

Analysis of Primary Chronic Lymphocytic Leukemia Cells’ Signaling Pathways

 , ,
, ,  , , ,
, , ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Donor Cells
2.2. Quantitative RT-PCR
2.3. Flow Cytometry
2.4. Western Blot Analysis
2.5. Cell Lines and Co-Culture Experiments
2.6. Statistics
3. Results
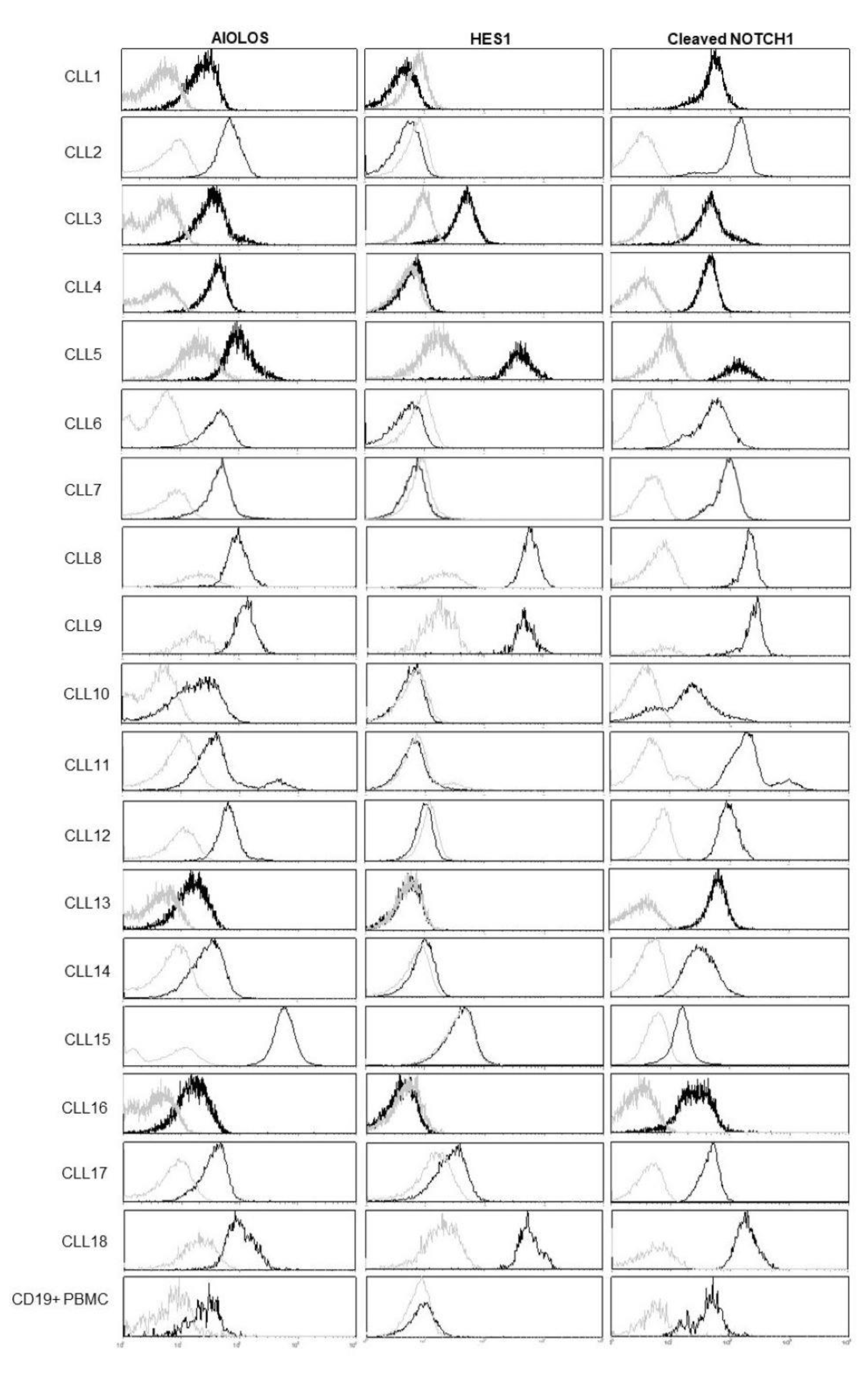
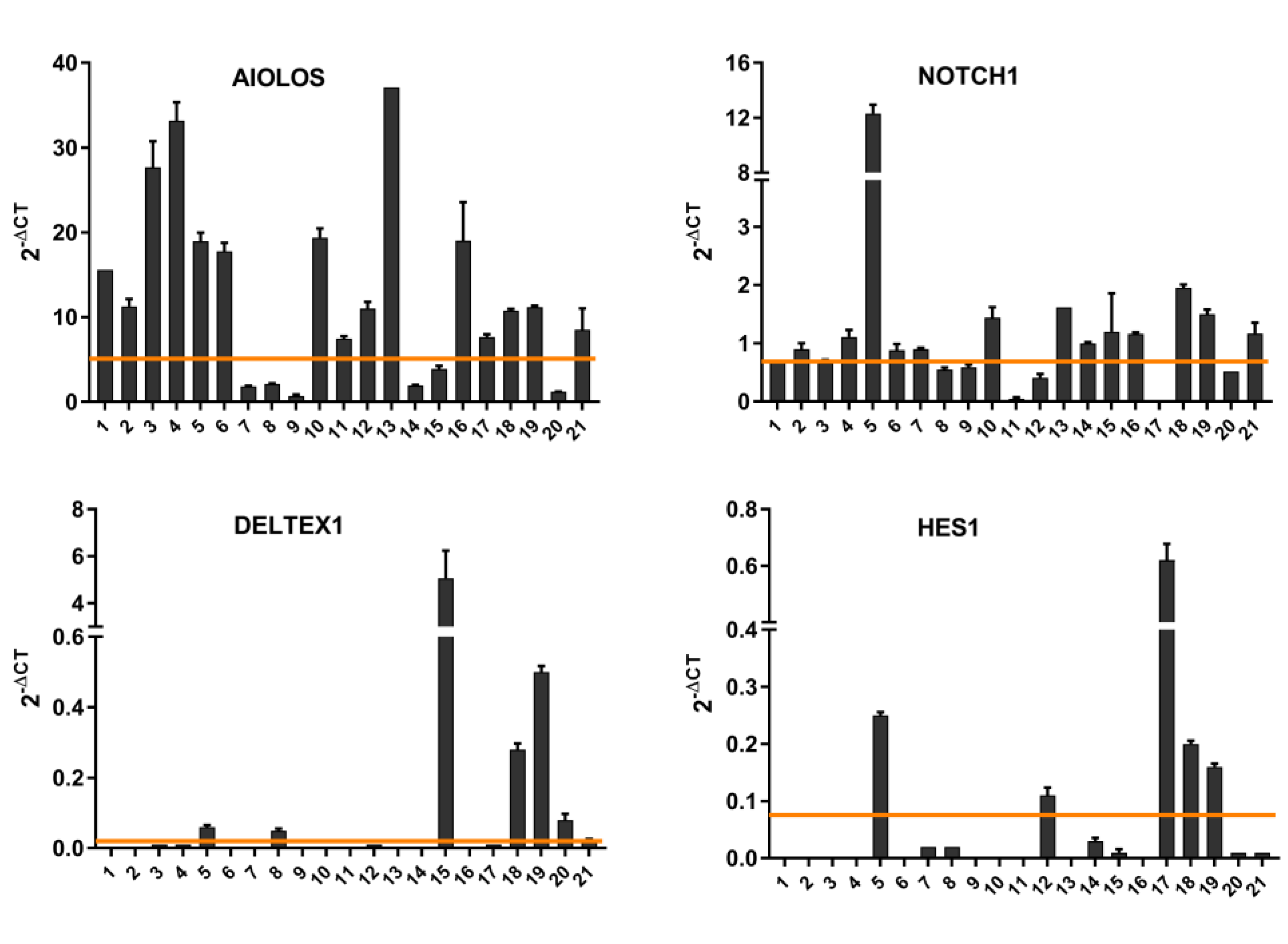
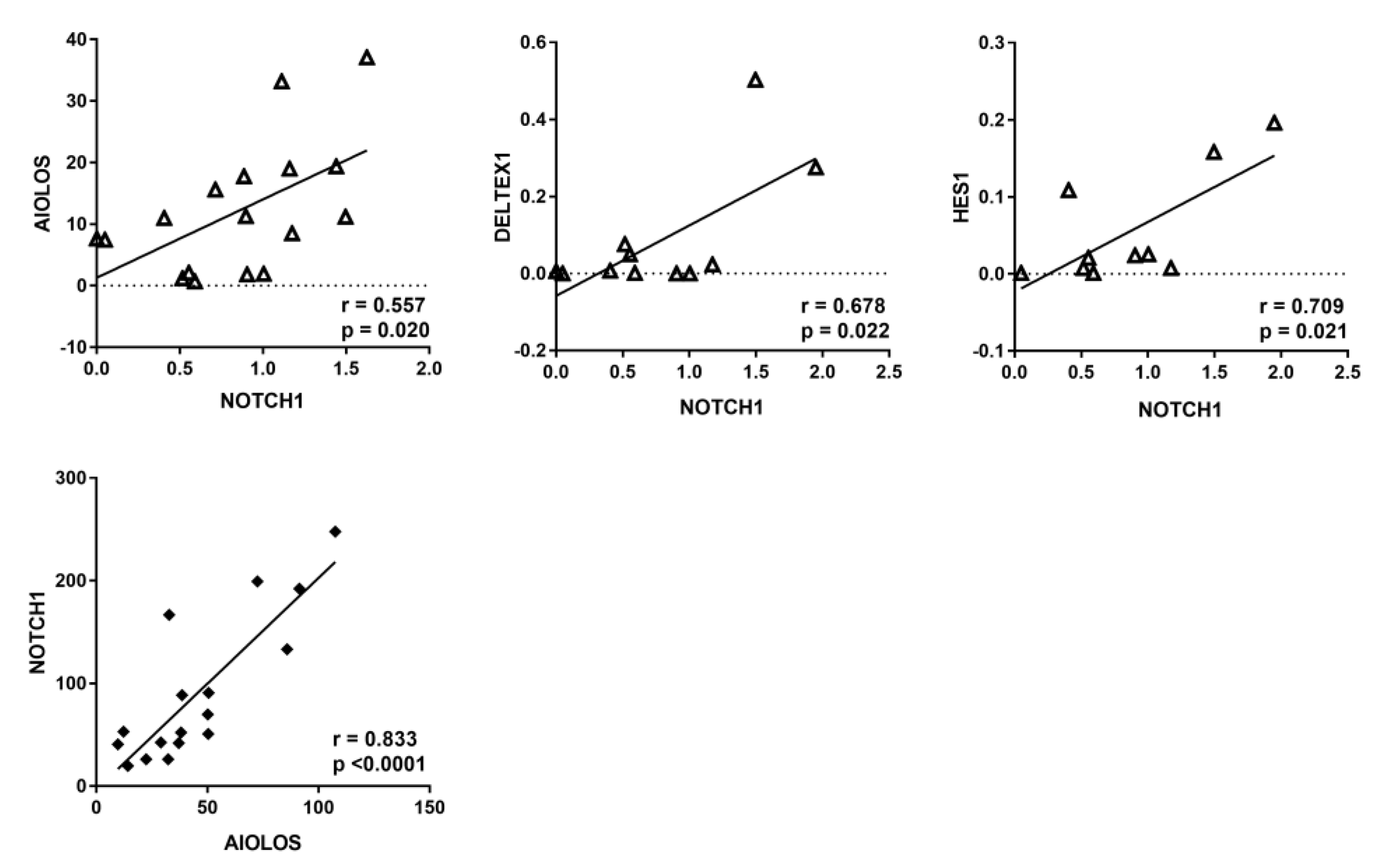
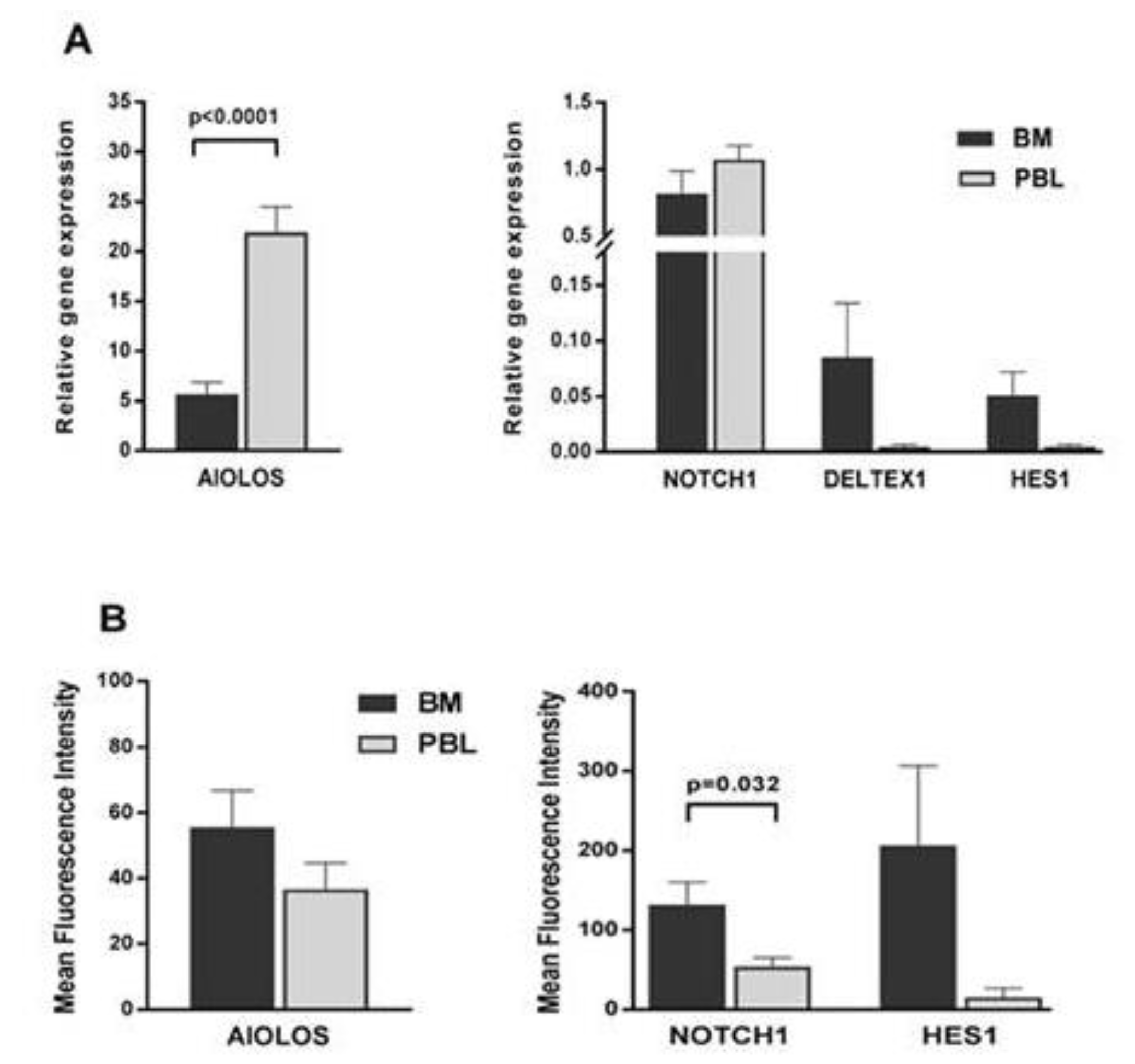
3.1. NOTCH1 Activation and AIOLOS Expression in CLL Patients
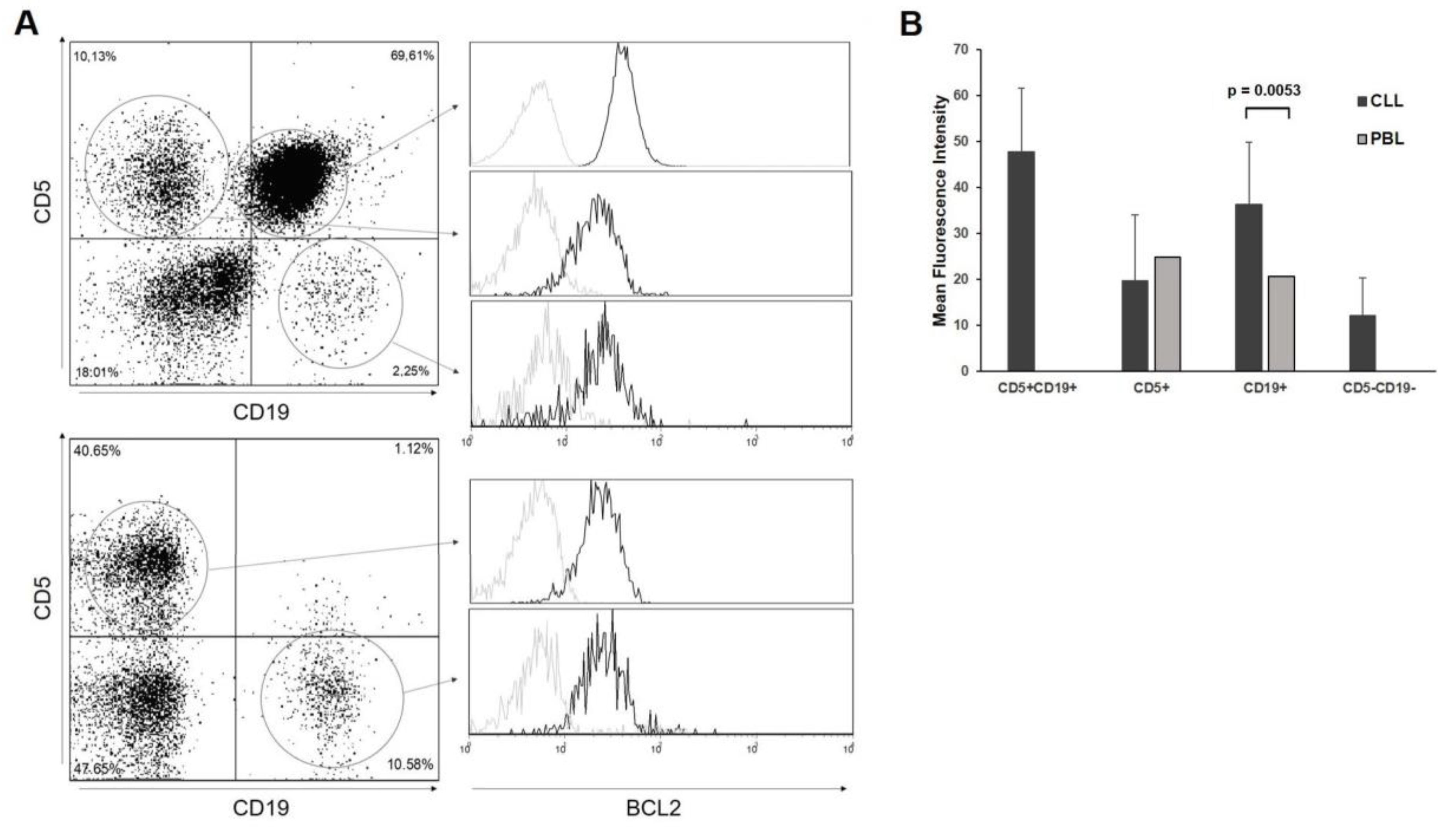
3.2. BCL2 Expression in CLL Patients
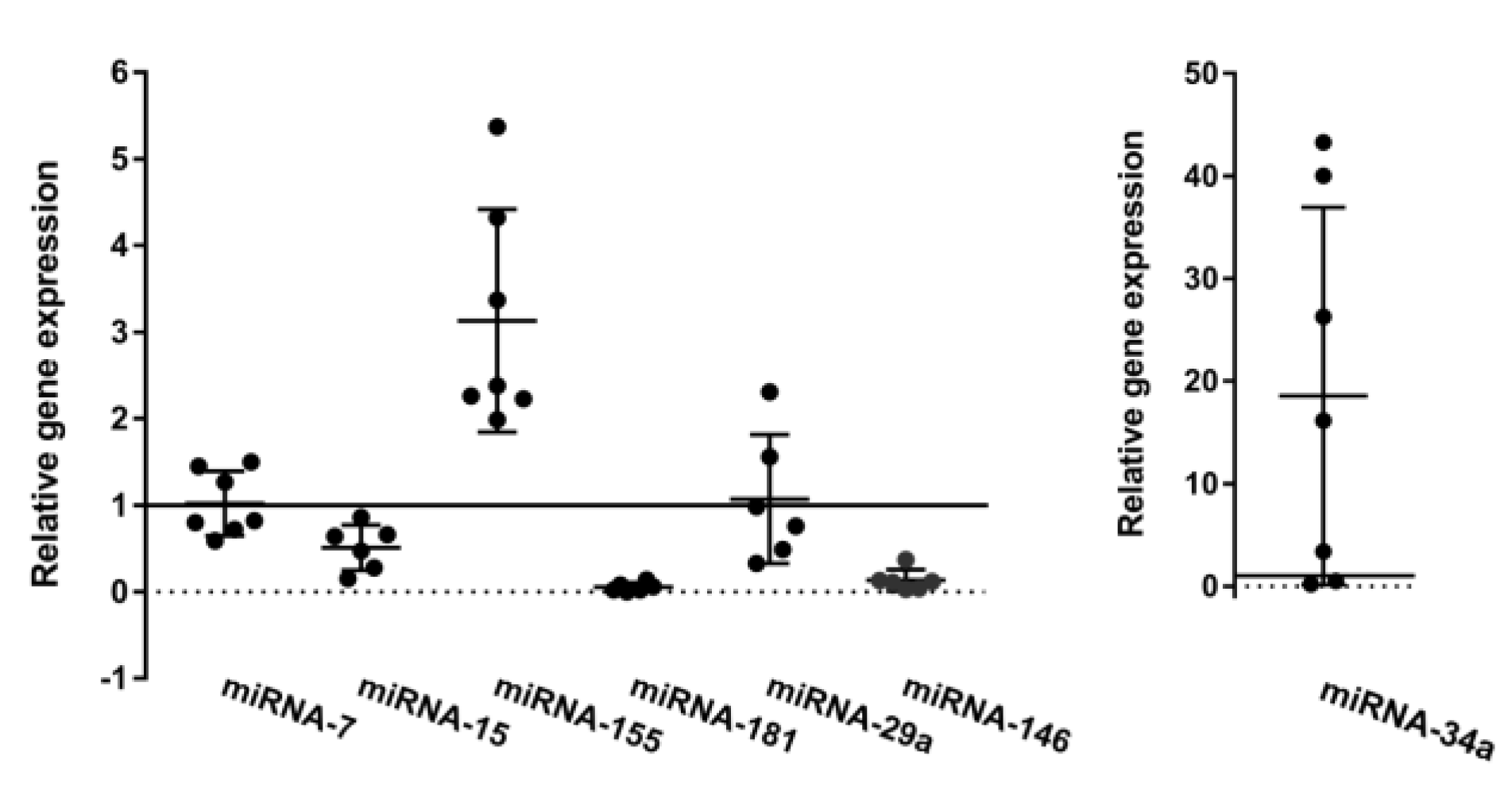
3.3. Oncogenic and Tumor Suppressor miRs Are Deregulated in CLL
3.4. Notch Pathway Activity in Bone Marrow and Peripheral Blood Samples
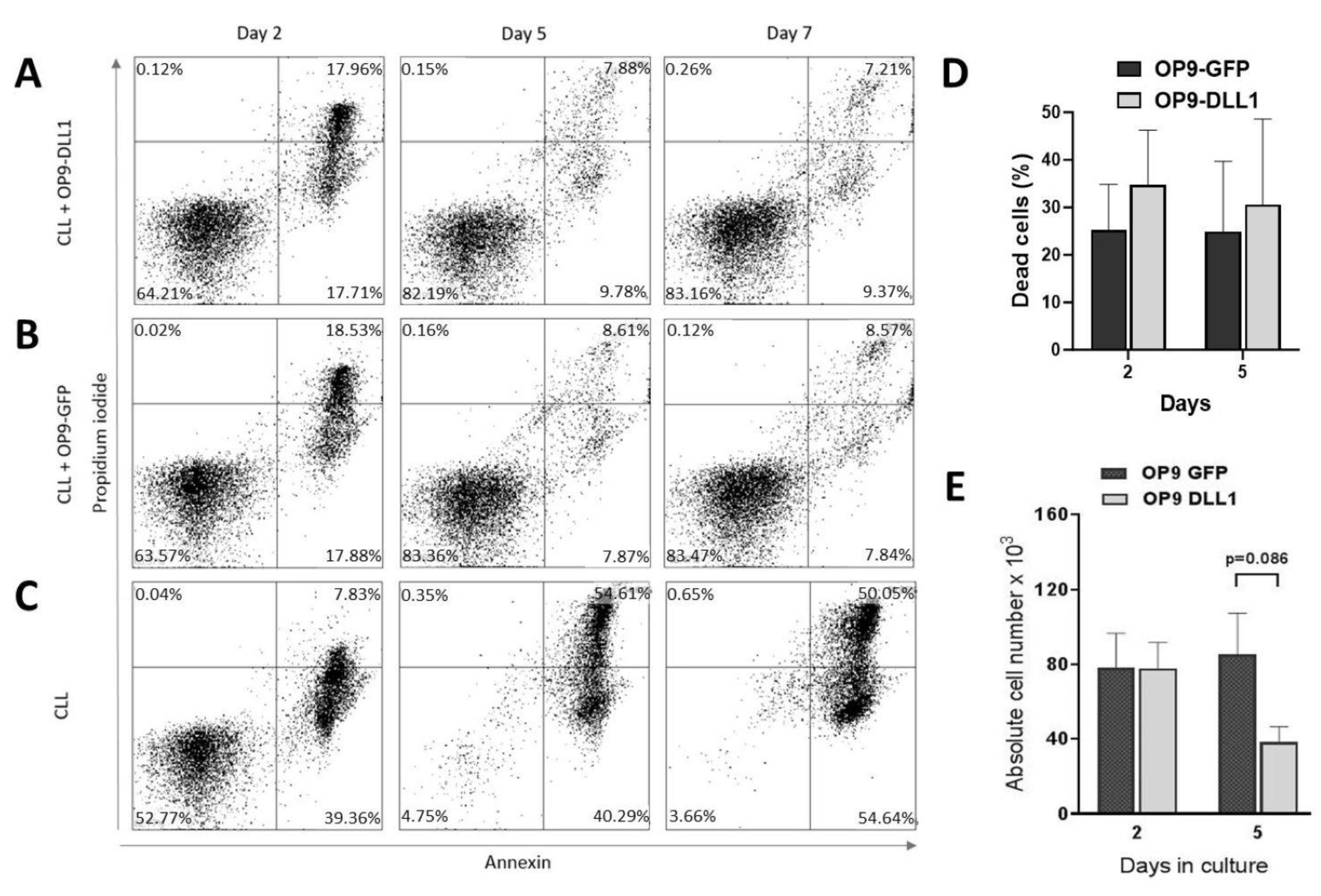
3.5. In Vitro CLL Cell Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hallek, M.; Al-Sawaf, O. Chronic lymphocytic leukemia: 2022 update on diagnostic and therapeutic procedures. Am. J. Hematol. 2021, 96, 1679–1705. [Google Scholar] [CrossRef]
- Caligaris-Cappio, F.; Ghia, P. The normal counterpart to the chronic lymphocytic leukemia B cell. Best Pract. Res. Clin. Haematol. 2007, 20, 385–397. [Google Scholar] [CrossRef]
- Scarfo, L.; Ferreri, A.J.M.; Ghia, P. Chronic lymphocytic leukaemia. Crit. Rev. Oncol. Hematol. 2016, 104, 169–182. [Google Scholar] [CrossRef]
- Kikushige, Y.; Ishikawa, F.; Miyamoto, T.; Shima, T.; Urata, S.; Yoshimoto, G.; Mori, Y.; Iino, T.; Yamauchi, T.; Eto, T.; et al. Self-renewing hematopoietic stem cell is the primary target in pathogenesis of human chronic lymphocytic leukemia. Cancer Cell 2011, 20, 246–259. [Google Scholar] [CrossRef]
- Rai, K.R.; Jain, P. Advances in the Clinical Staging of Chronic Lymphocytic Leukemia. Clin. Chem. 2011, 57, 1771–1772. [Google Scholar] [CrossRef]
- Rassenti, L.Z.; Jain, S.; Keating, M.J.; Wierda, W.G.; Grever, M.R.; Byrd, J.C.; Kay, N.E.; Brown, J.R.; Gribben, J.G.; Neuberg, D.S.; et al. Relative value of ZAP-70, CD38, and immunoglobulin mutation status in predicting aggressive disease in chronic lymphocytic leukemia. Blood 2008, 112, 1923–1930. [Google Scholar] [CrossRef]
