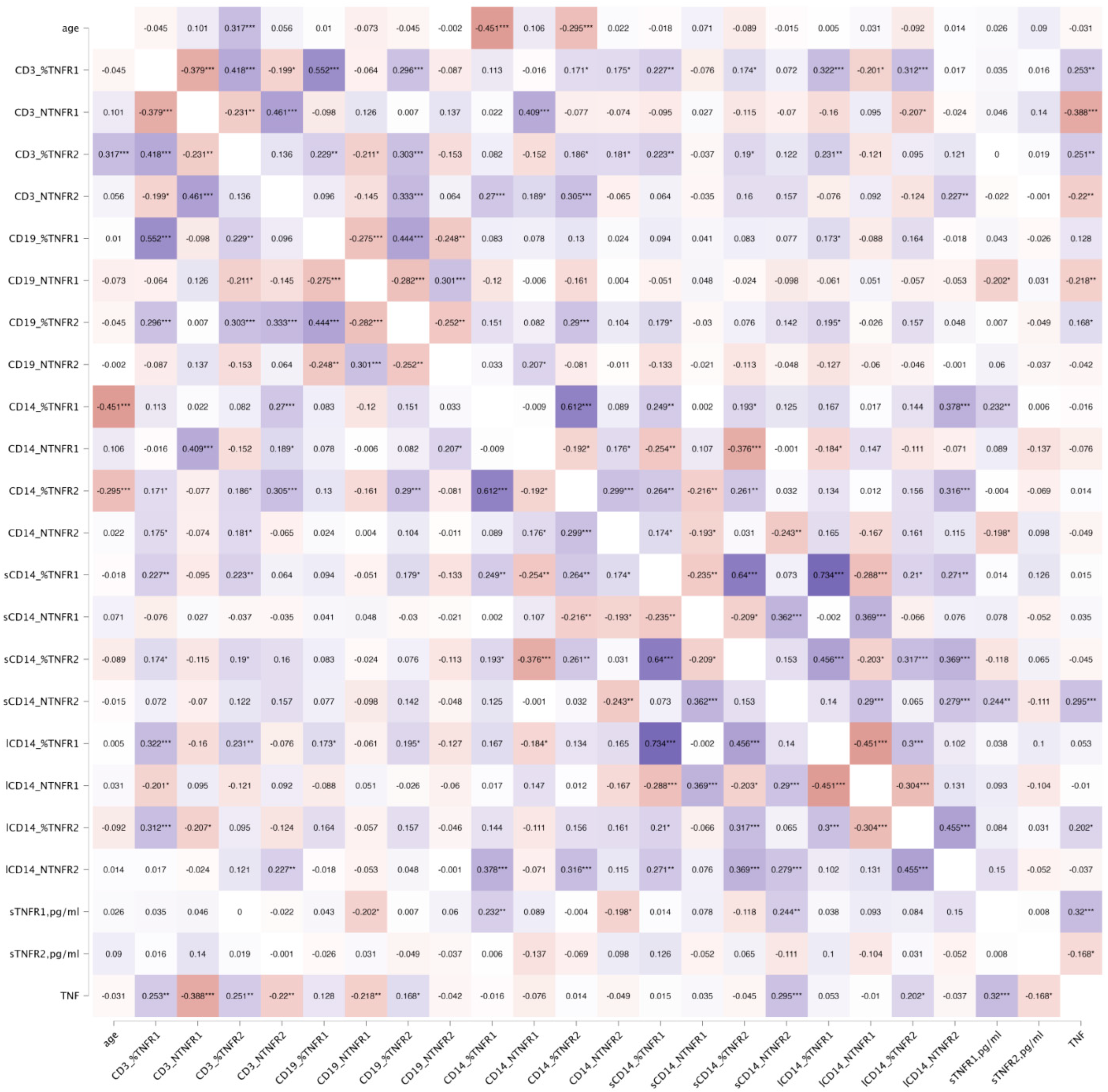
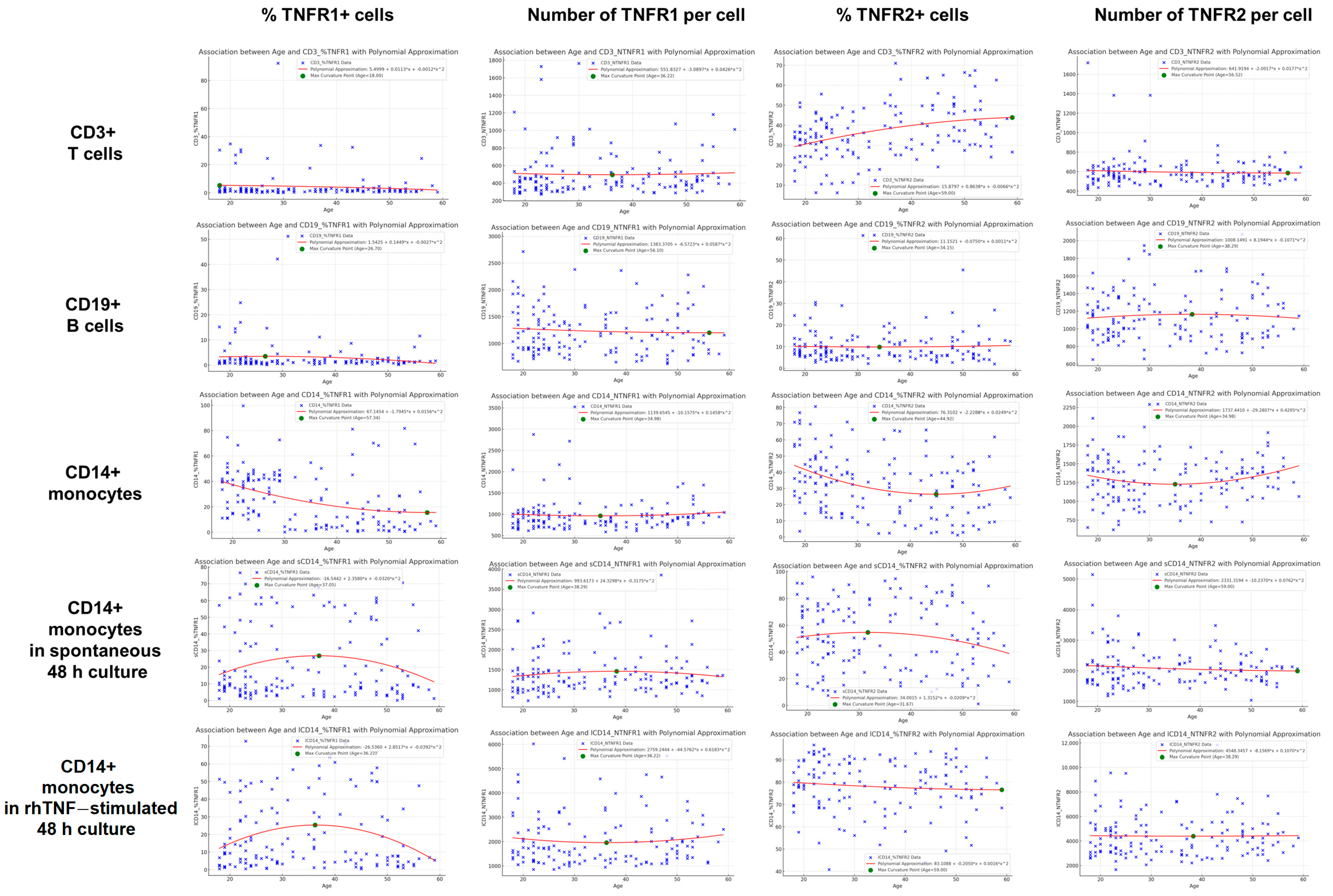
3. Results
3.1. Association Between TNF Receptor Expression and Age Across Genetic Polymorphisms
3.2. Characterization of the Association Types Between TNF Receptor Expression and Age
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, E.D.; Wu, D.; Meydani, S.N. Age-associated alterations in immune function and inflammation. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 118, 110576. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A. Immunosuppressive network promotes immunosenescence associated with aging and chronic inflammatory conditions. J. Mol. Med. 2021, 99, 1553–1569. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging: An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Ajoolabady, A.; Pratico, D.; Tang, D.; Zhou, S.; Franceschi, C.; Ren, J. Immunosenescence and inflammaging: Mechanisms and role in diseases. Ageing Res. Rev. 2024, 101, 102540. [Google Scholar] [CrossRef] [PubMed]
- Varfolomeev, E.; Vucic, D. Intracellular regulation of TNF activity in health and disease. Cytokine 2018, 101, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.H. TNF receptors: Structure-function relationships and therapeutic targeting strategies. Biochim. Biophys. Acta Biomembr. 2025, 1867, 184394. [Google Scholar] [CrossRef] [PubMed]
- Gough, P.; Myles, I.A. Tumor Necrosis Factor Receptors: Pleiotropic Signaling Complexes and Their Differential Effects. Front. Immunol. 2020, 11, 585880. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nikolich-Žugich, J. The twilight of immunity: Emerging concepts in aging of the immune system. Nat. Immunol. 2018, 19, 10–19, Erratum in Nat. Immunol. 2018, 19, 1146. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, J.; Brand, D.D.; Zheng, S.G. Role of TNF-TNF Receptor 2 Signal in Regulatory T Cells and Its Therapeutic Implications. Front. Immunol. 2018, 9, 784. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hearps, A.C.; Martin, G.E.; Angelovich, T.A.; Cheng, W.J.; Maisa, A.; Landay, A.L.; Jaworowski, A.; Crowe, S.M. Aging is associated with chronic innate immune activation and dysregulation of monocyte phenotype and function. Aging Cell 2012, 11, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Saleh, Z.; Mirzazadeh, S.; Mirzaei, F.; Heidarnejad, K.; Meri, S.; Kalantar, K. Alterations in metabolic pathways: A bridge between aging and weaker innate immune response. Front. Aging 2024, 5, 1358330. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Moriwaki, K.; Chan, F.K.M.; Miyoshi, E. Sweet modification and regulation of death receptor signalling pathway. J. Biochem. 2021, 169, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Ross, O.A.; Hinkle, K.M.; Rea, I.M. Cytokine polymorphisms and immunosenescence. In Handbook on Immunosenescence: Basic Understanding and Clinical Applications; Springer: Berlin/Heidelberg, Germany, 2009; pp. 631–658. [Google Scholar] [CrossRef]
