Abstract
Rift Valley fever is a vector-borne zoonotic disease caused by the Rift Valley fever virus (Phlebovirus genus) listed among the eight pathogens included in the Bluepoint list by the WHO. The transmission is mainly vehicled by Aedes and Culex mosquito species. Symptoms of the disease are varied and non-specific, making clinical diagnosis often challenging, especially in the early stages. Due to the difficulty in distinguishing Rift Valley fever from other viral hemorrhagic fevers, as well as many other diseases that cause fever, an early diagnosis of the infection is important to limit its spread and to provide appropriate care to patients. To date, there is no validated point-of-care diagnostic tool. The virus can only be detected in the blood for a brief period, suggesting that molecular methods alone are not sufficient for case determination. For this, it is preferable to combine both molecular and serological tests. The wide distribution of competent vectors in non-endemic areas, together with global climate change, elicit the spread of RVFV to continents other than Africa, making surveillance activities vital to prevent or to limit the impact of human outbreaks and for a rapid identification of positive cases, making diagnosis a key factor for this achievement.
1. Introduction
Rift Valley fever virus (RVFV) is included in the Bluepoint list by the World Health Organization (WHO) together with other pathogens (Ebola virus, Zika virus, Lassa fever virus, Nipah virus, Crimean–Congo hemorrhagic fever virus, severe acute respiratory syndrome coronavirus, and Middle East respiratory syndrome coronavirus), which are prioritized for research and development [1].
RVFV is a vector-borne zoonotic disease caused by a phlebovirus (family Phenuiviridae), firstly described in 1931 during an epidemic outbreak in the Rift Valley region of Kenya, with high rates of abortion among pregnant ewes and acute deaths of newborn lambs [2].
The RVFV is a phlebovirus belonging to the order Bunyavirales and family Phenuiviridae. This virus has a single-stranded RNA genome consisting of three segments: a small (S), medium (M), and large (L) segment, all of negative or ambisense polarity. The S segment encodes for the nucleoprotein (N), while its anti-genomic RNA is responsible for encoding the non-structural NSs protein, which acts as a major virulence factor [3]. The M segment encodes glycoproteins precursor Gc and Gn, together with the nonstructural proteins NSm and the proteins P78, P14, and P13. The L segment encodes the viral RNA-dependent RNA polymerase (RdRp), as shown in Figure 1 [3].
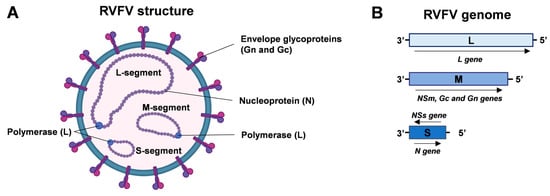
Figure 1.
RVFV structure and genome organization. (A) Enveloped RVFV virion comprises nucleoprotein (N) and envelope glycoproteins (Gn and Gc). Viral polymerase (L) and N proteins are associated with the viral genomic segments (RNA, negative or ambisense polarity). (B) Schematic representation of RVFV genome organization. The L segment encodes the viral RNA polymerase; the M segment encodes NSm, Gc, and Gn proteins; and the S segment encodes the N protein and NSs protein (ambisense polarity). Created with Biorender.com.
Viral reassortment can occur among Phlebovirus genera due to their segmented genomic RNA. Furthermore, genomic reassortment among different RVFV strains is also known to occur and has been demonstrated experimentally [4].
Ngari virus, which is grouped in the genus Orthobunyavirus within the Bunyaviridae family, is a zoonotic arbovirus, and it was identified during a RVFV outbreak in Mauritania in 2010; this can lead to co-infections in goats [5].
Another study with the RVFV MP-12 strain and a genetic variant showed that out of 47 plaques isolated from coinfected C6/36 mosquito cells, 83% were reassortants [6].
Protection against RVFV in all animal species is conferred by neutralizing antibodies, which can be detected within the first week post-infection [7]. Nucleoproteins (N) induce high levels of IgG and IgM antibodies in RVFV and other bunyavirus infections, but there is no evidence that anti-N antibodies exhibit virus-neutralizing activity [8].
In a study, long-lived IgG and T cell responses were detected against viral envelope glycoproteins, Gn and Gc. However, antigen-specific antibody depletion experiments showed that Gn-specific antibodies dominate the RVFV neutralizing antibodies (nAb) response. Finally, IgG avidity against Gn, but not Gc, correlated with nAb titers [9].
Recently, Quellec et al. (2023) showed that RVFV-infected astrocytes upregulated expression of genes associated with inflammatory and type I interferon responses at the mRNA level but not at the protein level [10].
The life cycle of RVFV is intricately linked with its transmission dynamics and the interactions between the virus, mosquitoes, and vertebrate hosts. Transmission predominantly occurs through the bites of infected mosquitoes, with Aedes and Culex species being key vectors. Upon entering a host organism, RVFV targets specific cells, initiating its replication process, which mainly occurs in the cell cytoplasm and probably starts near the site of endosomal fusion [11]. The S, M, and L segments are transcribed and translated, leading to the production of viral proteins and the formation of new viral particles within the Golgi apparatus. The assembled virions are then released from the host cell by exocytosis, allowing the virus to spread within the host organism and causing infections in various tissues. Importantly, infected animals serve as reservoirs for the virus, facilitating its transmission to mosquito vectors during blood feeding. As mosquitoes become infected, they play a crucial role in perpetuating the cycle by transmitting the virus to other animals or humans during subsequent bites. The transmission cycles of RVFV are summarized in Figure 2.
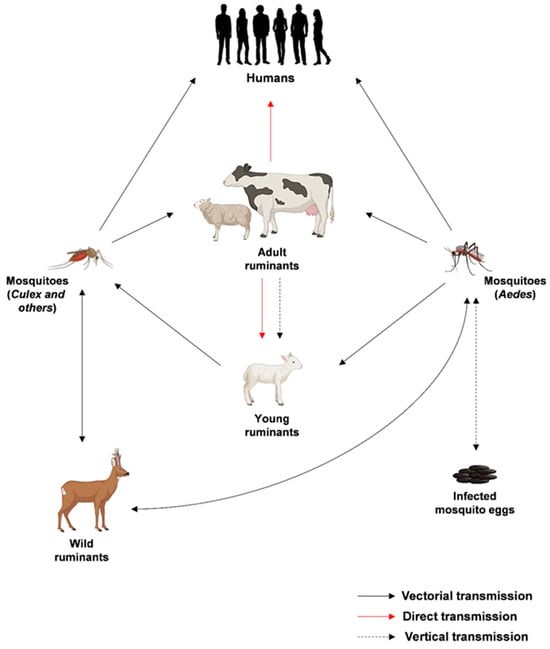
Figure 2.
Epidemiological transmission cycles of RVFV. Created with Biorender.com.
RVFV has an incubation period of 2–6 days in humans.
In most cases, people infected with RVFV are asymptomatic or paucisymptomatic (fever, headache, weakness, back pain, vertigo, anorexia, photophobia, and dizziness). Approximately 10% of people with RVFV develop severe symptoms, with eye disease in 0.5–2% of patients. Encephalitis or inflammation in the brain is observed in 1% of cases. Finally, only 1% of cases may manifest hemorrhagic fever. Hemorrhagic fever can lead to up to 50% fatality, and after about a week after the onset of hemorrhagic symptoms, there is death [8,12].
A histopathological examination of infected human brains demonstrated encephalitis with an infiltration of lymphocytes and necrosis in neurons [13]. Drosophila melanogaster (D. melanogaster) and vertebrates have similarities for the neural proliferation and brain circuit formation. Therefore, D. melanogaster can be used to better understand what cellular and molecular mechanisms of Rift Valley viruses are involved in neurological disorders [14].
Wild animals tend to have mild or inapparent infection, whereas domestic animals are more susceptible to RVF disease [15]. Clinical signs range from sudden death or abortion to mild, non-specific symptoms, depending on the virulence of the virus strain and the species involved. Mortality may reach 70–100% in lambs and kids, and 20–70% in adult sheep and calves. Cattle and camels are less susceptible to infection, and abortion rates may reach 85–100% within the affected herds [16]. In pregnant sheep or goats, RVFV infection results in above 100% fetal mortality [17].
RVFV infection in older non-pregnant animals is often asymptomatic, and abortion may be the only overt manifestation of the disease in a herd or flock [18].