- Zhou, B.; Lin, W.; Long, Y.; Yang, Y.; Zhang, H.; Wu, K.; Chu, Q. Notch signaling pathway: Architecture, disease, and therapeutics. Signal Transduct. Target Ther. 2022, 7, 95. [Google Scholar] [CrossRef]
- Ranganathan, P.; Weaver, K.L.; Capobianco, A.J. Notch signalling in solid tumours: A little bit of everything but not all the time. Nat. Rev. Cancer 2011, 11, 338–351. [Google Scholar] [CrossRef]
- Antica, M.; Cicin-Sain, L.; Kapitanovic, S.; Matulic, M.; Dzebro, S.; Dominis, M. Aberrant Ikaros, Aiolos, and Helios expression in Hodgkin and non-Hodgkin lymphoma. Blood 2008, 112, 452. [Google Scholar] [CrossRef]
- Billot, K.; Parizot, C.; Arrouss, I.; Mazier, D.; Debre, P.; Rogner, U.C.; Rebollo, A. Differential aiolos expression in human hematopoietic subpopulations. Leuk Res. 2010, 34, 289–293. [Google Scholar] [CrossRef]
- Rosati, E.; Sabatini, R.; Rampino, G.; Tabilio, A.; Di Ianni, M.; Fettucciari, K.; Bartoli, A.; Coaccioli, S.; Screpanti, I.; Marconi, P. Constitutively activated Notch signaling is involved in survival and apoptosis resistance of B-CLL cells Constitutively activated Notch signaling is involved in survival and apoptosis resistance of B-CLL cells. Blood 2009, 113, 856–865. [Google Scholar] [CrossRef]
- Rosati, E.; Sabatini, R.; De Falco, F.; Del Papa, B.; Falzetti, F.; Di Ianni, M.; Cavalli, L.; Fettucciari, K.; Bartoli, A.; Screpanti, I.; et al. γ-Secretase inhibitor I induces apoptosis in chronic lymphocytic leukemia cells by proteasome inhibition, endoplasmic reticulum stress increase and notch down-regulation. Int. J. Cancer 2013, 132, 1940–1953. [Google Scholar] [CrossRef] [PubMed]
- Villamor, N.; Conde, L.; Martínez-Trillos, A.; Cazorla, M.; Navarro, A.; Beà, S.; López, C.; Colomer, D.; Pinyol, M.; Aymerich, M.; et al. NOTCH1 mutations identify a genetic subgroup of chronic lymphocytic leukemia patients with high risk of transformation and poor outcome. Leukemia 2013, 27, 1100–1106. [Google Scholar] [CrossRef]