- Sennikov, S.V.; Vasilyev, F.F.; Lopatnikova, J.A.; Shkaruba, N.S.; Silkov, A.N. Polymorphisms in the tumor necrosis factor receptor genes affect the expression levels of membrane-bound type I and type II receptors. Mediators Inflamm. 2014, 2014, 745909. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Silkov, A.N.; Sennikova, N.S.; Goreva, E.P.; Lopatnikova, Y.A.; Sennikov, S.V. Production of TNF-α and IL-1β by peripheral blo od mononuclear cells in carriers of different allele variants of the gene. Bull. Exp. Biol. Med. 2012, 153, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.C.; Joshi, S.; Greenwood, H.; Panda, A.; Lord, J.M. Aging of the innate immune system. Curr. Opin. Immunol. 2010, 22, 507–513. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rodríguez-Carrio, J.; Alperi-López, M.; López, P.; Alonso-Castro, S.; Ballina-García, F.J.; Suárez, A. TNFα polymorphism as marker of immunosenescence for rheumatoid arthritis patients. Exp. Gerontol. 2015, 61, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Böyum, A. Isolation of mononuclear cells and granulocytes from human blood. Isolation of monuclear cells by one centrifugation, and of granulocytes by combining centrifugation and sedimentation at 1 g. Scand. J. Clin. Lab. Investig. Suppl. 1968, 97, 77–89. [Google Scholar]
- Allen, R.A.; Lee, E.M.; Roberts, D.H.; Park, B.K.; Pirmohamed, M. Polymorphisms in the TNF-alpha and TNF-receptor genes in patients with coronary artery disease. Eur. J. Clin. Invest. 2001, 31, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Culpan, D.; Cornish, A.; Love, S.; Kehoe, P.G.; Wilcock, G.K. Protein and gene expression of tumour necrosis factor receptors I and II and their promoter gene polymorphisms in Alzheimer’s disease. Exp. Gerontol. 2007, 42, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Willis, E.L.; Eberle, R.; Wolf, R.F.; White, G.L.; McFarlane, D. The effects of age and cytomegalovirus on markers of inflammation and lymphocyte populations in captive baboons. PLoS ONE 2014, 9, e107167. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shin, M.S.; Park, H.J.; Young, J.; Kang, I. Implication of IL-7 receptor alpha chain expression by CD8+ T cells and its signature in defining biomarkers in aging. Immun. Ageing 2022, 19, 66. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Alberti, S.; Cevenini, E.; Ostan, R.; Capri, M.; Salvioli, S.; Bucci, L.; Ginaldi, L.; De Martinis, M.; Franceschi, C.; Monti, D. Age-dependent modifications of Type 1 and Type 2 cytokines within virgin and memory CD4+ T cells in humans. Mech. Ageing Dev. 2006, 127, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Larbi, A.; Douziech, N.; Levesque, I.; Varin, A.; Herbein, G. Cytokine receptor signalling and aging. Mech. Ageing Dev. 2006, 127, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, B.; Wang, H.; Zhao, X.; Zhang, Z.; Ding, G.; Wei, F. Aging affects responsiveness of peripheral blood mononuclear cells to immunosuppression of periodontal ligament stem cells. J. Int. Med. Res. 2020, 48, 300060520930853. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, Q.; Westra, J.; van der Geest, K.S.; Moser, J.; Bijzet, J.; Kuiper, T.; Lorencetti, P.G.; Joosten, L.A.; Netea, M.G.; Heeringa, P.; et al. Reduced levels of cytosolic DNA sensor AIM2 are associated with impaired cytokine responses in healthy elderly. Exp. Gerontol. 2016, 78, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, G.; Braun, P.R.; Hing, B.W.Q.; Ratanatharathorn, A.; Klisares, M.J.; Duncan, G.N.; Jellison, S.S.; Heinzman, J.T.; Nagahama, Y.; Close, L.; et al. Epigenetics of Delirium and Aging: Potential Role of DNA Methylation Change on Cytokine Genes in Glia and Blood Along With Aging. Front. Aging Neurosci. 2018, 10, 311. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kushner, E.J.; Weil, B.R.; MacEneaney, O.J.; Morgan, R.G.; Mestek, M.L.; Van Guilder, G.P.; Diehl, K.J.; Stauffer, B.L.; DeSouza, C.A. Human aging and CD31+ T-cell number, migration, apoptotic susceptibility, and telomere length. J. Appl. Physiol. 2010, 109, 1756–1761. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gupta, S.; Gollapudi, S. CD95-mediated apoptosis in naïve, central and effector memory subsets of CD4+ and CD8+ T cells in aged humans. Exp. Gerontol. 2008, 43, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Quinn, K.M.; Fox, A.; Harland, K.L.; Russ, B.E.; Li, J.; Nguyen, T.H.O.; Loh, L.; Olshanksy, M.; Naeem, H.; Tsyganov, K.; et al. Age-Related Decline in Primary CD8+ T Cell Responses Is Associated with the Development of Senescence in Virtual Memory CD8+ T Cells. Cell Rep. 2018, 23, 3512–3524. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, T.U.; Wilkinson, P.A.; Cameron, M.J.; Ghneim, K.; Chiang, C.; Wertheimer, A.M.; Hiscott, J.B.; Nikolich-Zugich, J.; Haddad, E.K. Human Monocyte Subsets Are Transcriptionally and Functionally Altered in Aging in Response to Pattern Recognition Receptor Agonists. J. Immunol. 2017, 199, 1405–1417. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lu, Y.; Tan, C.T.; Nyunt, M.S.; Mok, E.W.; Camous, X.; Kared, H.; Fulop, T.; Feng, L.; Ng, T.P.; Larbi, A. Inflammatory and immune markers associated with physical frailty syndrome: Findings from Singapore longitudinal aging studies. Oncotarget 2016, 7, 28783–28795. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ronin, E.; Pouchy, C.; Khosravi, M.; Hilaire, M.; Grégoire, S.; Casrouge, A.; Kassem, S.; Sleurs, D.; Martin, G.H.; Chanson, N.; et al. Tissue-restricted control of established central nervous system autoimmunity by TNF receptor 2-expressing Treg cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2014043118. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Islam, M.S.; Yang, Y.; Chen, X. TNF-TNFR2 Signal Plays a Decisive Role in the Activation of CD4+Foxp3+ Regulatory T Cells: Implications in the Treatment of Autoimmune Diseases and Cancer. Adv. Exp. Med. Biol. 2021, 1278, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Ciccacci, C.; Latini, A.; Perricone, C.; Conigliaro, P.; Colafrancesco, S.; Ceccarelli, F.; Priori, R.; Conti, F.; Perricone, R.; Novelli, G.; et al. TNFAIP3 Gene Polymorphisms in Three Common Autoimmune Diseases: Systemic Lupus Erythematosus, Rheumatoid Arthritis, and Primary Sjogren Syndrome-Association with Disease Susceptibility and Clinical Phenotypes in Italian Patients. J. Immunol. Res. 2019, 2019, 6728694. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mendelson, M.M.; Johannes, R.; Liu, C.; Huan, T.; Yao, C.; Miao, X.; Murabito, J.M.; Dupuis, J.; Levy, D.; Benjamin, E.J.; et al. Epigenome-Wide Association Study of Soluble Tumor Necrosis Factor Receptor 2 Levels in the Framingham Heart Study. Front. Pharmacol. 2018, 9, 207. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ha, R.; Keynan, Y.; Rueda, Z.V. Increased susceptibility to pneumonia due to tumour necrosis factor inhibition and prospective immune system rescue via immunotherapy. Front. Cell Infect. Microbiol. 2022, 12, 980868. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Patel, J.R.; Brewer, G.J. Age-related changes to tumor necrosis factor receptors affect neuron survival in the presence of beta-amyloid. J. Neurosci. Res. 2008, 86, 2303–2313. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Richard, A.C.; Peters, J.E.; Lee, J.C.; Vahedi, G.; Schäffer, A.A.; Siegel, R.M.; Lyons, P.A.; Smith, K.G. Targeted genomic analysis reveals widespread autoimmune disease association with regulatory variants in the TNF superfamily cytokine signalling network. Genome Med. 2016, 8, 76. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Munthe-Kaas, M.C.; Torjussen, T.M.; Gervin, K.; Lødrup Carlsen, K.C.; Carlsen, K.H.; Granum, B.; Hjorthaug, H.S.; Undlien, D.; Lyle, R. CD14 polymorphisms and serum CD14 levels through childhood: A role for gene methylation? J. Allergy Clin. Immunol. 2010, 125, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Rechciński, T.; Grebowska, A.; Kurpesa, M.; Peruga, Z.; Dziuba, M.; Krzemińska-Pakuła, M.; Rudnicka, W.; Chmiela, M. CD14 gene polymorphism 159C/T in a group of patients with coronary artery disease from a population with high morbidity of cardiovascular diseases. Kardiol. Pol. 2007, 65, 237–244; discussion 245. [Google Scholar] [PubMed]