Transmission between animals by direct contact with infected tissues or fluids has been observed, together with iatrogenic route by use of infected needles used for vaccination, particularly in endemic regions with limited economic resources [7,19]. The infections in humans can occur by inhaling aerosols of infectious body fluids, and consumption of raw or unpasteurized milk has also been identified as a risk factor for RVFV infection [12].
The RVFV is transmitted from different mosquito species to animals or humans and from infected animals to humans [8,20].
RVFV-competent mosquito vectors fall under 73 species of mosquitoes in the 8 genera of the family Culicidae. Trans-ovarian or vertical transmission is also reported [20,21].
It is transmitted from ruminants to ruminants by mosquito bites, mainly from the genera Aedes and Culex, but also from the genera Anopheles and Mansonia, as recently suggested in Madagascar and Kenya [22,23]. Animals are infectious for mosquitoes during the viremic period. Viremia may be brief (6–18 h) or persist for 6 to 8 days [24].
However, the severity of clinical signs is different on the species: sheep are more susceptible than goats, which are themselves more susceptible than cattle and camels [25].
Human-to-human transmission of the RVFV has not been demonstrated, but vertical transmission readily occurs in animals and has also been reported in humans [26,27].
Outbreaks from West African Sahelian areas are not closely related to high rainfall, which is a contributing factor. In years with short-term dry periods punctuated by occasional precipitation, double cycles of Aedes vectors are observed, together with simultaneous waves of Aedes and Culex vectors [28,29].
Most human cases do not require treatment against RVFV. For severe cases of Rift Valley fever, there is no specific treatment but only a general supportive care treatment [12].
Several outbreaks have been described as causing severe economic and health consequences [1].
Overall, from 2000 to 2016, 11 outbreaks of RVFV occurred in humans in the Republic of Niger (2016), the Republic of Mauritania (2012), the Republic of South Africa (2010), Madagascar (2008 and 2009), Sudan (2007), Kenya, Somalia, Tanzania (2006), Egypt (2003), Saudi Arabia, and Yemen (2000) [30]. The fatality rate was about 19.5%, with 950 reported death cases [30]. To date, no outbreaks have been reported in Europe [31].
The percentage of seroprevalence in humans ranges from 1.8% (Kenya) to 11.1% (Saudi Arabia) [32,33]. In countries along the Mediterranean basin, anti-RVFV IgG prevalence ranged from 1.4% in Tunisia to 4.9% in Turkey [34,35], while in Italy, the authors found the antibody anti-IgG Rift Valley fever virus in 2.6% of serum patients with suspect arbovirus infection [36].
Wild animals have been found to be seropositive for RVFV antibodies; during the epizootic period in 2006–2007, the 31.8% of wild ruminants surveyed in Kenya were positive for anti-RVFV antibodies [37].
Considering that mosquitoes of the Aedes and Culex species are now circulating in Italy and Europe, mostly due to climate change, there is a risk of arbovirus introduction to continents other than Africa [38].
2. Epidemiology
The first reported case of RVFV was in 1930 in Kenya, during an outbreak in sheep [30]. Afterwards, outbreaks affecting livestock and humans occurred in other countries of Africa such as Egypt, Kenya, South Africa, Madagascar, Mauritania, Senegal, and Gambia [38]. Over the past 25 years, RVF disease has expanded its historic geographic range in the livestock-raising areas of eastern and southern Africa and into the Middle East (Saudi Arabia and Yemen), following an infected livestock trade from the horn of Africa, causing several epizootics and epidemics [39]. This first episode of RFV in humans and animals outside the African continent raised concerns that it could be extended in other parts of Asia. According to what was reported for 2019 from the CDC, as for 2017 and 2018, European countries reported four cases of RVF in humans. There were imported cases from Western Africa: three from Mali (first confirmed case in 2015 and second European confirmed cases in 2016) [30,40] and one from Ghana (one suspected case in 2016) [41].
All cases were males, and the mean age at infection was 32 years. All four cases were European residents. No associated deaths were reported [42]. RVFV was historically confined to the African continent until 2000, and it is enzootic in many African countries and Madagascar, see Figure 3. Global changes, including climatic ones, may cause an expansion in the geographical distribution of the RVFV, together with legal or illegal animal movements and mosquitoe distribution [43,44].
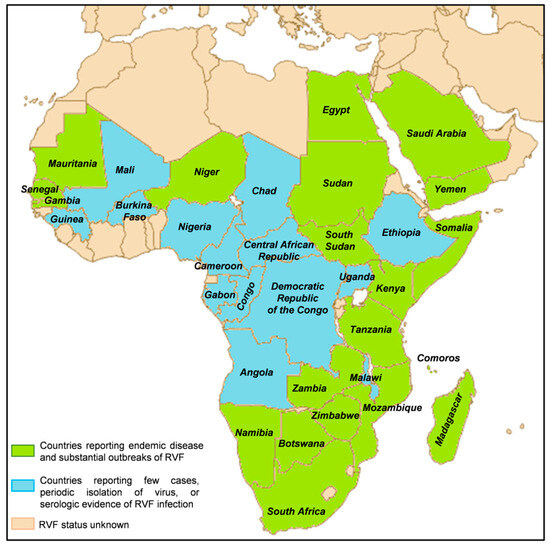
Figure 3.
RVF distribution map. Created with Biorender.com.
3. Diagnosis
The early stages of Rift Valley infection are difficult to diagnose because the symptoms are nonspecific. In particular, the symptomatology is similar to that of other hemorrhagic fevers and fever-causing infections such as malaria, shigellosis, typhoid fever, and yellow fever.
3.1. Molecular Diagnostics
The RNA detection of the RVFV genome has been performed over the years with several methods.
At the beginning of the 2000s, a one-step RT-PCR method was developed with a sensitivity of 0.5 pfu/reaction. This method was able to detect several RVFV strains such as Gabek, Forest, Gordil, Saint Floris, Arumwot, Belterra, ArD38661, AnD100286, and MP12 [45]. Afterwards, different real-time PCR methods were developed, targeting the G2 gene, a region of the S segment [46,47,48,49]. RT-loop-mediated amplification (RT-LAMP) is currently the most used platform. This method uses different regions of the viral genome as a template: L segment [50,51], S segment [52,53], or M segment [54]. The most sensitive assay was based on the L region target: 10 copies/reaction [51]. Moreover, an isothermal recombinase polymerase amplification (RPA) assay had been set up by Euler, reaching a low limit detection (LLOD) of 19 RNA molecules/reaction [53]. Furthermore, RVFV detection was included in a multiplex RT-qPCR for hemorrhagic fever pathogens. However, the sensitivity was very low in comparison with those reached by RT-LAMP or real-time RT-PCR (105–106 copies/mL) [55]. The identification of different RVFV strains could be carried out by analysis of the melting curve of the L, M, and S regions amplification [56]. Other assays that were based on the microarray method were developed; one detecting the GP gene was validated with a real-time PCR, but few samples were tested, and no data about specificity were reported [57]. A second one detected the M segment with an average detection limit of 6.36 DNA copies, targeting the other 28 target pathogens on the array in addition to the RVFV, see Table 1 [58].
Although NGS (next-generation sequencing) has been applied for the detection of numerous arboviruses, including Chikungunya, Zika [59], and West Nile viruses [60], and show a sensitivity comparable to real-time PCR, it has not yet been developed for RVFV.

Table 1.
Molecular assays for RVFV.
Table 1.