- Willander, K.; Dutta, R.K.; Ungerbäck, J.; Gunnarsson, R.; Juliusson, G.; Fredrikson, M.; Linderholm, M.; Soderkvist, P. NOTCH1 mutations influence survival in chronic lymphocytic leukemia patients. BMC Cancer 2013, 13, 274. [Google Scholar] [CrossRef]
- Kapoor, I.; Bodo, J.; Hill, B.T.; Hsi, E.D.; Almasan, A. Targeting BCL-2 in B-cell malignancies and overcoming therapeutic resistance. Cell Death Dis. 2020, 11, 941. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Cimmino, A.; Fabbri, M.; Ferracin, M.; Wojcik, S.E.; Shimizu, M.; Taccioli, C.; Zanesi, N.; Garzon, R.; Aqeilan, R.I.; et al. MiR-15a and miR-16-1 cluster functions in human leukemia. Proc. Nat. Acad. Sci. USA 2008, 105, 5166–5171. [Google Scholar] [CrossRef]
- Fabbri, G.; Rasi, S.; Rossi, D.; Trifonov, V.; Khiabanian, H.; Ma, J.; Grunn, A.; Fangazio, M.; Capello, D.; Monti, S.; et al. Analysis of the chronic lymphocytic leukemia coding genome: Role of NOTCH1 mutational activation. J. Exp. Med. 2011, 208, 1389–1401. [Google Scholar] [CrossRef]
- De Oliveira, T.D.; vom Stein, A.; Rebollido-Rios, R.; Lobastova, L.; Lettau, M.; Janssen, O.; Wagle, P.; Nguyen, P.-H.; Hallek, M.; Hansen, H.P. Stromal cells support the survival of human primary chronic lymphocytic leukemia (CLL) cells through Lyn-driven extracellular vesicles. Front. Med. 2023, 9, 1059028. [Google Scholar] [CrossRef] [PubMed]
- Pekarsky, Y.; Croce, C.M. Role of miR-15/16 in CLL. Cell Death Differ. 2015, 22, 6–11. [Google Scholar] [CrossRef]
- Khalid, K.; Padda, J.; Syam, M.; Moosa, A.; Kakani, V.; Sanka, S.; Zubair, U.; Padda, S.; Cooper, A.C.; Jean-Charles, G. 13q14 Deletion and Its Effect on Prognosis of Chronic Lymphocytic Leukemia. Cureus 2021, 2, 13. [Google Scholar] [CrossRef]
- Von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P.; STROBE Initiative. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. Bull. World Health Org. 2007, 85, 867–872. [Google Scholar] [CrossRef]
- Alaggio, R.; Amador, C.; Anagnostopoulos, I.; Attygalle, A.D.; Araujo, I.B.O.; Berti, E.; Bhagat, G.; Borges, A.M.; Boyer, D.; Calaminici, M.; et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Lymphoid Neoplasms. Leukemia 2022, 36, 1720–1748. [Google Scholar] [CrossRef] [PubMed]
