
Are There Any Differences in the Healing Capacity between the Medial Collateral Ligament’s (MCL) Proximal and Distal Parts in the Human Knee? Quantitative and Immunohistochemical Analysis of CD34, α-Smooth Muscle Actin (α-SMA), and Vascular Endothelial Growth Factor (VEGF) Expression Regarding the Epiligament (EL) Theory

 , ,
, ,  , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Preparation
2.2. Light Microscopy
2.3. Immunohistochemistry (IHC)
2.4. Semiquantitative Analysis
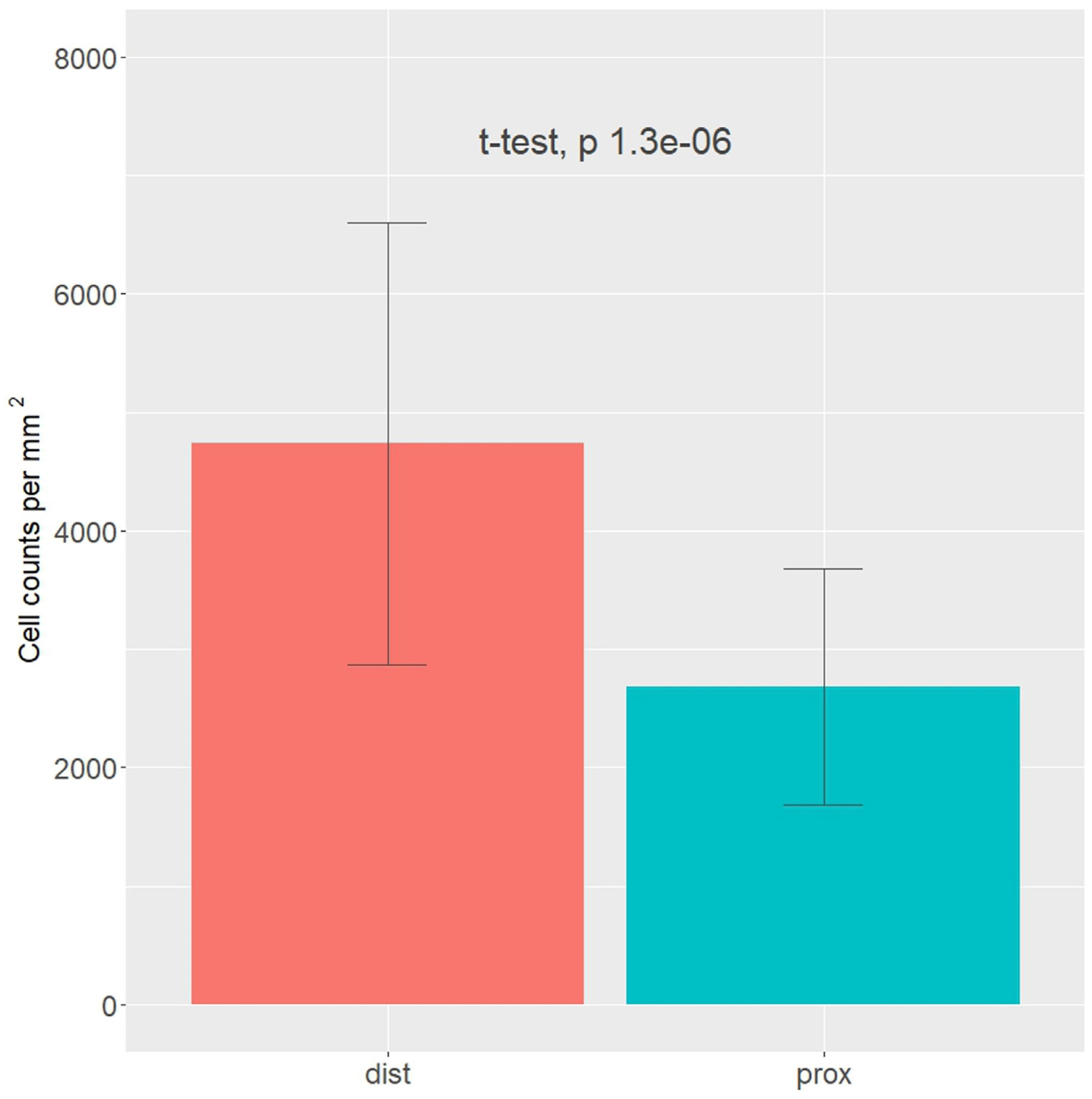
2.5. Cell Numbers
2.6. Statistical Analysis