Molecular assays for RVFV.
| Test | Test Type | Virus Detected | Target Gene | Biological Matrix | Reference Assay | Sample Size | Sensitivity | Specificity | Author |
|---|---|---|---|---|---|---|---|---|---|
| RVFV RT-nested PCR | Homemade one-step RT-PCR nested method | RVFV different strains (Gabek forest, Gordil, saint Floris, Arumwot, Belterra, ArD38661, AnD100286, MP 12) | NS coding region of S segment | Virus produced in Vero E6 cells; serum from infected mice | Virus isolation method | ND | 0.5 pfu/reaction | Nd | Sall et al., 2001 [45] |
| RVFV quantitative real-time PCR | qRT-PCR with fluorescent signal from probes for quality control | RVFV; MP12, ZH501, ZH548, ArD38661, 74 HB59 strains | NS coding region of S segment | Virus produced in Vero E6 cells; serum from infected mice | ND | ND | 50–100 copies/reaction | No amplification with Toscana, Icoraci, and Belterra closely related phlebovirus | Garcia et al., 2001 [46] |
| RT-Real-time PCR | 5′ nuclease technology on a light cycler instrument | RVFV | G2 gene | Synthetic RNA | ND | ND | 2835 geq/mL | no cross-reactivity with other HCV, HBV, HSV1, CMV, Modoc virus, Mycobacterium tuberculosis, Mycobacterium leprae, Borrelia spp., Leptospira spp., Neisseria spp., Plasmodium spp., Leishmania spp. | Drosten et al., 2002 [48] |
| RT-Real-time PCR homemade | Fluorescent nested PCR TaqMan assay | RVFV | S segment | Synthetic RNA | ND | ND | 100 copies/reaction | SFNV cell culture | Weidmann et al., 2008 [47] |
| Real-time qRT-PCR homemade | qRT-PCR with fluorescent reporter dye detected at each PCR cycle | RVFV | G2 gene | Plasma of suspected patients with HVF | 272 RVFV confirmed cases | 2ND | 100 infectious particles/mL | IgM anti RVFV positive sera 100% | Njenga et al., 2009 [49] |
| RT-LAMP homemade | Reverse transcription-loop-mediated isothermal amplification with a vertical | RVFV | L segment | Serum samples | TaqMan Real Time | 64 | Whole blood: LLOD: 10 copies RNA/reaction | No cross reactivity with phleboviruses; flaviviruses and chikungunja virus | Peyrefitte et al., 2008 [50] |
| RT-LAMP homemade | Reverse transcription-loop-mediated isothermal amplification with a vertical | RVFV | L segment | Bleed samples from sheep (n = 20), human plasma from suspected cases (n = 65); 3 liver, kidney, serum from animals | Whole blood: LLOD: 10 copies/ reaction | Six African phleboviruses and unrelated arbovirus did not give cross reactivity | Le Roux et al., 2009 [51] | ||
| RT-LAMP homemade | Reverse transcription-loop-mediated isothermal amplification | RVFV | S segment | Synthetic RNA | Real-time RT-PCR | ND | whole blood: LLOD: 1.94 copies/microliters within 60 min | No cross reactivity with JEV, H3N2 influenza virus, EBOV, MARV | Han et al., 2020 [52] |
| RT-LAMP homemade | Reverse transcription-loop-mediated isothermal amplification | RVFV | M segment | Blood samples | Real-time RT-PCR | 130 | 98.36% sensitivity | 100%; no cross-reactivity with PPR and capripox viruses | Wekesa et al., 2023 [54] |
| Isothermal recombinase polymerase amplification (RPA) | Isothermal exponential nucleic acid amplification and detection method | RVFV | S segment | Synthetic RNA | ND | ND | 19RNA molecules/reaction | No cross reactivity with Yersinia pestis, Francisella tularensis, Bacillus antracis, vaccinia virus, Ebola virus, Marburg virus, Crimean–Congo virus and phleboviruses | Euler et al., 2012 [53] |
| RT-qPCR genotyping assay | One step RT-qPCR for typing different strains of RVFV, melting curve to identify different strains of RVFV | RVFV | L, M, S segments | ND | Sanger sequencing | ND | Balaraman et al., 2023 [56] | ||
| BioT DNA multiplex PCR-enzyme hybridization assay | Multiplex RT-PCR | RVFV | GP2 gene | 196 swabs, 45 skin swabs,15 serum, 7 sputum | ND | 260 clinical samples | 105–106 copies/mL with nucleic acid extraction | No cross reactivity with Influenza A, EBV, CMV, RSV A, ADV C, human metapneumovirus | He et al., 2009 [55] |
| Oligonucleotide microarray | Microarray | RVFV | GP gene | Culture samples | Real-time PCR | 60 | 100% | Yao et al., 2021 [57] | |
| Real-time qRT-PCR commercial | qRT-PCR with fluorescent reporter dye detected at each PCR cycle | RVFV | 0.89 copies/μL | cross-reactivity with flavivirus, Marburg virus, and Ebola virus | [61] |
ADV, adenovirus; CMV, cytomegalovirus; RSV, respiratory syncytial virus; JEV, Japanese encephalitis virus.
3.2. Serological Diagnosis
Serological assays are key to epidemiological studies for the identification of active infection or previous exposure to the RVFV [62]. Serological diagnosis of RVF can be performed in the laboratory using the following tests:
- Virus neutralization test (VNT)
- Indirect immunofluorescent assay (IFA)
- IgG and IgM antibody enzyme-linked immunosorbent assay (ELISA)
ELISA tests and virus neutralization tests are the most widely used methods for antibody detection. The agar gel immunodiffusion (AGID), radioimmunoassays, hemagglutination inhibition (HI), and complement fixation (CF) are disused. Animal samples used for antibody detection may contain live viruses, and therefore, it would be important to apply inactivation procedures before proceeding to the sample testing. The inactivation step could be conducted by a combination of thermal and chemical inactivation [63].
Multiplex detection systems for the detection of antibodies against multiple highly pathogenic agents simultaneously are being developed, making them valuable tools for disease surveillance and diagnosis [64,65]. In the case of RVFV, a specific in situ hybridization (ISH) has been reported for the detection of viral RNA of several RVFV strains in different fixed tissues [66].
3.2.1. Virus Neutralization Test Assay
To detect specific neutralizing antibodies against RVFV, neutralization tests to the virus are used, such as the plaque reduction neutralization test (PRNT).
VNT is generally accepted as one of the standard assay systems for the quantitative determination of neutralizing antibodies in serum samples in naturally infected and vaccinated animals, but while it is highly specific and useful to test samples from any species, it is also costly, time consuming, and requires a high biosecurity laboratory capable of working safely with live RVFV. VNT is very specific, with cross reactions with other phleboviruses being limited [67,68]. This assay is performed in a BSL3 laboratory and requires highly specialized and vaccinated personnel. VNT is useful to check the protective immunity of a serum that is correlated to the level of neutralizing antibodies, see Figure 4. The sera are serially diluted and incubated with a well-defined number of viral particles (100–300 median tissue culture infective dose (TCID50) per mL) before being added to Vero or baby hamster kidney (BHK) cells (3–4 × 105 cells per mL). After 3/5 days, the cytopathic effect of RVFV infection can be observed using a microscope. For confirmation of the results, plates can be fixed with 10% formalin containing 0.05% crystal violet and re-visualized [69]. Gn and Gc are targets for neutralizing antibodies and play an important role in host cell entry and exit [70].
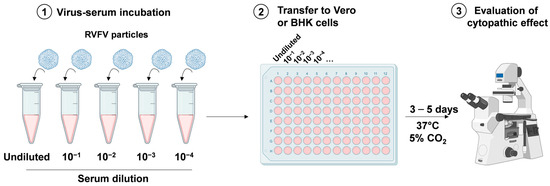
Figure 4.
Viral neutralization assay. Serum to be tested is serially diluted and incubated with a defined quantity of viral particles (1). The virus-serum solutions are added to Vero or BHK cells (2) and incubated for 3/5 days at 37 °C in a humidified atmosphere with 5% CO2. Finally, the cytopathic effect of viral infection is evaluated through microscopic observation (3). Created with Biorender.com.
3.2.2. Viral Isolation
RVFV can be identified during the acute phase of the disease by viral isolation from whole blood or serum. In post-mortem cases, the virus can also be isolated from other body districts such as the brain, liver, spleen, and organs of aborted fetuses [71].
In natural and experimental infections, RVFV infects many tissues.
The RVFV antigen can be present in various tissues of naturally infected sheep, such as the liver, kidney, lungs, testes, vascular tissues, and adrenal glands [72]. In the past, an in vitro culture of live agents was considered the principal method for viral detection, but it has been replaced by PCR and next-generation sequencing. This assay is not used for primary diagnosis because it is less sensitive than PCR. Rift Valley viral isolation, Figure 5, can be performed in different cell lines, including African green monkey kidney (Vero), baby hamster kidney (BHK), and AP61 mosquito cells [73]; a period ranging from 3 to 6 days is necessary to detect the cytopathic effect, and the procedure requires a level 3 laboratory (BSL3 facilities).
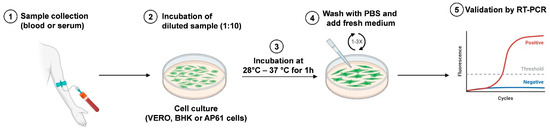
Figure 5.