- Jakšić, B.; Pejša, V.; Ostojić-Kolonić, S.; Kardum-Skelin, I.; Bašić-Kinda, S.; Coha, B.; Gverić-Krečak, V.; Vrhovac, R.; Jakšić, O.; Aurer, I.; et al. Guidelines for Diagnosis and Treatment of Chronic Lymphocytic Leukemia. Krohem B-Cll 2017. Acta Clin. Croat. 2018, 57, 190–215. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Horvat, L.; Skelin, J.; Puškarić, B.; Feliciello, I.; Heckel, D.; Madunić, J.; Kardum-Skelin, I.; Matulić, M.; Radić-Krišto, D.; Antica, M. Notch pathway connections in primary leukaemia samples of limited size. Transl. Med. Commun. 2018, 3, 8. [Google Scholar] [CrossRef]
- Horvat, L.; Antica, M.; Matulić, M. Effect of Notch and PARP Pathways’ Inhibition in Leukemic Cells. Cells 2018, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Antica, M.; Dubravčić, K.; Weber, I.; Rajić, L.; Labar, B.; Batinić, D. A search for a mutation of the Aiolos phosphorylation domain in lymphocytes from patients with leukemia. Haematologica 2007, 92, 260–261. [Google Scholar] [CrossRef]
- Matulic, M.; Skelin, J.; Radic-Kristo, D.; Kardum-Skelin, I.; Grcevic, D.; Antica, M. Notch affects the prodifferentiating effect of retinoic acid and PMA on leukemic cells. Cytometry A 2015, 87, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T.M.; Zúñiga-Pflücker, J.C. Induction of T cell development from hematopoietic progenitor cells by delta-like-1 in vitro. Immunity 2002, 17, 749–756. [Google Scholar] [CrossRef]
- Zenz, T.; Mertens, D.; Küppers, R.; Döhner, H.; Stilgenbauer, S. From pathogenesis to treatment of chronic lymphocytic leukaemia. Nat. Rev. Cancer 2010, 10, 37–50. [Google Scholar] [CrossRef]
- Georgopoulos, K. The making of a lymphocyte: The choice among disparate cell fates and the IKAROS enigma. Genes Dev. 2017, 31, 439–450. [Google Scholar] [CrossRef]
- Billot, K.; Soeur, J.; Chereau, F.; Arrouss, I.; Merle-Béral, H.; Huang, M.E.; Mazier, D.; Baud, V.; Rebollo, A. Deregulation of Aiolos expression in chronic lymphocytic leukemia is associated with epigenetic modifications. Blood 2011, 117, 1917–1927. [Google Scholar] [CrossRef] [PubMed]