3. Results
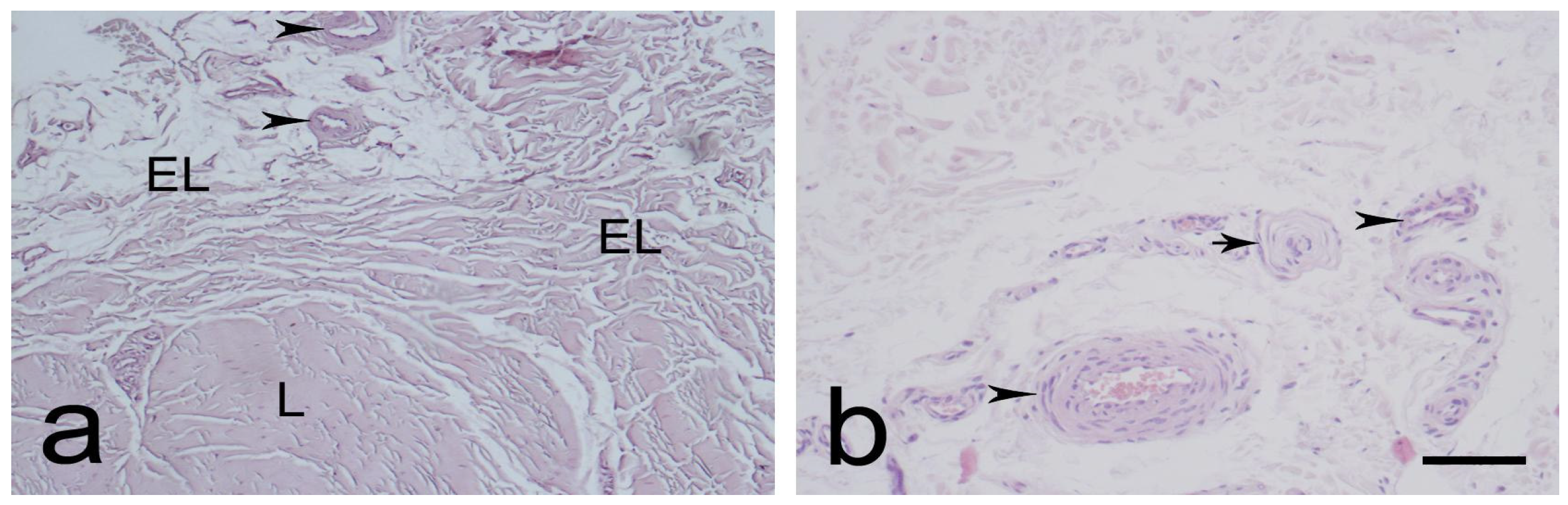
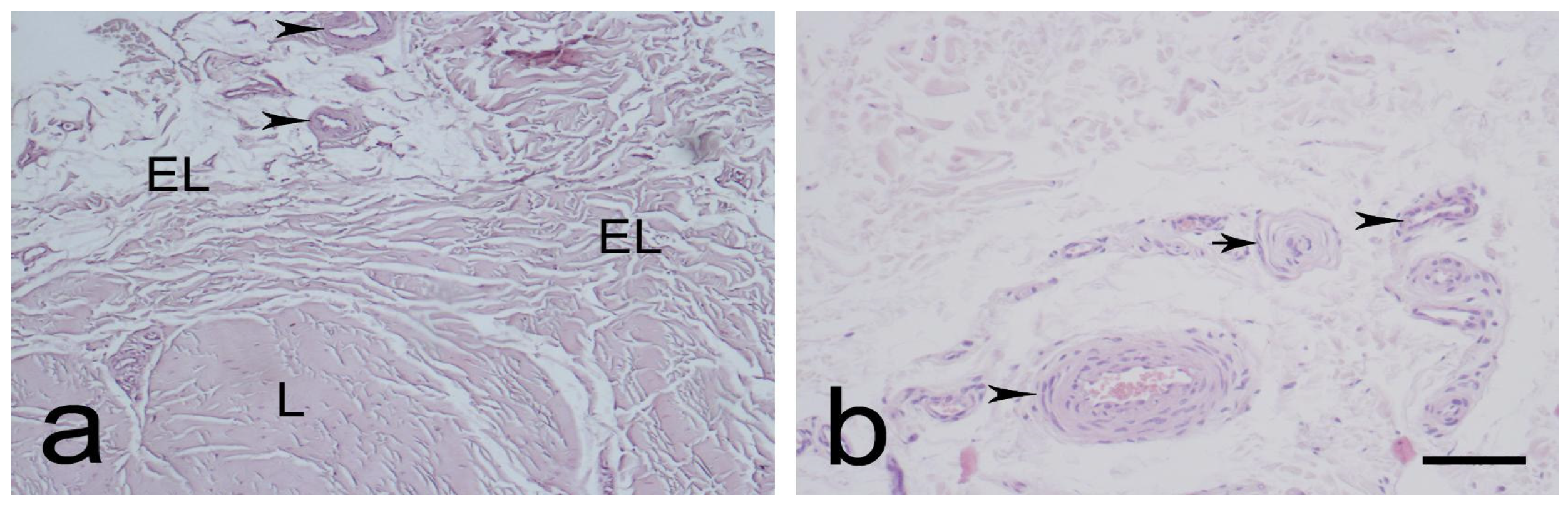
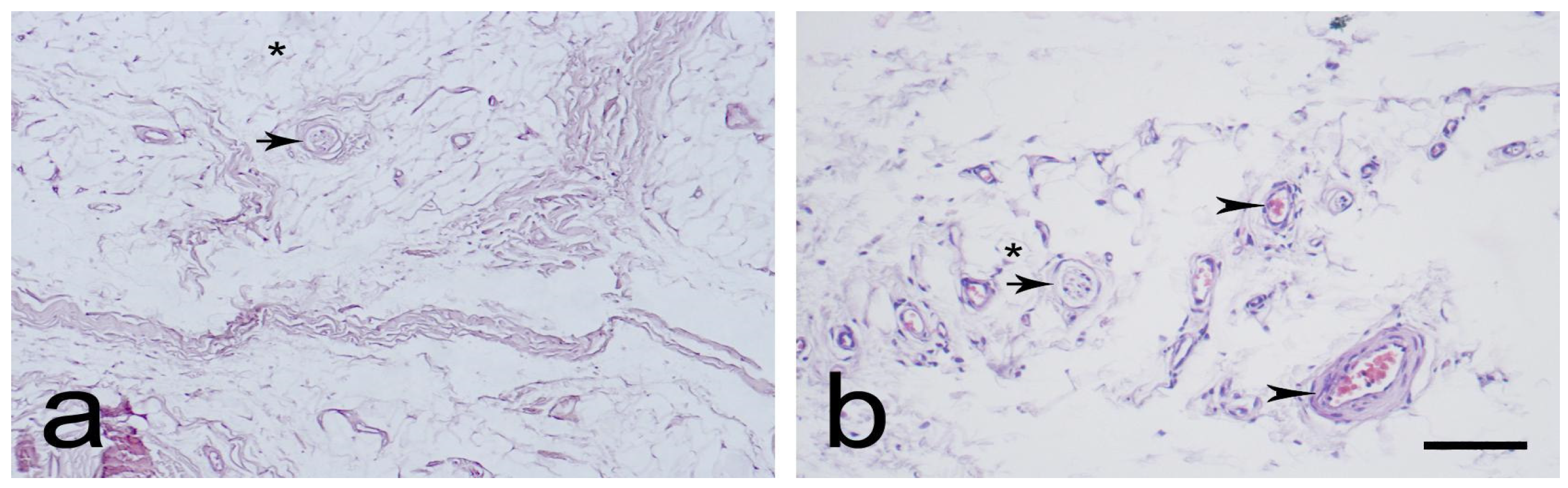
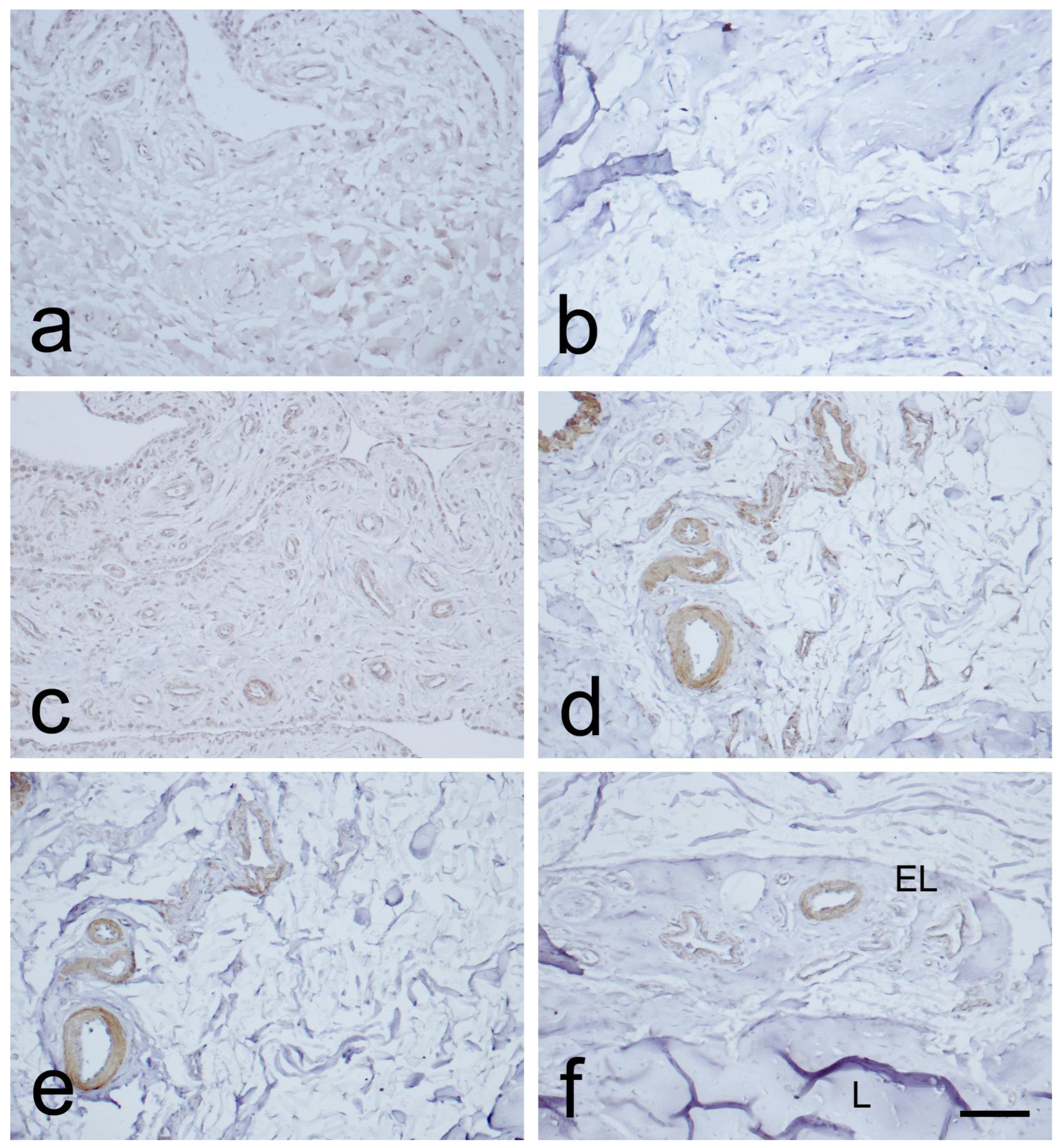
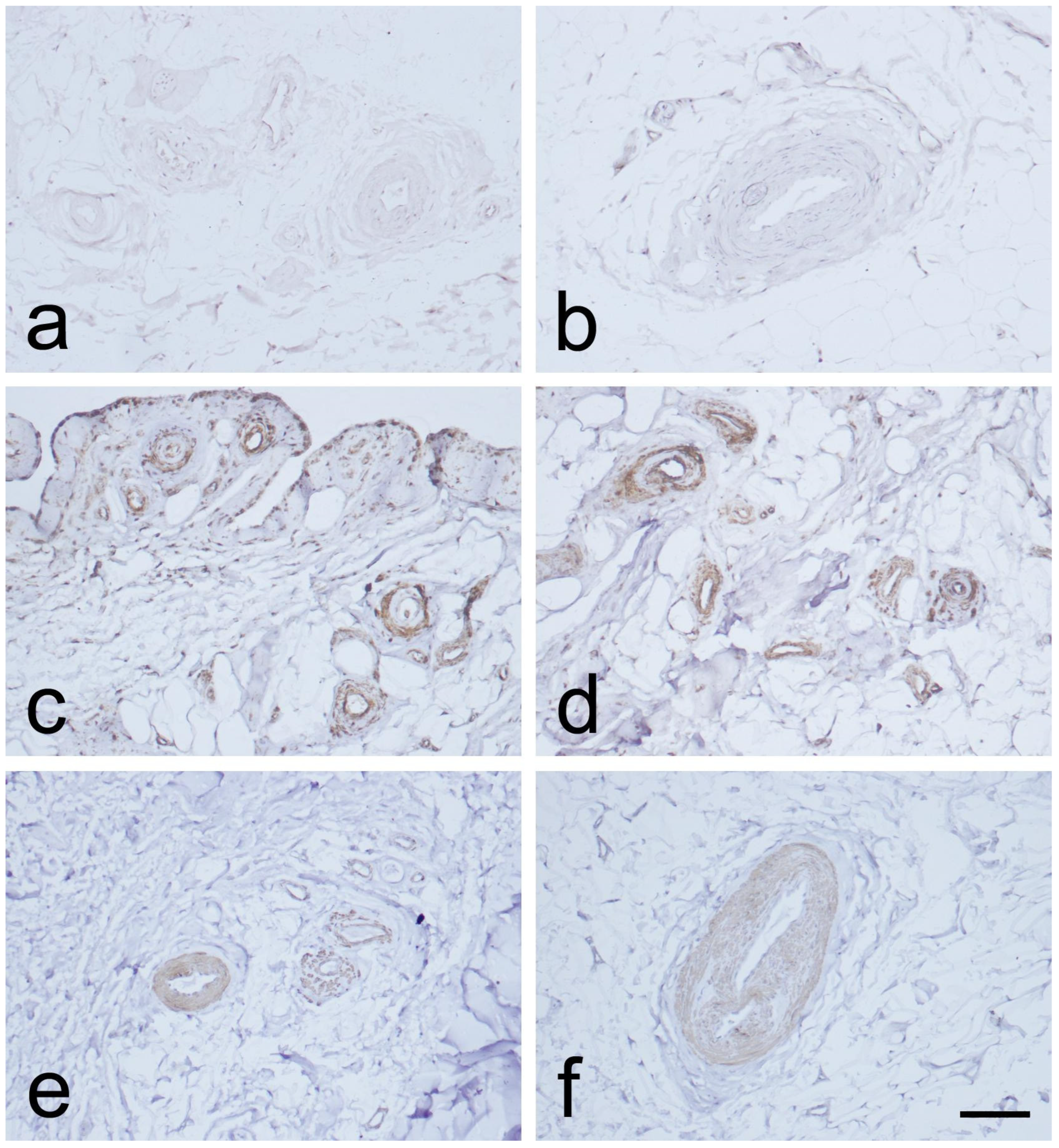
3.1. Light Microscopic Observations
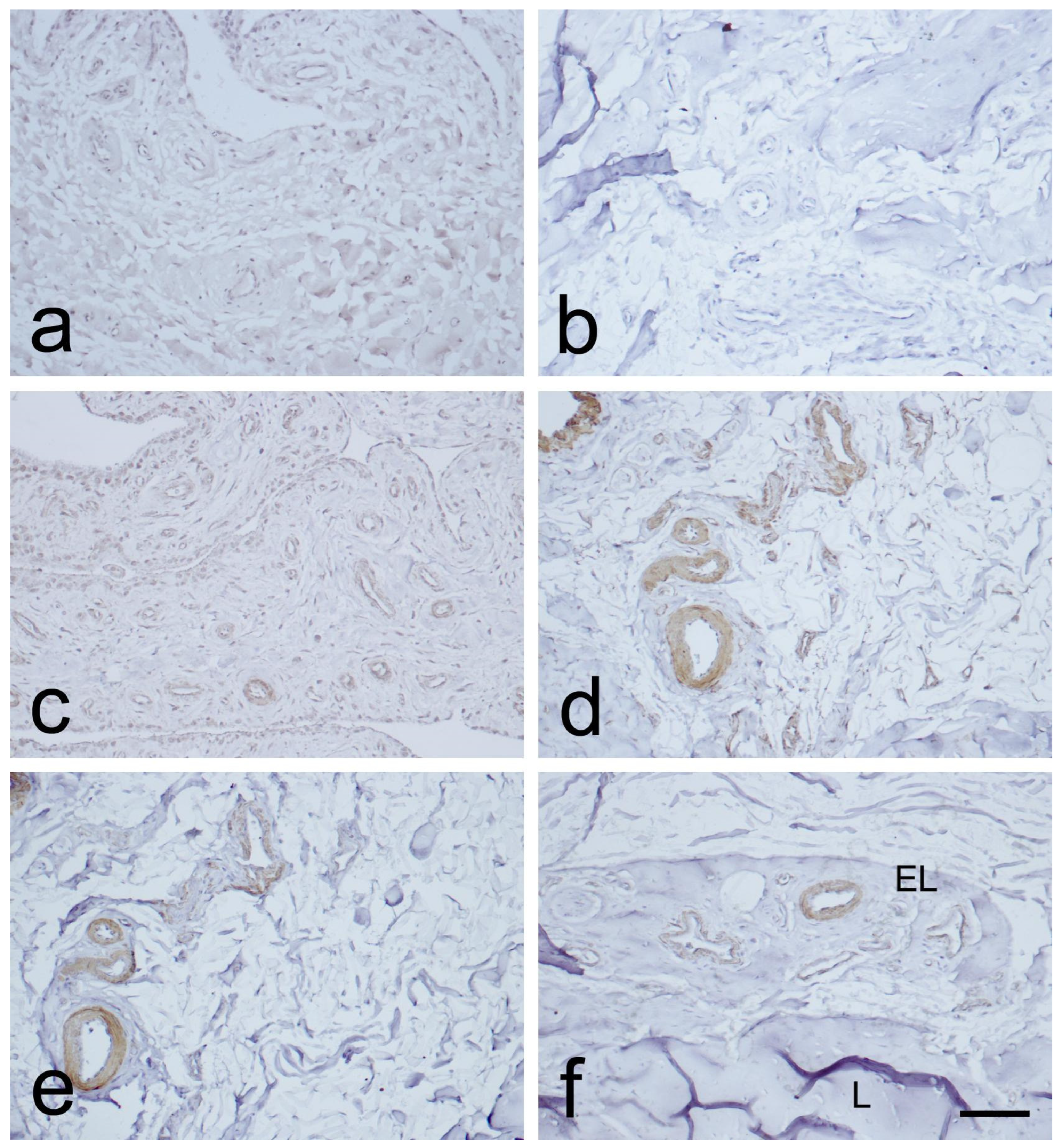
3.2. Expression of CD34, α-SMA, and VEGF in the MCL EL
4. Discussion
- The cadavers’ age could potentially introduce bias due to age-related changes [53]. In response, recently deceased cadavers with a mean age of 55 years, free from osteoarthritis or trauma, were used.