Schematic representation of RVFV isolation. The blood or serum sample is collected from infected hosts (1) and inoculated in cell culture (2). After incubation at 28 °C (AP61 cells) or 37 °C (Vero and BHK cells) for 1 h (3), cell monolayers are washed with PBS and fresh medium is added (4). Virus isolation is validated by RT-PCR analysis (5). Created with Biorender.com.
They are inoculated with 1/10 dilution of the sample and incubated at 37 °C for 1 h (in the case of mosquito cell lines, the incubation should be performed at 28 °C for 1 h).
After removal of the inoculum, washing with phosphate-buffered saline (PBS) or culture media is performed. Finally, the wash solution is removed, and culture medium is added and incubated for 5–6 days.
The confirmation of virus isolation should be performed using a reverse-transcription polymerase chain reaction (RT-PCR) [74].
3.2.3. Indirect Immunofluorescent Assay
Immunofluorescence assays are still used, although cross-reactions may occur between RVFV and other phleboviruses [69].
An indirect immunofluorescent assay (IFA) using virus-infected Vero cells can be used for the detection of antibodies of Rift Valley fever viruses in BSL3. The samples can be tested at a screening dilution 1:20, both for IgM and IgG detection. Moreover, the positive samples were serially diluted from 1:20 down to 1:1280 to estimate the antibody titer. The IgG and IgM titers are reported as the reciprocal of the highest dilution with positive fluorescence [36]. To date, there are Euroimmun commercial tests for IFA Rift Valley fever virus IgG and IgM detection [75].
3.2.4. ELISA Assay
The ELISA test is a sensitive and rapid test suited to the needs of large-scale testing and the most widely used method for the detection of IgM and IgG isotypes, also commercially available.
IgG and IgM are indeed key markers for RVFV seroprevalence studies, but data on kinetics or persistence during human convalescence are limited. ELISA tests are used routinely in many countries for single-case diagnosis, outbreak management, and surveillance. IgM-capture ELISA allows for the diagnosis of recent infections. IgG-ELISA could be used to determine the rise in antibody response.
The ELISA test is a very versatile test that can be performed with an inactivated antigen, but cross-reactions may occur between the RVF virus and other phleboviruses.
An ELISA (enzyme-linked immunosorbent assay) kit is commercially available to detect anti-Rift Valley fever virus IgG and IgM antibodies, and it is used for serological diagnosis in ruminant animals. At the earliest, it can detect antibodies as soon as four days following infection or vaccination in animals reacting very early, and eight days post-vaccination for 100% of animals. The diagnostic sensitivity was in cattle 84.31%, buffalo 94.44%, sheep 98.91%, and goat 99.18%. The diagnostic specificity was in cattle 99.34%, buffalo 98.28%, sheep 99.16%, goat 99.23%, and other game ruminants 99.26% [76]. Another indirect ELISA based on a recombinant RVFV nucleoprotein has been developed using the cloning, sequencing, and bacterial expression of the N protein of the RVFV. Sera from 106 laboratory workers vaccinated with inactivated RVFV, 16 patients infected with RVFV, 168 serial bleeds from 8 sheep infected by experimentation with RVFV, and 210 serial bleeds from 10 sheep vaccinated with a live attenuated Smithburn RVFV strain were used in this study. The sera positive in the indirect test (I-ELISA) were also positive in the virus-neutralization test. A high correlation (R2 = 0.8571) was found between the tests in human vaccines [77].
Another study also used the recombinant nucleoprotein (N) of the RVF virus to test for diagnostic applicability in an indirect ELISA (I-ELISA). The experimentally infected sheep were 128, vaccinated sheep were 240, and field-collected sera in sheep were 251, goats were 362, and cattle were 100. In goats, the diagnostic sensitivity and specificity of the I-ELISA was 100% when using the anti-species IgG conjugate. Using protein G as a detection system, the sensitivity and specificity in goats were 99.4% and 99.5%, respectively; in sheep, the field sera were both 100%, while in cattle, they were 100% and 98.3%, respectively. The I-ELISA based on the recombinant N-protein has the potential to complement the traditional assays for the serodiagnosis of RVFV [78].
An inhibition (competitive) ELISA has been validated, and the sensitivity and specificity were valuated. In fact, it is now commercially available for the detection of antibodies in different species and in domestic and wild ruminants. Field samples were collected from mainland France for the known-negative sera in 192 cattle, 119 goats, 192 sheep, and from ruminants of a French overseas territory (Mayotte) for the known-positive sera. This study showed a sensitivity and specificity of 100%. The results demonstrate that this ELISA may be a suitable diagnostic tool for disease surveillance programs [79].
Commercial assay kits are available, and several in-house protocols have been published [80,81,82]; furthermore, any ELISA has been validated for the detection of antibodies in different species, including humans and domestic and wild ruminants [79,83]. Sensitivity and specificity vary according to the different antigens employed and species under investigation [83,84].
4. Surveillance in Humans and Animals
According to the Commission Implementing Regulation (EU) 2018/1882, RVF is categorized as a Category A disease [85] and a notifiable disease, as it has been designated by the World Organisation for Animal Health (WOAH). To date, there is no EU legislation or diagnostic manual for the early detection or surveillance with respect to RVF (EFSA). Except for the outbreaks involving multiple human cases in a French overseas department (Mayotte) in 2018–2019, RVF has never been reported in continental Europe or in countries neighboring the European Union.
According to EFSA opinion [86], because of the possible risk of introduction of RVFV into the EU through vector imports (ports, airports, cargo, and container yards), cattle and small ruminant herds should be included in a surveillance system.
Specifically, passive surveillance can be considered as a first choice for early detection of infection during the peak and late vector season. This should be conducted in areas with the highest risk of introduction (EFSA) in cases of reported animal abortions, stillbirths, or neonatal mortality of cattle, sheep, and goats.
Infected hosts can function as the initial amplifying host, but importation of animals is under restrictions of international trade and checked by the veterinary system. RVFV can be spread to a new geographic area via the movement of infected vectors that can be dispersed via wind, with an estimated travel distance ranging from 110 to 1350 km within 24 h [87], or mechanical transport. This last hypothesis is more likely to be the potential mode; numerous vectors have been discovered alive within aircraft and luggage after international flights [88]. In Kenya, an enhanced surveillance for RFV has been performed, preventing a possible spillover to humans, through a pilot model communication network for emergency reporting of animal health status between farmers, county government surveillance officers, and the national government [89]. Due to climate change, together with the globalization of the animal trade and the wide variety of mosquito species, vector surveillance is crucially important in detecting virus activity as early as possible for a rapid response to reduce an outbreak [90]. Using a model called MINTRISK (Method to INTegrate all relevant RISK aspects), the risk of the introduction of RVFV in the EU was estimated to be very low in all regions (less than one outbreak every 500 years) by the movements of both infected animals and/or vectors [38], but it is not excluded that animal movement may contribute to viral spread, threatening countries in Europe where competent vectors are present [43].
Nevertheless, the surveillance of EU countries should be strengthened due to the spread of the virus in the neighboring countries.
5. Discussion
To date, there is no validated point-of-care diagnostic tool. Rapid point-of-care tests would be critically important because they can be used at the patient’s bedside and would provide an alternative that does not require handling with clinical specimens and extraction of nucleic acids from samples. These rapid tests in this way should be easy to use but compatible with biosafety, inexpensive, and do not require well-trained personnel, making diagnosis faster.
Classical diagnosis employs mainly expensive and not so easy-to-use RT-PCR assays, which must be implemented on specific and expensive machines by personnel trained for this purpose, and there are few laboratories capable of performing serosurveillance in asymptomatic people [38,91].