- Mastio, J.; Simand, C.; Cova, G.; Kastner, P.; Chan, S.; Kirstetter, P. Ikaros cooperates with Notch activation and antagonizes TGFβ signaling to promote pDC development. PLoS Genet. 2018, 14, e1007485. [Google Scholar] [CrossRef] [PubMed]
- Talbot, H.; Saada, S.; Barthout, E.; Gallet, P.F.; Gachard, N.; Abraham, J.; Jaccard, A.; Troutaud, D.; Lalloué, F.; Naves, T.; et al. BDNF belongs to the nurse-like cell secretome and supports survival of B chronic lymphocytic leukemia cells. Sci. Rep. 2020, 10, 12572. [Google Scholar] [CrossRef] [PubMed]
- Trimarco, V.; Ave, E.; Facco, M.; Chiodin, G.; Frezzato, F.; Martini, V.; Gattazzo, C.; Lessi, F.; Giorgi, C.A.; Visentin, A.; et al. Cross-talk between chronic lymphocytic leukemia (CLL) tumor B cells and mesenchymal stromal cells (MSCs): Implications for neoplastic cell survival. Oncotarget 2015, 6, 42130–42149. [Google Scholar] [CrossRef] [PubMed]
- Rosati, E.; Baldoni, S.; De Falco, F.; Del Papa, B.; Dorillo, E.; Rompietti, C.; Albi, E.; Falzetti, F.; Di Ianni, M.; Sportoletti, P. NOTCH1 Aberrations in Chronic Lymphocytic Leukemia. Front. Oncol. 2018, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Nwabo Kamdje, A.H.; Bassi, G.; Pacelli, L.; Malpeli, G.; Amati, E.; Nichele, I.; Pizzolo, G.; Krampera, M. Role of stromal cell-mediated Notch signaling in CLL resistance to chemotherapy. Blood Cancer J. 2012, 2, e73. [Google Scholar] [CrossRef] [PubMed]
- Menendez, P.; Vargas, A.; Bueno, C.; Barrena, S.; Almeida, J.; De Santiago, M.; López, A.; Roa, S.; San Miguel, J.F.; Orfao, A. Quantitative analysis of bcl-2 expression in normal and leukemic human B-cell differentiation. Leukemia 2004, 18, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Ferracin, M.; Cimmino, A.; Di Leva, G.; Shimizu, M.; Wojcik, S.E.; Iorio, M.V.; Visone, R.; Sever, N.I.; Fabbri, M.; et al. MicroRNA signature associated with prognosis and progression in chronic lymphocytic leukemia. N. Engl. J. Med. 2005, 353, 1793–1801. [Google Scholar] [CrossRef]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Nat. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef]