- Gowers, I.R.; Walters, K.; Kiss-Toth, E.; Read, R.C.; Duff, G.W.; Wilson, A.G. Age-related loss of CpG methylation in the tumour necrosis factor promoter. Cytokine 2011, 56, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, Y.; Minato, N.; Hattori, M. The impact of senescence-associated T cells on immunosenescence and age-related disorders. Inflamm. Regen. 2018, 38, 24. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Quan, X.Q.; Ruan, L.; Zhou, H.R.; Gao, W.L.; Zhang, Q.; Zhang, C.T. Age-related changes in peripheral T-cell subpopulations in elderly individuals: An observational study. Open Life Sci. 2023, 18, 20220557. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Onyema, O.O.; Njemini, R.; Bautmans, I.; Renmans, W.; De Waele, M.; Mets, T. Cellular aging and senescence characteristics of human T-lymphocytes. Biogerontology 2012, 13, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Sansoni, P.; Vescovini, R.; Fagnoni, F.; Biasini, C.; Zanni, F.; Zanlari, L.; Telera, A.; Lucchini, G.; Passeri, G.; Monti, D.; et al. The immune system in extreme longevity. Exp. Gerontol. 2008, 43, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Kaszubowska, L.; Foerster, J.; Kmieć, Z. NKT-like (CD3+ CD56+) cells differ from T cells in expression level of cellular protective proteins and sensitivity to stimulation in the process of ageing. Immun. Ageing 2022, 19, 18. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nyugen, J.; Agrawal, S.; Gollapudi, S.; Gupta, S. Impaired functions of peripheral blood monocyte subpopulations in aged humans. J. Clin. Immunol. 2010, 30, 806–813. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, C.; Cheng, Y.; Li, B.; Qiu, X.; Hu, H.; Zhang, X.; Lu, Z.; Zheng, F. Transcriptional characteristics and functional validation of three monocyte subsets during aging. Immun. Ageing 2023, 20, 50. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pence, B.D.; Yarbro, J.R. Classical monocytes maintain ex vivo glycolytic metabolism and early but not later inflammatory responses in older adults. Immun. Ageing 2019, 16, 3. [Google Scholar] [CrossRef]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune–metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef]
- De Maeyer, R.P.H.; Chambers, E.S. The impact of ageing on monocytes and macrophages. Immunol. Lett. 2021, 230, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Chen, J.; Liu, K.; Gao, L.; Wu, H.; Ma, L.; Zhou, J.; Liu, Z.; Han, J.J. Human PBMC scRNA-seq-based aging clocks reveal ribosome to inflammation balance as a single-cell aging hallmark and super longevity. Sci. Adv. 2023, 9, eabq7599. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Parker, D.; Sloane, R.; Pieper, C.F.; Hall, K.S.; Kraus, V.B.; Kraus, W.E.; Huebner, J.L.; Ilkayeva, O.R.; Bain, J.R.; Newby, L.K.; et al. Age-Related Adverse Inflammatory and Metabolic Changes Begin Early in Adulthood. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 283–289. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Faubion, L.; White, T.A.; Peterson, B.J.; Geske, J.R.; LeBrasseur, N.K.; Schafer, M.J.; Mielke, M.M.; Miller, V.M. Effect of menopausal hormone therapy on proteins associated with senescence and inflammation. Physiol. Rep. 2020, 8, e14535. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kleeblatt, E.; Lazki-Hagenbach, P.; Nabet, E.; Cohen, R.; Bahri, R.; Rogers, N.; Langton, A.; Bulfone-Paus, S.; Frenkel, D.; Sagi-Eisenberg, R. p16Ink4a-induced senescence in cultured mast cells as a model for ageing reveals significant morphological and functional changes. Immun. Ageing 2024, 21, 77. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gane, J.M.; Stockley, R.A.; Sapey, E. TNF-α Autocrine Feedback Loops in Human Monocytes: The Pro- and Anti-Inflammatory Roles of the TNF-α Receptors Support the Concept of Selective TNFR1 Blockade In Vivo. J. Immunol. Res. 2016, 2016, 1079851. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mytych, J.; Romerowicz-Misielak, M.; Koziorowski, M. Klotho protects human monocytes from LPS-induced immune impairment associated with immunosenescent-like phenotype. Mol. Cell Endocrinol. 2018, 470, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wolk, K.; Grütz, G.; Witte, K.; Volk, H.D.; Sabat, R. The expression of legumain, an asparaginyl endopeptidase that controls antigen processing, is reduced in endotoxin-tolerant monocytes. Genes Immun. 2005, 6, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Hauk, V.; Fraccaroli, L.; Grasso, E.; Eimon, A.; Ramhorst, R.; Hubscher, O.; Pérez Leirós, C. Monocytes from Sjögren’s syndrome patients display increased vasoactive intestinal peptide receptor 2 expression and impaired apoptotic cell phagocytosis. Clin. Exp. Immunol. 2014, 177, 662–670. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alshevskaya, A.; Zhukova, J.; Lopatnikova, J.; Vasilyev, F.; Khutornoy, I.; Golikova, E.; Kireev, F.; Sennikov, S. Nonlinear Dynamics of TNFR1 and TNFR2 Expression on Immune Cells: Genetic and Age-Related Aspects of Inflamm-Aging Mechanisms. Biomedicines 2025, 13, 852. https://doi.org/10.3390/biomedicines13040852
Alshevskaya A, Zhukova J, Lopatnikova J, Vasilyev F, Khutornoy I, Golikova E, Kireev F, Sennikov S. Nonlinear Dynamics of TNFR1 and TNFR2 Expression on Immune Cells: Genetic and Age-Related Aspects of Inflamm-Aging Mechanisms. Biomedicines. 2025; 13(4):852. https://doi.org/10.3390/biomedicines13040852
Chicago/Turabian StyleAlshevskaya, Alina, Julia Zhukova, Julia Lopatnikova, Filipp Vasilyev, Ivan Khutornoy, Elena Golikova, Fedor Kireev, and Sergey Sennikov. 2025. "Nonlinear Dynamics of TNFR1 and TNFR2 Expression on Immune Cells: Genetic and Age-Related Aspects of Inflamm-Aging Mechanisms" Biomedicines 13, no. 4: 852. https://doi.org/10.3390/biomedicines13040852
APA StyleAlshevskaya, A., Zhukova, J., Lopatnikova, J., Vasilyev, F., Khutornoy, I., Golikova, E., Kireev, F., & Sennikov, S. (2025). Nonlinear Dynamics of TNFR1 and TNFR2 Expression on Immune Cells: Genetic and Age-Related Aspects of Inflamm-Aging Mechanisms. Biomedicines, 13(4), 852. https://doi.org/10.3390/biomedicines13040852