There are few BSL-3 reference laboratories where samples can be managed, not always in the country of the epidemic. The fact that the virus can only be detected in the blood for a brief period (3–5 days post onset of the disease) suggests that RT-PCR alone is not sufficient for case determination. It is preferable to combine both RT-PCR and IgM ELISA or IFA (IgM has a 6-week window in the blood). However, the quality of in-house tests is frequently unknown, and there are few validated serology assays for human specimens. It is necessary to investigate the development of diagnostics intended to detect viruses in other body fluids. Before beginning vaccine clinical trials, laboratory capacity and surveillance systems must be strengthened, and diagnostic tests must be evaluated and validated. For test evaluation, having access to well-characterized samples and reference standards will be necessary (WHO, efficacy trials of Rift Valley fever vaccines and therapeutics) [30]. In 2019, a project (VHFMoDRAD) involving 13 partners started, with an end date set for 31 December 2023. The aim was to develop and deliver rapid and multiplex point-of-care diagnostic tools that will significantly increase the capacity to handle outbreaks of filoviruses and other viral hemorrhagic fever diseases in Africa, such as RVFV. In this project, Gregor Km et al. (2021) carried out a study where they observed how different antibodies can be useful for diagnosis and detection of RVFV in tissues from animals or mosquitoes. The authors found that antibodies against nucleoproteins can be a good tool to find infected cells in animals such as sheep and mice, and antibodies against glycoproteins can be used to find infected tissues in insects [92].
Finally, the development of multiplex detection systems serves numerous purposes, such as surveillance, monitoring the presence of RVFV in animals or humans, diagnosis, confirming RVFV infection, and research, studying the biology of RVFV and developing new vaccines and treatments. Despite some limitations, such as complexity, elevated cost and training requirements, compared to traditional diagnostic methods, multiplex detection systems offer several advantages: sensitivity, detecting smaller amounts of viral antigens; specificity, distinguishing between different pathogens; and rapidity, delivering results relatively quickly [64,65].
These findings bring us one step closer to the development of effective diagnostic tools.
6. Conclusions
Since 2019, the WHO has added RVFV to the Blueprint priority disease list. Surveillance activities and the rapid identification of positive cases are still important to prevent or limit the impact of human outbreaks, and diagnosis is a key factor for this success.
A plausible pathway for the introduction of RVFV in Europe, in particular for those countries where this can occur, is the movement of infected animals and vectors (shipped by air, sea container, or road transport) [38]. To mitigate the efforts in the risk areas, besides enhanced surveillance activities, there can be the imposition of animal movement restrictions/quarantines, the distribution of mosquito nets, and the dissemination of information to reduce human contact with infected animal products and vectors [91].
Author Contributions
Conceptualization, writing, draft, D.L., A.R.G. and S.T.; reviewing, D.L., S.T. and F.M.; data curation: S.P. and I.R.; resources: D.L. and S.T.; writing—original draft preparation: D.L., I.R., S.T. and S.P.; supervision: M.T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ricerca Corrente Linea 1, funded by the Italian Ministry of Health.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jonkmans, N.; D’Acremont, V.; Flahault, A. Scoping Future Outbreaks: A Scoping Review on the Outbreak Prediction of the WHO Blueprint List of Priority Diseases. BMJ Glob. Health 2021, 6, e006623. [Google Scholar] [CrossRef] [PubMed]
- Daubney, R.; Hudson, J.R.; Garnham, P.C. Enzootic Hepatitis or Rift Valley Fever. An Undescribed Virus Disease of Sheep Cattle and Man from East Africa. J. Pathol. Bacteriol. 1931, 34, 545–579. [Google Scholar] [CrossRef]
- Ikegami, T.; Makino, S. The Pathogenesis of Rift Valley Fever. Viruses 2011, 3, 493–519. [Google Scholar] [CrossRef] [PubMed]
- Gaudreault, N.N.; Indran, S.V.; Balaraman, V.; Wilson, W.C.; Richt, J.A. Molecular Aspects of Rift Valley Fever Virus and the Emergence of Reassortants. Virus Genes 2019, 55, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Saluzzo, J.F.; Smith, J.F. Use of Reassortant Viruses to Map Attenuating and Temperature-Sensitive Mutations of the Rift Valley Fever Virus MP-12 Vaccine. Vaccine 1990, 8, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Ly, H.J.; Lokugamage, N.; Nishiyama, S.; Ikegami, T. Risk Analysis of Inter-Species Reassortment through a Rift Valley Fever Phlebovirus MP-12 Vaccine Strain. PLoS ONE 2017, 12, e0185194. [Google Scholar] [CrossRef]
- Pepin, M.; Bouloy, M.; Bird, B.H.; Kemp, A.; Paweska, J. Rift Valley Fever Virus (Bunyaviridae: Phlebovirus): An Update on Pathogenesis, Molecular Epidemiology, Vectors, Diagnostics and Prevention. Vet. Res. 2010, 41, 61. [Google Scholar] [CrossRef]
- Wright, D.; Kortekaas, J.; Bowden, T.A.; Warimwe, G.M. Rift Valley Fever: Biology and Epidemiology. J. Gen. Virol. 2019, 100, 1187–1199. [Google Scholar] [CrossRef]
- Wright, D.; Allen, E.R.; Clark, M.H.A.; Gitonga, J.N.; Karanja, H.K.; Hulswit, R.J.G.; Taylor, I.; Biswas, S.; Marshall, J.; Mwololo, D.; et al. Naturally Acquired Rift Valley Fever Virus Neutralizing Antibodies Predominantly Target the Gn Glycoprotein. iScience 2020, 23, 101669. [Google Scholar] [CrossRef]
- Quellec, J.; Pédarrieu, A.; Piro-Mégy, C.; Barthelemy, J.; Simonin, Y.; Salinas, S.; Cêtre-Sossah, C. Rift Valley Fever Virus Modulates Apoptosis and Immune Response during Infection of Human Astrocytes. Emerg. Microbes Infect. 2023, 12, 2207672. [Google Scholar] [CrossRef]
- Wichgers Schreur, P.J.; Kortekaas, J. Single-Molecule FISH Reveals Non-Selective Packaging of Rift Valley Fever Virus Genome Segments. PLoS Pathog. 2016, 12, e1005800. [Google Scholar] [CrossRef]
- World Health Organization. Rift Valley Fever. Available online: https://www.who.int/news-room/fact-sheets/detail/rift-valley-fever (accessed on 28 December 2023).
- Alrajhi, A.A.; Al-Semari, A.; Al-Watban, J. Rift Valley Fever Encephalitis. Emerg. Infect. Dis. 2004, 10, 554–555. [Google Scholar] [CrossRef]
- Nayak, N.; Mishra, M. Drosophila Melanogaster as a Model to Understand the Mechanisms of Infection Mediated Neuroinflammation in Neurodegenerative Diseases. J. Integr. Neurosci. 2022, 21, 66. [Google Scholar] [CrossRef]
- Evans, A.; Gakuya, F.; Paweska, J.T.; Rostal, M.; Akoolo, L.; Van Vuren, P.J.; Manyibe, T.; Macharia, J.M.; Ksiazek, T.G.; Feikin, D.R.; et al. Prevalence of Antibodies against Rift Valley Fever Virus in Kenyan Wildlife. Epidemiol. Infect. 2008, 136, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Depner, K.; Drewe, J.A.; Garin-Bastuji, B.; Rojas, J.L.G.; Schmidt, C.G.; Michel, V.; et al. Rift Valley Fever—Epidemiological Update and Risk of Introduction into Europe. EFSA J. Eur. Food Saf. Auth. 2020, 18, e06041. [Google Scholar] [CrossRef] [PubMed]
- Hartman, A. Rift Valley Fever. Clin. Lab. Med. 2017, 37, 285–301. [Google Scholar] [CrossRef]
- Paweska, J.T. Rift Valley Fever. Rev. Sci. Tech. OIE 2015, 34, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Davies, F.G. The Historical and Recent Impact of Rift Valley Fever in Africa. Am. J. Trop. Med. Hyg. 2010, 83, 73–74. [Google Scholar] [CrossRef]
- Lumley, S.; Horton, D.L.; Hernandez-Triana, L.L.M.; Johnson, N.; Fooks, A.R.; Hewson, R. Rift Valley Fever Virus: Strategies for Maintenance, Survival and Vertical Transmission in Mosquitoes. J. Gen. Virol. 2017, 98, 875–887. [Google Scholar] [CrossRef]
- Linthicum, K.J.; Britch, S.C.; Anyamba, A. Rift Valley Fever: An Emerging Mosquito-Borne Disease. Annu. Rev. Entomol. 2016, 61, 395–415. [Google Scholar] [CrossRef]
- Ratovonjato, J.; Olive, M.-M.; Tantely, L.M.; Andrianaivolambo, L.; Tata, E.; Razainirina, J.; Jeanmaire, E.; Reynes, J.-M.; Elissa, N. Detection, Isolation, and Genetic Characterization of Rift Valley Fever Virus from Anopheles (Anopheles) Coustani, Anopheles (Anopheles) Squamosus, and Culex (Culex) Antennatus of the Haute Matsiatra Region, Madagascar. Vector-Borne Zoonotic Dis. 2011, 11, 753–759. [Google Scholar] [CrossRef]
- Sang, R.C.; Ahmed, O.; Faye, O.; Kelly, C.L.H.; Yahaya, A.A.; Mmadi, I.; Toilibou, A.; Sergon, K.; Brown, J.; Agata, N.; et al. Entomologic Investigations of a Chikungunya Virus Epidemic in the Union of the Comoros, 2005. Am. J. Trop. Med. Hyg. 2008, 78, 77–82. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Rift Valley Fever Surveillance. Available online: https://www.fao.org/3/i8475en/I8475EN.pdf (accessed on 18 November 2023).