- The subjective nature of visual quantification of IHC images exposes them to considerable inter- and intra-observer variability. The IHC Profiler plugin integrated with ImageJ software was implemented to mitigate this problem.
- Only healthy MCL EL was investigated.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aimes, R.T.; Quigley, J.P. Matrix metalloproteinase-2 is an interstitial collagenase—Inhibitor-free enzyme that catalyzes the cleavage of collagen fibrils and soluble native type-I collagen generating the specific 3/4-length and 1/4-length fragments. J. Biol. Chem. 1995, 270, 5872–5876. [Google Scholar] [CrossRef] [PubMed]
- Blázquez-Prieto, J.; López-Alonso, I.; Amado-Rodríguez, L.; Huidobro, C.; González-López, A.; Kuebler, W.M.; Albaiceta, G.M. Impaired lung repair during neutropenia can be reverted by matrix metalloproteinase-9. Thorax 2018, 73, 321–330. [Google Scholar] [CrossRef]
- Bray, R.C.; Fisher, A.W.; Frank, C.B. Fine vascular anatomy of adult rabbit knee ligaments. J. Anat. 1990, 172, 69–79. [Google Scholar] [PubMed]
- Bray, R.C.; Leonard, C.A.; Salo, P.T. Correlation of healing capacity with vascular response in the anterior cruciate and medial collateral ligaments of the rabbit. J. Orthop. Res. 2003, 21, 1118–1123. [Google Scholar] [CrossRef]
- DeGrace, D.M.; Gill, T.J., IV; Gill, T.J., III. Analysis of medial collateral ligament injuries of the knee. Harvard Orthop. J. 2013, 15, 13–24. [Google Scholar]
- Woo, S.L.; Abramowitch, S.D.; Kilger, R.; Liang, R. Biomechanics of knee ligaments: Injury, healing, and repair. J. Biomech. 2006, 39, 1–20. [Google Scholar] [CrossRef]
- Georgiev, G.P. Epiligament or paratenon is more appropriate for describing the enveloping tissue of the cruciate ligaments of the human knee. Folia Morphol. 2021, 81, 258–259. [Google Scholar] [CrossRef]
- Georgiev, G.P.; Iliev, A.; Kotov, G.; Kinov, P.; Slavchev, S.; Landzhov, B. Light and electron microscopic study of the medial collateral ligament epiligament tissue in human knees. World J. Orthop. 2017, 8, 372–378. [Google Scholar] [CrossRef]
- Georgiev, G.P.; Iliev, A.; Kotov, G.; Nedialkova, V.K.; Kirkov, V.; Landzhov, B. Epiligament Tissue of the medial collateral ligament in rat knee joint: Ultrastructural study. Cureus 2019, 11, e3812. [Google Scholar] [CrossRef]
- Georgiev, G.P.; Kotov, G.; Iliev, A.; Kinov, P.; Angelova, J.; Landzhov, B. Comparison between the operative and non-operative treatment of the medial collateral ligament: Histological and ultrastructural findings during early healing in the epiligament tissue in a rat knee model. Cells Tissues Organs 2018, 206, 165–182. [Google Scholar] [CrossRef]
- Georgiev, G.P.; Landzhov, B.; Dimitrova, I.; Malinova, L.; Ovtscharoff, W. Expression of fibronectin during early healing of the medial collateral ligament epiligament in rat knee model. C. R. Acad. Bulg. Sci. 2016, 69, 639–644. [Google Scholar]
- Georgiev, G.P.; Landzhov, B.; Kotov, G.; Slavchev, S.A.; Iliev, A. Matrix metalloproteinase-2 and -9 expression in the epiligament of the medial collateral and anterior cruciate ligament in human knees: A comparative study. Cureus 2018, 10, e3550. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, G.P.; Vidinov, N.K.; Kinov, P.S. Histological and ultrastructural evaluation of the early healing of the lateral collateral ligament epiligament tissue in a rat knee model. BMC Musculoskelet. Disord. 2010, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, C.S.; Crowley, E.; Vanderby, R. The spatio-temporal dynamics of ligament healing. Wound Repair Regen. 2009, 17, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Iliev, A.A.; Kotov, G.; Stamenov, N.; Landzhov, B.V.; Kirkov, V.; Georgiev, G.P. A comparative immunohistochemical and quantitative study of the epiligament of the medial collateral and anterior cruciate ligament in rat knee. Int. J. Morphol. 2021, 39, 151–159. [Google Scholar] [CrossRef]