- Ferrajoli, A.; Shanafelt, T.D.; Ivan, C.; Shimizu, M.; Rabe, K.G.; Nouraee, N.; Ikuo, M.; Ghosh, A.K.; Lerner, S.; Rassenti, L.Z.; et al. Prognostic value of miR-155 in individuals with monoclonal B-cell lymphocytosis and patients with B chronic lymphocytic leukemia. Blood 2013, 122, 1891–1899. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, H.; Rodriguez, S.; Cao, L.; Parish, J.; Mumaw, C.; Zollman, A.; Kamoka, M.M.; Mu, J.; Chen, D.Z.; et al. Notch-dependent repression of miR-155 in the bone marrow niche regulates hematopoiesis in an NF-κB-dependent manner. Cell Stem Cell 2014, 15, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Filip, A.A.; Grenda, A.; Popek, S.; Koczkodaj, D.; Michalak-Wojnowska, M.; Budzyński, M.; Wąsik-Szczepanek, E.; Zmorzyński, S.; Karczmarczyk, A.; Giannopoulos, K. Expression of circulating miRNAs associated with lymphocyte differentiation and activation in CLL-another piece in the puzzle. Ann. Hematol. 2017, 96, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Pekarsky, Y.; Santanam, U.; Cimmino, A.; Palamarchuk, A.; Efanov, A.; Maximov, V.; Volinia, S.; Alder, H.; Liu, C.G.; Rassenti, L.; et al. Tcl1 expression in chronic lymphocytic leukemia is regulated by miR-29 and miR-181. Cancer Res. 2006, 66, 11590–11593. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.X.; Zhu, W.; Fang, C.; Fan, L.; Zou, Z.J.; Wang, Y.H.; Liu, P.; Hong, M.; Miao, K.R.; Liu, P.; et al. miR-181a/b significantly enhances drug sensitivity in chronic lymphocytic leukemia cells via targeting multiple anti-apoptosis genes. Carcinogenesis 2012, 33, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xu, Y.; Jing, Z.; Wang, X.; Zha, X.; Zeng, C.; Chen, S.; Yang, L.; Luo, G.; Li, B.; et al. Altered expression pattern of miR-29a, miR-29b and the target genes in myeloid leukemia. Expl. Hematol. Oncol. 2014, 3, 17. [Google Scholar] [CrossRef]
- Santanam, U.; Zanesi, N.; Efanov, A.; Costinean, S.; Palamarchuk, A.; Hagan, J.P.; Volinia, S.; Alder, H.; Rassenti, L.; Kipps, T.; et al. Chronic lymphocytic leukemia modeled in mouse by targeted miR-29 expression. Proc. Nat. Acad. Sci. USA 2010, 107, 12210–12215. [Google Scholar] [CrossRef] [PubMed]