- Gerdes, G.H. Rift Valley Fever. Rev. Sci. Tech. OIE 2004, 23, 613–623. [Google Scholar] [CrossRef]
- Adam, I.; Karsany, M.S. Case Report: Rift Valley Fever with Vertical Transmission in a Pregnant Sudanese Woman. J. Med. Virol. 2008, 80, 929. [Google Scholar] [CrossRef]
- Antonis, A.F.G.; Kortekaas, J.; Kant, J.; Vloet, R.P.M.; Vogel-Brink, A.; Stockhofe, N.; Moormann, R.J.M. Vertical Transmission of Rift Valley Fever Virus Without Detectable Maternal Viremia. Vector-Borne Zoonotic Dis. 2013, 13, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Soti, V.; Tran, A.; Degenne, P.; Chevalier, V.; Lo Seen, D.; Thiongane, Y.; Diallo, M.; Guégan, J.-F.; Fontenille, D. Combining Hydrology and Mosquito Population Models to Identify the Drivers of Rift Valley Fever Emergence in Semi-Arid Regions of West Africa. PLoS Negl. Trop. Dis. 2012, 6, e1795. [Google Scholar] [CrossRef]
- Caminade, C.; Ndione, J.; Diallo, M.; MacLeod, D.; Faye, O.; Ba, Y.; Dia, I.; Morse, A. Rift Valley Fever Outbreaks in Mauritania and Related Environmental Conditions. Int. J. Environ. Res. Public Health 2014, 11, 903–918. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Efficacy Trials of Rift Valley Fever Vaccines and Therapeutics. Guidance on Clinical Trial Design. Meeting Report. 2019. Available online: https://cdn.who.int/media/docs/default-source/blue-print/rift-valley-fever-blueprint-trial-design-meeting-report-2019.pdf?sfvrsn=ee74e0fb_3 (accessed on 5 December 2023).
- European Centre for Disease Prevention and Control. Facts about Rift Valley Fever. Available online: https://www.ecdc.europa.eu/en/rift-valley-fever/facts (accessed on 25 August 2023).
- Grossi-Soyster, E.N.; Banda, T.; Teng, C.Y.; Muchiri, E.M.; Mungai, P.L.; Mutuku, F.M.; Gildengorin, G.; Kitron, U.; King, C.H.; Desiree Labeaud, A. Rift Valley Fever Seroprevalence in Coastal Kenya. Am. J. Trop. Med. Hyg. 2017, 97, 115–120. [Google Scholar] [CrossRef]
- Memish, Z.A.; Masri, M.A.; Anderson, B.D.; Heil, G.L.; Merrill, H.R.; Khan, S.U.; Alsahly, A.; Gray, G.C. Elevated Antibodies against Rift Valley Fever Virus among Humans with Exposure to Ruminants in Saudi Arabia. Am. J. Trop. Med. Hyg. 2015, 92, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Bosworth, A.; Ghabbari, T.; Dowall, S.; Varghese, A.; Fares, W.; Hewson, R.; Zhioua, E.; Chakroun, M.; Tiouiri, H.; Ben Jemaa, M.; et al. Serologic Evidence of Exposure to Rift Valley Fever Virus Detected in Tunisia. New Microbes New Infect. 2016, 9, 1–7. [Google Scholar] [CrossRef]
- Tezcan-Ulger, S.; Kurnaz, N.; Ulger, M.; Aslan, G.; Emekdas, G. Serological Evidence of Rift Valley Fever Virus among Humans in Mersin Province of Turkey. J. Vector Borne Dis. 2019, 56, 373–379. [Google Scholar] [CrossRef]
- Lapa, D.; Specchiarello, E.; Francalancia, M.; Girardi, E.; Maggi, F.; Garbuglia, A.R. Detection of Anti-Rift Valley Fever Virus Antibodies in Serum Samples of Patients with Suspected Arbovirus Infection. Microorganisms 2023, 11, 2081. [Google Scholar] [CrossRef]
- Britch, S.C.; Binepal, Y.S.; Ruder, M.G.; Kariithi, H.M.; Linthicum, K.J.; Anyamba, A.; Small, J.L.; Tucker, C.J.; Ateya, L.O.; Oriko, A.A.; et al. Rift Valley Fever Risk Map Model and Seroprevalence in Selected Wild Ungulates and Camels from Kenya. PLoS ONE 2013, 8, e66626. [Google Scholar] [CrossRef]
- Kwaśnik, M.; Rożek, W.; Rola, J. Rift Valley Fever—A Growing Threat to Humans and Animals. J. Vet. Res. 2021, 65, 7–14. [Google Scholar] [CrossRef]
- Zouaghi, K.; Bouattour, A.; Aounallah, H.; Surtees, R.; Krause, E.; Michel, J.; Mamlouk, A.; Nitsche, A.; M’ghirbi, Y. First Serological Evidence of Crimean-Congo Hemorrhagic Fever Virus and Rift Valley Fever Virus in Ruminants in Tunisia. Pathogens 2021, 10, 769. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, T.; Boulianne, C.; Vincent, M.J.; Pezzanite, L.; Al-Qahtani, M.M.; Al-Mazrou, Y.; Khan, A.S.; Rollin, P.E.; Swanepoel, R.; Ksiazek, T.G.; et al. Genetic Analysis of Viruses Associated with Emergence of Rift Valley Fever in Saudi Arabia and Yemen, 2000–2001. Emerg. Infect. Dis. 2002, 8, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (eCDC). Rift Valley Fever. Annual Epidemiological Report for 2019; ECDC: Stockholm, Sweden, 2021; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/AER-Rift-Valley-fever-2019_0.pdf (accessed on 10 February 2024).
- Gossner, C.M.; Hallmaier-Wacker, L.; Briet, O.; Haussig, J.M.; de Valk, H.; Wijermans, A.; Bakonyi, T.; Madubuko, T.; Frank, C.; Noel, H.; et al. Arthropod-Borne Diseases among Travellers Arriving in Europe from Africa, 2015 to 2019. Euro Surveill. Bull. Eur. Sur Mal. Transm. Eur. Commun. Dis. Bull. 2023, 28, 2200270. [Google Scholar] [CrossRef]
- Chevalier, V. Relevance of Rift Valley Fever to Public Health in the European Union. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2013, 19, 705–708. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). RVF Distribution Map. Available online: https://www.cdc.gov/vhf/rvf/outbreaks/distribution-map.html#print (accessed on 14 January 2024).