- Tavian, M.; Zheng, B.; Oberlin, E.; Crisan, M.; Sun, B.; Huard, J.; Péault, B.M. The vascular wall is a source of stem cells. Ann. N. Y. Acad. Sci. 2005, 1044, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Kirizuki, S.; Matsumoto, T.; Ueha, T.; Uefuji, A.; Inokuchi, T.; Takayama, K.; Hashimoto, S.; Hayashi, S.; Matsushita, T.; Kuroda, R. The influence of ruptured scar pattern on the healing potential of anterior cruciate ligament remnant cells. Am. J. Sports Med. 2018, 46, 1382–1388. [Google Scholar] [CrossRef]
- Howson, K.M.; Aplin, A.C.; Gelati, M.; Alessandri, G.; Parati, E.A.; Nicosia, R.F. The postnatal rat aorta contains pericyte progenitor cells that form spheroidal colonies in suspension culture. Am. J. Physiol. Cell Physiol. 2005, 289, C1396–C1407. [Google Scholar] [CrossRef]
- Chamberlain, C.S.; Crowley, E.M.; Kobayashi, H.; Eliceiri, K.W.; Vanderby, R. Quantification of collagen organization and extracellular matrix factors within the healing ligament. Microsc. Microanal. 2011, 17, 779–787. [Google Scholar] [CrossRef]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol. Biol. Cell. 2001, 12, 2730–2741. [Google Scholar] [CrossRef]
- Hinz, B. Formation and function of the myofibroblast during tissue repair. J. Investig. Dermatol. 2007, 127, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Menetrey, J.; Laumonier, T.; Garavaglia, G.; Hoffmeyer, P.; Fritschy, D.; Gabbiani, G.; Bochaton-Piallat, M.L. α-Smooth muscle actin and TGF-β receptor I expression in the healing rabbit medial collateral and anterior cruciate ligaments. Injury 2011, 42, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Houck, K.; Jakeman, L.; Leung, D.W. Molecular and biological properties of the vascular endothelial growth factor family of proteins. Endocr. Rev. 1992, 13, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Mao, Z.; Hou, Y.; Lin, L.; Xue, T.; Chen, L.; Wang, H.; Yu, C. Local administration of TGFbeta-1/VEGF165 gene-transduced bone mesenchymal stem cells for Achilles allograft replacement of the anterior cruciate ligament in rabbits. Biochem. Biophys. Res. Commun. 2011, 406, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, G.P.; Tubbs, R.S.; Olewnik, Ł.; Zielinska, N.; Telang, M.; Ananiev, J.; Dimitrova, I.N.; Slavchev, S.A.; Yordanov, Y.; LaPrade, R.F.; et al. A comparative study of the epiligament of the medial collateral and anterior cruciate ligaments in the human knee: Immunohistochemical analysis of CD 34, α-smooth muscle actin and vascular endothelial growth factor in relation to epiligament theory. Knee 2022, 39, 78–90. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef] [PubMed]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. ilastik: Interactive machine learning for (bio)image analysis. Nat. Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef]
- Mifune, Y.; Matsumoto, T.; Ota, S.; Nishimori, M.; Usas, A.; Kopf, S.; Kuroda, R.; Kurosaka, M.; Fu, F.H.; Huard, J. Therapeutic potential of anterior cruciate ligament-derived stem cells for anterior cruciate ligament reconstruction. Cell Transplant. 2012, 21, 1651–1665. [Google Scholar] [CrossRef]
- Tei, K.; Matsumoto, T.; Mifune, Y.; Ishida, K.; Sasaki, K.; Shoji, T.; Kubo, S.; Kawamoto, A.; Asahara, T.; Kurosaka, M.; et al. Administrations of peripheral blood CD34-positive cells contribute to medial collateral ligament healing via vasculogenesis. Stem Cells 2008, 26, 819–830. [Google Scholar] [CrossRef]
- Faryniarz, D.A.; Chaponnier, C.; Gabbiani, G.; Yannas, I.V.; Spector, M. Myofibroblasts in the healing lapine medial collateral ligament: Possible mechanisms of contraction. J. Orthop. Res. 1996, 14, 228–237. [Google Scholar] [CrossRef]
- Ferrara, N.; Davis-Smyth, T. The biology of vascular endothelial growth factor. Endocr. Rev. 1997, 18, 4–25. [Google Scholar] [CrossRef] [PubMed]
- Molloy, T.; Wang, Y.; Murrell, G. The roles of growth factors in tendon and ligament healing. Sports Med. 2003, 33, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Nicosia, R.F.; Lin, Y.J.; Hazelton, D.; Qian, X. Endogenous regulation of angiogenesis in the rat aorta model. Role of vascular endothelial growth factor. Am. J. Pathol. 1997, 151, 1379–1386. [Google Scholar] [PubMed]