- Negrini, M.; Cutrona, G.; Bassi, C.; Fabris, S.; Zagatti, B.; Colombo, M.; Ferracin, M.; D’Abundo, L.; Saccenti, E.; Matis, S.; et al. microRNAome expression in chronic lymphocytic leukemia: Comparison with normal B-cell subsets and correlations with prognostic and clinical parameters. Clin. Cancer Res. 2014, 20, 4141–4153. [Google Scholar] [CrossRef] [PubMed]
- Blume, C.J.; Hotz-Wagenblatt, A.; Hüllein, J.; Sellner, L.; Jethwa, A.; Stolz, T.; Slabicki, M.; Lee, K.; Sharathchandra, A.; Benner, A.; et al. p53-dependent non-coding RNA networks in chronic lymphocytic leukemia. Leukemia 2015, 29, 2015–2023. [Google Scholar] [CrossRef]
- Dufour, A.; Palermo, G.; Zellmeier, E.; Mellert, G.; Duchateau-Nguyen, G.; Schneider, S.; Benthaus, T.; Kakadia, P.M.; Spiekermann, K.; Hiddemann, W.; et al. Inactivation of TP53 correlates with disease progression and low miR-34a expression in previously treated chronic lymphocytic leukemia patients. Blood 2013, 121, 3650–3657. [Google Scholar] [CrossRef]
- Novak, I.; Jaksić, O.; Kulis, T.; Batinjan, K.; Znaor, A. Incidence and mortality trends of leukemia and lymphoma in Croatia, 1988–2009. Croat. Med. J. 2012, 53, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Cerreto, M.; Foà, R.; Natoni, A. The Role of the Microenvironment and Cell Adhesion Molecules in Chronic Lymphocytic Leukemia. Cancers 2023, 15, 5160. [Google Scholar] [CrossRef] [PubMed]
- Koehrer, S.; Burger, J.A. Chronic Lymphocytic Leukemia: Disease Biology. Acta Haematol. 2024, 147, 8–21. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLL Classification | Stage | No. of Patients | Age, Years | Leukocytes (×109/L) | Lymphocytes (×109/L) | Neutrophils (×109/L) | Thrombocytes (×109/L) | Hemoglobin (g/L) |
| Binet | A | 8 | 64 (37–77) | 55 (11–107) | 47 (5–99) | 4.6 (2–7) | 203 (111–304) | 139 (127–160) |
| B | 6 | 61 (45–70) | 68 (19–250) | 62 (12–262) | 3.2 (2–5) | 214 (128–491) | 120 (106–140) | |
| C | 6 | 66 (54–75) | 189 (23–645) | 175 (20–612) | 7.0 (1–13) | 125 (43–262) | 88 (73–117) | |