- Sall, A.A.; Thonnon, J.; Sene, O.K.; Fall, A.; Ndiaye, M.; Baudez, B.; Mathiot, C.; Bouloy, M. Single-Tube and Nested Reverse Transcriptase-Polymerase Chain Reaction for Detection of Rift Valley Fever Virus in Human and Animal Sera. J. Virol. Methods 2001, 91, 85–92. [Google Scholar] [CrossRef]
- Garcia, S.; Crance, J.M.; Billecocq, A.; Peinnequin, A.; Jouan, A.; Bouloy, M.; Garin, D. Quantitative Real-Time PCR Detection of Rift Valley Fever Virus and Its Application to Evaluation of Antiviral Compounds. J. Clin. Microbiol. 2001, 39, 4456–4461. [Google Scholar] [CrossRef][Green Version]
- Weidmann, M.; Sanchez-Seco, M.P.; Sall, A.A.; Ly, P.O.; Thiongane, Y.; Lô, M.M.; Schley, H.; Hufert, F.T. Rapid Detection of Important Human Pathogenic Phleboviruses. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2008, 41, 138–142. [Google Scholar] [CrossRef]
- Drosten, C.; Göttig, S.; Schilling, S.; Asper, M.; Panning, M.; Schmitz, H.; Günther, S. Rapid Detection and Quantification of RNA of Ebola and Marburg Viruses, Lassa Virus, Crimean-Congo Hemorrhagic Fever Virus, Rift Valley Fever Virus, Dengue Virus, and Yellow Fever Virus by Real-Time Reverse Transcription-PCR. J. Clin. Microbiol. 2002, 40, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Njenga, M.K.; Paweska, J.; Wanjala, R.; Rao, C.Y.; Weiner, M.; Omballa, V.; Luman, E.T.; Mutonga, D.; Sharif, S.; Panning, M.; et al. Using a Field Quantitative Real-Time PCR Test to Rapidly Identify Highly Viremic Rift Valley Fever Cases. J. Clin. Microbiol. 2009, 47, 1166–1171. [Google Scholar] [CrossRef]
- Peyrefitte, C.N.; Boubis, L.; Coudrier, D.; Bouloy, M.; Grandadam, M.; Tolou, H.J.; Plumet, S. Real-Time Reverse-Transcription Loop-Mediated Isothermal Amplification for Rapid Detection of Rift Valley Fever Virus. J. Clin. Microbiol. 2008, 46, 3653–3659. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, C.A.; Kubo, T.; Grobbelaar, A.A.; van Vuren, P.J.; Weyer, J.; Nel, L.H.; Swanepoel, R.; Morita, K.; Paweska, J.T. Development and Evaluation of a Real-Time Reverse Transcription-Loop-Mediated Isothermal Amplification Assay for Rapid Detection of Rift Valley Fever Virus in Clinical Specimens. J. Clin. Microbiol. 2009, 47, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Zhang, S.; Liu, D.; Yan, F.; Wang, H.; Huang, P.; Bi, J.; Jin, H.; Feng, N.; Cao, Z.; et al. Development of a Visible Reverse Transcription-Loop-Mediated Isothermal Amplification Assay for the Detection of Rift Valley Fever Virus. Front. Microbiol. 2020, 11, 590732. [Google Scholar] [CrossRef]
- Euler, M.; Wang, Y.; Nentwich, O.; Piepenburg, O.; Hufert, F.T.; Weidmann, M. Recombinase Polymerase Amplification Assay for Rapid Detection of Rift Valley Fever Virus. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2012, 54, 308–312. [Google Scholar] [CrossRef]
- Wekesa, F.; Wamalwa, M.; Oduor, R.; Binepal, Y.; Ateya, L.; Okumu, N.; M’kwenda, A.; Masaba, C.; Mukhaye, E. Development and Validation of Rapid Colorimetric Reverse Transcription Loop-Mediated Isothermal Amplification for Detection of Rift Valley Fever Virus. Adv. Virol. 2023, 2023, 1863980. [Google Scholar] [CrossRef]
- He, J.; Kraft, A.J.; Fan, J.; Van Dyke, M.; Wang, L.; Bose, M.E.; Khanna, M.; Metallo, J.A.; Henrickson, K.J. Simultaneous Detection of CDC Category “A” DNA and RNA Bioterrorism Agents by Use of Multiplex PCR & RT-PCR Enzyme Hybridization Assays. Viruses 2009, 1, 441–459. [Google Scholar] [CrossRef]
- Balaraman, V.; Gaudreault, N.N.; Trujillo, J.D.; Indran, S.V.; Wilson, W.C.; Richt, J.A. RT-qPCR Genotyping Assays for Differentiating Rift Valley Fever Phlebovirus Strains. J. Virol. Methods 2023, 315, 114693. [Google Scholar] [CrossRef]
- Yao, W.; Yang, Z.; Lou, X.; Mao, H.; Yan, H.; Zhang, Y. Simultaneous Detection of Ebola Virus and Pathogens Associated with Hemorrhagic Fever by an Oligonucleotide Microarray. Front. Microbiol. 2021, 12, 713372. [Google Scholar] [CrossRef]
- Venter, M.; Zaayman, D.; Van Niekerk, S.; Stivaktas, V.; Goolab, S.; Weyer, J.; Paweska, J.T.; Swanepoel, R. Macroarray Assay for Differential Diagnosis of Meningoencephalitis in Southern Africa. J. Clin. Virol. 2014, 60, 50–56. [Google Scholar] [CrossRef]
- Sardi, S.I.; Somasekar, S.; Naccache, S.N.; Bandeira, A.C.; Tauro, L.B.; Campos, G.S.; Chiu, C.Y. Coinfections of Zika and Chikungunya Viruses in Bahia, Brazil, Identified by Metagenomic Next-Generation Sequencing. J. Clin. Microbiol. 2016, 54, 2348–2353. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.R.; Zimmermann, L.L.; Crawford, E.D.; Sample, H.A.; Soni, P.R.; Baker, A.N.; Khan, L.M.; DeRisi, J.L. Acute West Nile Virus Meningoencephalitis Diagnosed Via Metagenomic Deep Sequencing of Cerebrospinal Fluid in a Renal Transplant Patient. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2017, 17, 803–808. [Google Scholar] [CrossRef]
- Altona Diagnostics. RealStar®® Rift Valley Fever Virus RT-PCR Kit 1.0. Available online: https://www.altona-diagnostics.com/files/public/Content%20Homepage/-%2002%20RealStar/MAN%20-%20CE%20-%20EN/RealStar%20RVFV%20RT-PCR%20Kit%201.0_WEB_CE_EN-S02.pdf (accessed on 15 January 2024).
- Petrova, V.; Kristiansen, P.; Norheim, G.; Yimer, S.A. Rift Valley Fever: Diagnostic Challenges and Investment Needs for Vaccine Development. BMJ Glob. Health 2020, 5, e002694. [Google Scholar] [CrossRef]
- WOAH (World Organisation for Animal Health). Report of the Meeting of the OIE Biological Standards Commission; World Organisation for Animal Health: Paris, France, 2013; Available online: https://www.woah.org/fileadmin/Home/eng/Internationa_Standard_Setting/docs/pdf/BSC/A_BSC_Feb2013.pdf (accessed on 17 January 2024).
- Hoste, A.C.R.; Ruiz, T.; Fernández-Pacheco, P.; Jiménez-Clavero, M.Á.; Djadjovski, I.; Moreno, S.; Brun, A.; Edwards, T.A.; Barr, J.N.; Rueda, P.; et al. Development of a Multiplex Assay for Antibody Detection in Serum against Pathogens Affecting Ruminants. Transbound. Emerg. Dis. 2021, 68, 1229–1239. [Google Scholar] [CrossRef]
- Surtees, R.; Stern, D.; Ahrens, K.; Kromarek, N.; Lander, A.; Kreher, P.; Weiss, S.; Hewson, R.; Punch, E.K.; Barr, J.N.; et al. Development of a Multiplex Microsphere Immunoassay for the Detection of Antibodies against Highly Pathogenic Viruses in Human and Animal Serum Samples. PLoS Negl. Trop. Dis. 2020, 14, e0008699. [Google Scholar] [CrossRef]
- Ragan, I.K.; Schuck, K.N.; Upreti, D.; Odendaal, L.; Richt, J.A.; Trujillo, J.D.; Wilson, W.C.; Davis, A.S. Rift Valley Fever Viral RNA Detection by In Situ Hybridization in Formalin-Fixed, Paraffin-Embedded Tissues. Vector Borne Zoonotic Dis. 2019, 19, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Peters, C.J.; Meegan, J.M. Studies on the Antigenic Relationship among Phleboviruses. Am. J. Trop. Med. Hyg. 1982, 31, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Liu, D.; Nunes, M.R.T.; DA Rosa, A.P.A.T.; Tesh, R.B.; Xiao, S.-Y. Antigenic and Genetic Relationships among Rift Valley Fever Virus and Other Selected Members of the Genus Phlebovirus (Bunyaviridae). Am. J. Trop. Med. Hyg. 2007, 76, 1194–1200. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, Twelfth Edition 2023. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/A_summry.htm (accessed on 20 December 2023).