- Jackson, J.R.; Minton, J.A.; Ho, M.L.; Wei, N.; Winkler, J.D. Expression of vascular endothelial growth factor in synovial fibroblasts is induced by hypoxia and interleukin 1beta. J. Rheumatol. 1997, 24, 1253–1259. [Google Scholar] [PubMed]
- Zumstein, M.A.; Rumian, A.; Lesbats, V.; Schaer, M.; Boileau, P. Increased vascularization during early healing after biologic augmentation in repair of chronic rotator cuff tears using autologous leukocyte- and platelet-rich fibrin (L-PRF): A prospective randomized controlled pilot trial. J. Shoulder Elbow Surg. 2014, 23, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Flamme, I.; Reutern, M.V.; Drexler, H.C.; Syed-Ali, S.; Risau, W. Overexpression of vascular endothelial growth factor in the avian embryo induces hypervascularization and increased vascular permeability without alterations of embryonic pattern formation. Dev. Biol. 1995, 171, 399–414. [Google Scholar] [CrossRef]
- Corral, C.J.; Siddiqui, A.; Wu, L.; Farrell, C.L.; Lyons, D.; Mustoe, T.A. Vascular endothelial growth factor is more important than basic fibroblastic growth factor during ischemic wound healing. Arch. Surg. 1999, 134, 200–205. [Google Scholar] [CrossRef]
- Nishimori, M.; Matsumoto, T.; Ota, S.; Kopf, S.; Mifune, Y.; Harner, C.D.; Ochi, M.; Fu, F.H.; Huard, J. Role of angiogenesis after muscle derived stem cell transplantation in injured medial collateral ligament. J. Orthop. Res. 2012, 30, 627–633. [Google Scholar] [CrossRef]
- Phillips, G.D.; Stone, A.M.; Jones, B.D.; Schultz, J.C.; Whitehead, R.A.; Knighton, D.R. Vascular endothelial growth factor (rhVEGF165) stimulates direct angiogenesis in the rabbit cornea. In Vivo 1994, 8, 961–965. [Google Scholar]
- Kanazawa, T.; Soejima, T.; Murakami, H.; Inoue, T.; Katouda, M.; Nagata, K. An immunohistological study of the integration at the bone-tendon interface after reconstruction of the anterior cruciate ligament in rabbits. J. Bone Jt. Surg. Br. 2006, 88, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Frank, C.; Amiel, D.; Akeson, W.H. Healing of the medial collateral ligament of the knee. A morphological and biochemical assessment in rabbits. Acta Orthop. Scand. 1983, 54, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Breuls, R.G.; Klumpers, D.D.; Everts, V.; Smit, T.H. Collagen type V modulates fibroblast behavior dependent on substrate stiffness. Biochem. Biophys. Res. Commun. 2009, 380, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Frank, C.; Shrive, N.; Hiraoka, H.; Nakamura, N.; Kaneda, Y.; Hart, D. Optimization of the biology of soft tissue repair. J. Sci. Med. Sport 1999, 2, 190–210. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, G.; Iliev, A.; Landzhov, B.; Dimitrova, I.; Kotov, G.; Malinova, L.; Ovtscharoff, W. Localization of matrix metalloproteinase-2 in injured medial collateral ligament epiligament in rat knee. C. R. Acad. Bulg. Sci. 2017, 70, 273–278. [Google Scholar]
- Lo, I.K.; Marchuk, L.L.; Leatherbarrow, K.E.; Frank, C.B.; Hart, D.A. Collagen fibrillogenesis and mRNA levels in the maturing rabbit medial collateral ligament and patellar tendon. Connect. Tissue Res. 2004, 45, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, G.P.; Landzhov, B.; Dimitrova, I.N.; Slavchev, S.; Malinova, L.; Kartelov, Y.; Ankova, D.; Ovtscharoff, W. Light microscopic and immunohistochemical study of the medial collateral ligament epiligament in rat knee. C. R. Acad. Bulg. Sci. 2015, 68, 95–100. [Google Scholar]
- Chowdhury, P.; Matyas, J.R.; Frank, C.B. The “epiligament” of the rabbit medial collateral ligament: A quantitative morphological study. Connect. Tissue Res. 1991, 27, 33–50. [Google Scholar] [CrossRef]
- Berkson, E.M.; Nolan, D.; Fleming, K.; Spang, R.P.; Wong, J.S.; Asnis, P.D.; Kawadler, J. Knee: Ligamentous and patellar tendon injuries. Pathol. Interv. Musculoskelet. Rehabil. 2016, 2, 740. [Google Scholar]