| Rai | 0 | 2 | 69 (63–74) | 41 (11–70) | 30 (5–54) | 5.0 (5–5) | 251 (198–304) | 130 (127–133) |
| I | 5 | 57 (37–75) | 45 (15–107) | 40 (11–99) | 4.0 (2–5) | 204 (183–228) | 142 (130–160) | |
| II | 5 | 69 (65–77) | 48 (15–85) | 42 (12–78) | 4.0 (2–7) | 147 (119–166) | 129 (112–146) | |
| III | 4 | 64 (45–75) | 157 (46–250) | 149 (42–242) | 5.3 (2–8) | 269 (152–491) | 91 (67–111) | |
| IV | 4 | 63 (54–69) | 200 (23–645) | 184 (20–612) | 6.8 (1–13) | 80 (43–98) | 95 (73–117) | |
| TTM | 0–9 | 8 | 61 (37–75) | 41 (11–107) | 35 (5–99) | 3.9 (1–5) | 203 (98–304) | 133 (92–160) |
| 10–15 | 7 | 69 (63–77) | 64 (15–92) | 55 (12–83) | 5.3 (2–8) | 145 (85–262) | 118 (81–146) | |
| >15 | 5 | 60 (54–70) | 240 (20–645) | 227 (13–612) | 6.0 (2–13) | 203 (43–491) | 94 (67–112) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skelin, J.; Matulić, M.; Milković, L.; Heckel, D.; Skoko, J.; Škreb, K.A.; Jelić Puškarić, B.; Kardum-Skelin, I.; Čičin-Šain, L.; Radić-Krišto, D.; et al. Analysis of Primary Chronic Lymphocytic Leukemia Cells’ Signaling Pathways. Biomedicines 2024, 12, 524. https://doi.org/10.3390/biomedicines12030524
Skelin J, Matulić M, Milković L, Heckel D, Skoko J, Škreb KA, Jelić Puškarić B, Kardum-Skelin I, Čičin-Šain L, Radić-Krišto D, et al. Analysis of Primary Chronic Lymphocytic Leukemia Cells’ Signaling Pathways. Biomedicines. 2024; 12(3):524. https://doi.org/10.3390/biomedicines12030524
Chicago/Turabian StyleSkelin, Josipa, Maja Matulić, Lidija Milković, Darko Heckel, Jelena Skoko, Kristina Ana Škreb, Biljana Jelić Puškarić, Ika Kardum-Skelin, Lipa Čičin-Šain, Delfa Radić-Krišto, and et al. 2024. "Analysis of Primary Chronic Lymphocytic Leukemia Cells’ Signaling Pathways" Biomedicines 12, no. 3: 524. https://doi.org/10.3390/biomedicines12030524
APA StyleSkelin, J., Matulić, M., Milković, L., Heckel, D., Skoko, J., Škreb, K. A., Jelić Puškarić, B., Kardum-Skelin, I., Čičin-Šain, L., Radić-Krišto, D., & Antica, M. (2024). Analysis of Primary Chronic Lymphocytic Leukemia Cells’ Signaling Pathways. Biomedicines, 12(3), 524. https://doi.org/10.3390/biomedicines12030524