- Smith, M.R.; Schirtzinger, E.E.; Wilson, W.C.; Davis, A.S. Rift Valley Fever Virus: Propagation, Quantification, and Storage. Curr. Protoc. Microbiol. 2019, 55, e92. [Google Scholar] [CrossRef]
- Anderson, G.W.; Saluzzo, J.F.; Ksiazek, T.G.; Smith, J.F.; Ennis, W.; Thureen, D.; Peters, C.J.; Digoutte, J.P. Comparison of in Vitro and in Vivo Systems for Propagation of Rift Valley Fever Virus from Clinical Specimens. Res. Virol. 1989, 140, 129–138. [Google Scholar] [CrossRef]
- Odendaal, L.; Clift, S.J.; Fosgate, G.T.; Davis, A.S. Lesions and Cellular Tropism of Natural Rift Valley Fever Virus Infection in Adult Sheep. Vet. Pathol. 2019, 56, 61–77. [Google Scholar] [CrossRef]
- Digoutte, J.P.; Jouan, A.; Le Guenno, B.; Riou, O.; Philippe, B.; Meegan, J.; Ksiazek, T.G.; Peters, C.J. Isolation of the Rift Valley Fever Virus by Inoculation into Aedes Pseudoscutellaris Cells: Comparison with Other Diagnostic Methods. Res. Virol. 1989, 140, 31–41. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health (OIE). Rift Valley Fever (Infection with Rift Valley Fever Virus). Available online: https://www.woah.org/app/uploads/2021/03/3-01-18-rvf-1.pdf (accessed on 15 January 2024).
- Euroimmun. IFA per Infettivologia. Available online: https://www.euroimmun.it/prodotti/infettivologia/ifa-2/ (accessed on 1 November 2023).
- Paweska, J.T.; Smith, S.J.; Wright, I.M.; Williams, R.; Cohen, A.S.; Van Dijk, A.A.; Grobbelaar, A.A.; Croft, J.E.; Swanepoel, R.; Gerdes, G.H. Indirect Enzyme-Linked Immunosorbent Assay for the Detection of Antibody against Rift Valley Fever Virus in Domestic and Wild Ruminant Sera. Onderstepoort J. Vet. Res. 2003, 70, 49–64. [Google Scholar] [PubMed]
- Jansen van Vuren, P.; Potgieter, A.C.; Paweska, J.T.; van Dijk, A.A. Preparation and Evaluation of a Recombinant Rift Valley Fever Virus N Protein for the Detection of IgG and IgM Antibodies in Humans and Animals by Indirect ELISA. J. Virol. Methods 2007, 140, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Fafetine, J.M.; Tijhaar, E.; Paweska, J.T.; Neves, L.C.B.G.; Hendriks, J.; Swanepoel, R.; Coetzer, J.a.W.; Egberink, H.F.; Rutten, V.P.M.G. Cloning and Expression of Rift Valley Fever Virus Nucleocapsid (N) Protein and Evaluation of a N-Protein Based Indirect ELISA for the Detection of Specific IgG and IgM Antibodies in Domestic Ruminants. Vet. Microbiol. 2007, 121, 29–38. [Google Scholar] [CrossRef]
- Cêtre-Sossah, C.; Billecocq, A.; Lancelot, R.; Defernez, C.; Favre, J.; Bouloy, M.; Martinez, D.; Albina, E. Evaluation of a Commercial Competitive ELISA for the Detection of Antibodies to Rift Valley Fever Virus in Sera of Domestic Ruminants in France. Prev. Vet. Med. 2009, 90, 146–149. [Google Scholar] [CrossRef]
- van Vuren, P.J.; Paweska, J.T. Comparison of Enzyme-Linked Immunosorbent Assay-Based Techniques for the Detection of Antibody to Rift Valley Fever Virus in Thermochemically Inactivated Sheep Sera. Vector Borne Zoonotic Dis. 2010, 10, 697–699. [Google Scholar] [CrossRef]
- Fafetine, J.M.; Jansen van Vuren, P.; Paweska, J.T. Comparison of a Recombinant Nucleocapsid IgG Indirect ELISA with an IgG Sandwich ELISA for the Detection of Antibodies to Rift Valley Fever Virus in Small Ruminants. Vector Borne Zoonotic Dis. 2012, 12, 1062–1064. [Google Scholar] [CrossRef]
- Kortekaas, J.; Kant, J.; Vloet, R.; Cêtre-Sossah, C.; Marianneau, P.; Lacote, S.; Banyard, A.C.; Jeffries, C.; Eiden, M.; Groschup, M.; et al. European Ring Trial to Evaluate ELISAs for the Diagnosis of Infection with Rift Valley Fever Virus. J. Virol. Methods 2013, 187, 177–181. [Google Scholar] [CrossRef]
- Paweska, J.T.; Burt, F.J.; Swanepoel, R. Validation of IgG-Sandwich and IgM-Capture ELISA for the Detection of Antibody to Rift Valley Fever Virus in Humans. J. Virol. Methods 2005, 124, 173–181. [Google Scholar] [CrossRef]
- Faburay, B.; Wilson, W.C.; Secka, A.; Drolet, B.; McVey, D.S.; Richt, J.A. Evaluation of an Indirect Enzyme-Linked Immunosorbent Assay Based on Recombinant Baculovirus-Expressed Rift Valley Fever Virus Nucleoprotein as the Diagnostic Antigen. J. Clin. Microbiol. 2019, 57, e01058-19. [Google Scholar] [CrossRef]
- UNEP. Law and Environment Assistance Platform. Commission Implementing Regulation (EU) 2018/1882 on the Application of Certain Disease Prevention and Control Rules to Categories of Listed Diseases and Establishing a List of Species and Groups of Species Posing a Considerable Risk for the Spread of Those Listed Diseases. Available online: https://leap.unep.org/en/countries/eu/national-legislation/commission-implementing-regulation-eu-20181882-application (accessed on 14 August 2023).
- EFSA Panel on Animal Health and Welfare (EFSA AHAW Panel); Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Depner, K.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortázar Schmidt, C.; et al. Rift Valley Fever—Assessment of Effectiveness of Surveillance and Control Measures in the EU. EFSA J. Eur. Food Saf. Auth. 2020, 18, e06292. [Google Scholar] [CrossRef]
- Kasari, T.R.; Carr, D.A.; Lynn, T.V.; Weaver, J.T. Evaluation of Pathways for Release of Rift Valley Fever Virus into Domestic Ruminant Livestock, Ruminant Wildlife, and Human Populations in the Continental United States. J. Am. Vet. Med. Assoc. 2008, 232, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Rolin, A.I.; Berrang-Ford, L.; Kulkarni, M.A. The Risk of Rift Valley Fever Virus Introduction and Establishment in the United States and European Union. Emerg. Microbes Infect. 2013, 2, e81. [Google Scholar] [CrossRef]
- Oyas, H.; Holmstrom, L.; Kemunto, N.P.; Muturi, M.; Mwatondo, A.; Osoro, E.; Bitek, A.; Bett, B.; Githinji, J.W.; Thumbi, S.M.; et al. Enhanced Surveillance for Rift Valley Fever in Livestock during El Niño Rains and Threat of RVF Outbreak, Kenya, 2015–2016. PLoS Negl. Trop. Dis. 2018, 12, e0006353. [Google Scholar] [CrossRef]
- Kim, H.-J.; Lyoo, H.-R.; Park, J.-Y.; Choi, J.-S.; Lee, J.-Y.; Jeoung, H.-Y.; Cho, Y.-S.; Cho, I.-S.; Yoo, H.S. Surveillance of Rift Valley Fever Virus in Mosquito Vectors of the Republic of Korea. Vector Borne Zoonotic Dis. 2016, 16, 131–135. [Google Scholar] [CrossRef]
- Anyamba, A.; Linthicum, K.J.; Small, J.; Britch, S.C.; Pak, E.; de La Rocque, S.; Formenty, P.; Hightower, A.W.; Breiman, R.F.; Chretien, J.-P.; et al. Prediction, Assessment of the Rift Valley Fever Activity in East and Southern Africa 2006–2008 and Possible Vector Control Strategies. Am. J. Trop. Med. Hyg. 2010, 83, 43–51. [Google Scholar] [CrossRef]
- Gregor, K.M.; Michaely, L.M.; Gutjahr, B.; Rissmann, M.; Keller, M.; Dornbusch, S.; Naccache, F.; Schön, K.; Jansen, S.; Heitmann, A.; et al. Rift Valley Fever Virus Detection in Susceptible Hosts with Special Emphasis in Insects. Sci. Rep. 2021, 11, 9822. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).