- Lee, J.K.; Jo, S.; Lee, Y.L.; Park, H.; Song, J.S.; Sung, I.H.; Kim, T.H. Anterior cruciate ligament remnant cells have different potentials for cell differentiation based on their location. Sci. Rep. 2020, 10, 3097. [Google Scholar] [CrossRef]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef]
- Zengin, E.; Chalajour, F.; Gehling, U.M.; Ito, W.D.; Treede, H.; Lauke, H.; Weil, J.; Reichenspurner, H.; Kilic, N.; Ergün, S. Vascular wall resident progenitor cells: A source for postnatal vasculogenesis. Development 2006, 133, 1543–1551. [Google Scholar] [CrossRef]
- Iwanaga, J.; Singh, V.; Ohtsuka, A.; Hwang, Y.; Kim, H.J.; Moryś, J.; Ravi, K.S.; Ribatti, D.; Trainor, P.A.; Sañudo, J.R.; et al. Acknowledging the use of human cadaveric tissues in research papers: Recommendations from anatomical journal editors. Clin. Anat. 2021, 34, 2–4. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
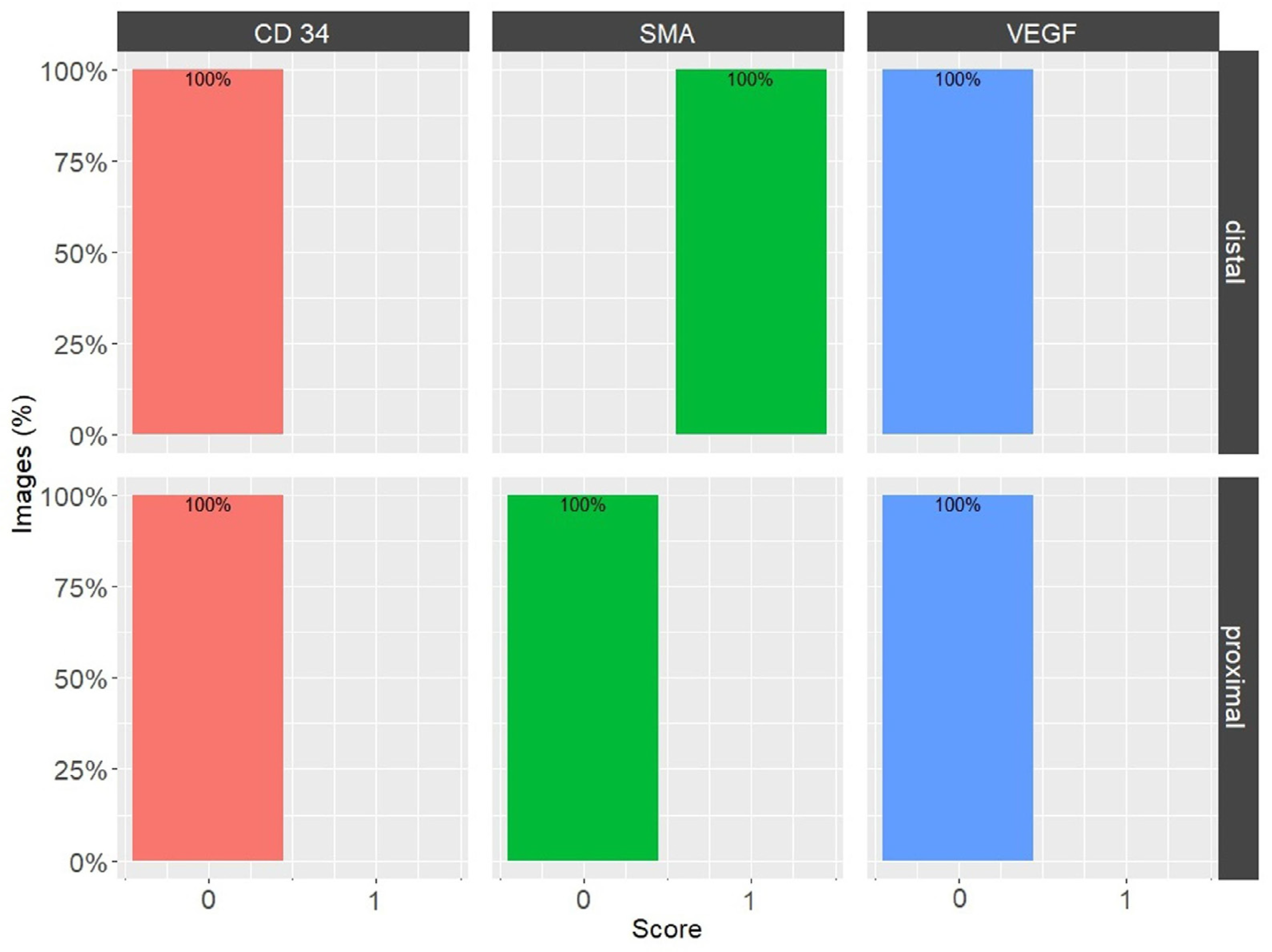
| IHC Marker | Distal Part of the MCL EL | Proximal Part of the MCL EL | ||
|---|---|---|---|---|
| Score | % | Score | % | |
| VEGF | High Positive (3+) | 0.1 | High Positive (3+) | 0.1 |
| Positive (2+) | 2.3 | Positive (2+) | 2.1 | |
| Low Positive (1+) | 14.5 | Low Positive (1+) | 14.6 | |
| Negative (0) | 83.1 | Negative (0) | 83.2 | |
| CD34 | High Positive (3+) (0.0%) | 0.0 | High Positive (3+) | 0.1 |
| Positive (2+) | 0.4 | Positive (2+) | 0.7 | |
| Low Positive (1+) | 3.8 | Low Positive (1+) | 6.1 | |
| Negative (0) | 95.9 | Negative (0) | 93.0 | |
| α-SMA | High Positive (3+) | 2.5 | High Positive (3+) | 0.2 |
| Positive (2+) | 12.8 | Positive (2+) | 5.1 | |
| Low Positive (1+) | 23.0 | Low Positive (1+) | 21.0 | |
| Negative (0) | 61.8 | Negative (0) | 73.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgiev, G.P.; Yordanov, Y.; Gaydarski, L.; Tubbs, R.S.; Olewnik, Ł.; Zielinska, N.; Piagkou, M.; Ananiev, J.; Dimitrova, I.N.; Slavchev, S.A.; et al. Are There Any Differences in the Healing Capacity between the Medial Collateral Ligament’s (MCL) Proximal and Distal Parts in the Human Knee? Quantitative and Immunohistochemical Analysis of CD34, α-Smooth Muscle Actin (α-SMA), and Vascular Endothelial Growth Factor (VEGF) Expression Regarding the Epiligament (EL) Theory. Biomedicines 2024, 12, 659. https://doi.org/10.3390/biomedicines12030659
Georgiev GP, Yordanov Y, Gaydarski L, Tubbs RS, Olewnik Ł, Zielinska N, Piagkou M, Ananiev J, Dimitrova IN, Slavchev SA, et al. Are There Any Differences in the Healing Capacity between the Medial Collateral Ligament’s (MCL) Proximal and Distal Parts in the Human Knee? Quantitative and Immunohistochemical Analysis of CD34, α-Smooth Muscle Actin (α-SMA), and Vascular Endothelial Growth Factor (VEGF) Expression Regarding the Epiligament (EL) Theory. Biomedicines. 2024; 12(3):659. https://doi.org/10.3390/biomedicines12030659
Chicago/Turabian StyleGeorgiev, Georgi P., Yordan Yordanov, Lyubomir Gaydarski, Richard Shane Tubbs, Łukasz Olewnik, Nicol Zielinska, Maria Piagkou, Julian Ananiev, Iva N. Dimitrova, Svetoslav A. Slavchev, and et al. 2024. "Are There Any Differences in the Healing Capacity between the Medial Collateral Ligament’s (MCL) Proximal and Distal Parts in the Human Knee? Quantitative and Immunohistochemical Analysis of CD34, α-Smooth Muscle Actin (α-SMA), and Vascular Endothelial Growth Factor (VEGF) Expression Regarding the Epiligament (EL) Theory" Biomedicines 12, no. 3: 659. https://doi.org/10.3390/biomedicines12030659
APA StyleGeorgiev, G. P., Yordanov, Y., Gaydarski, L., Tubbs, R. S., Olewnik, Ł., Zielinska, N., Piagkou, M., Ananiev, J., Dimitrova, I. N., Slavchev, S. A., Terziev, I., Suwannakhan, A., & Landzhov, B. (2024). Are There Any Differences in the Healing Capacity between the Medial Collateral Ligament’s (MCL) Proximal and Distal Parts in the Human Knee? Quantitative and Immunohistochemical Analysis of CD34, α-Smooth Muscle Actin (α-SMA), and Vascular Endothelial Growth Factor (VEGF) Expression Regarding the Epiligament (EL) Theory. Biomedicines, 12(3), 659. https://doi.org/10.3390/biomedicines12030659