

L-DOS47 Elevates Pancreatic Cancer Tumor pH and Enhances Response to Immunotherapy
 , , , , , , , , , and
, , , , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Transduction and Selection of CEACAM6-Expressing Clone
2.2. Flow Cytometry
2.2.1. CEACAM6 Expression
2.2.2. Tumor Digestion and Single-Cell Preparation for Flow Cytometry Analysis
2.3. In Vitro Metabolic Profiling
2.3.1. Oxygen Consumption and Extracellular Acidification Measurements
2.3.2. Lactate Measurement
2.4. KPC961 Orthotopic Tumor Model
2.5. CEST Imaging
2.6. Treatments for Efficacy Study
2.7. Ultrasound Imaging
2.8. Statistical Analyses
3. Results
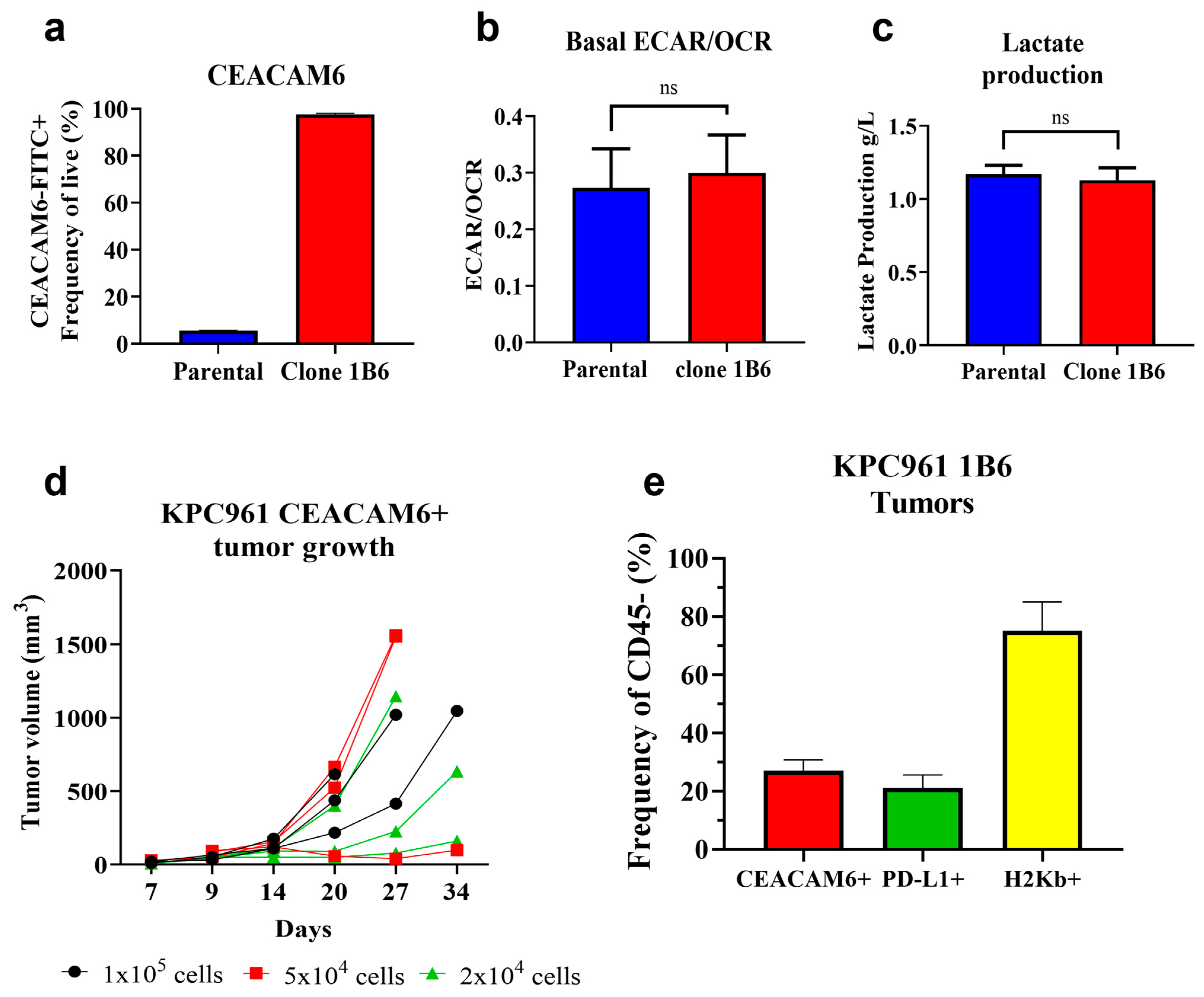
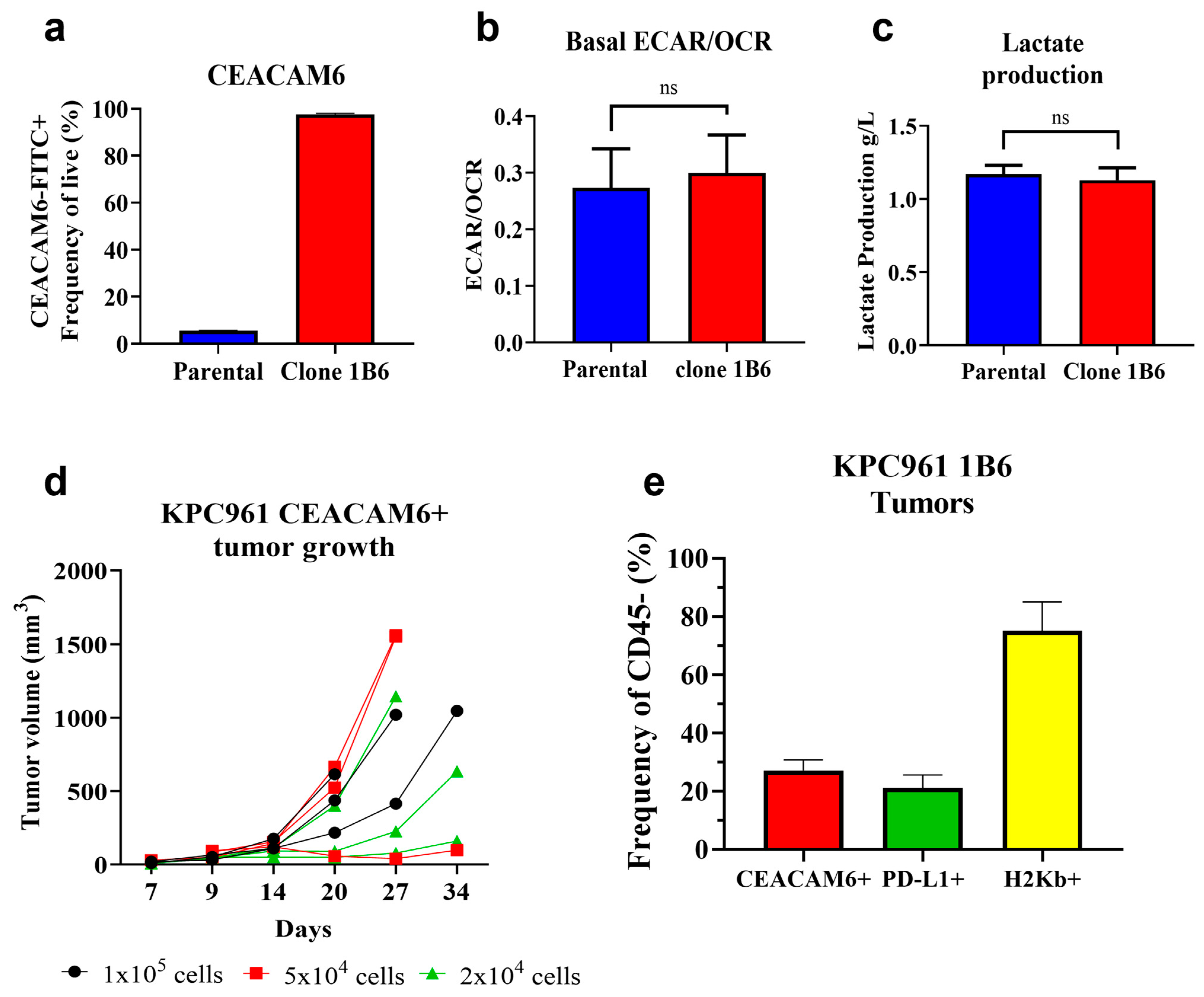
3.1. Development of an Orthotopic Pancreatic Tumor Model That Expresses Human CEACAM6
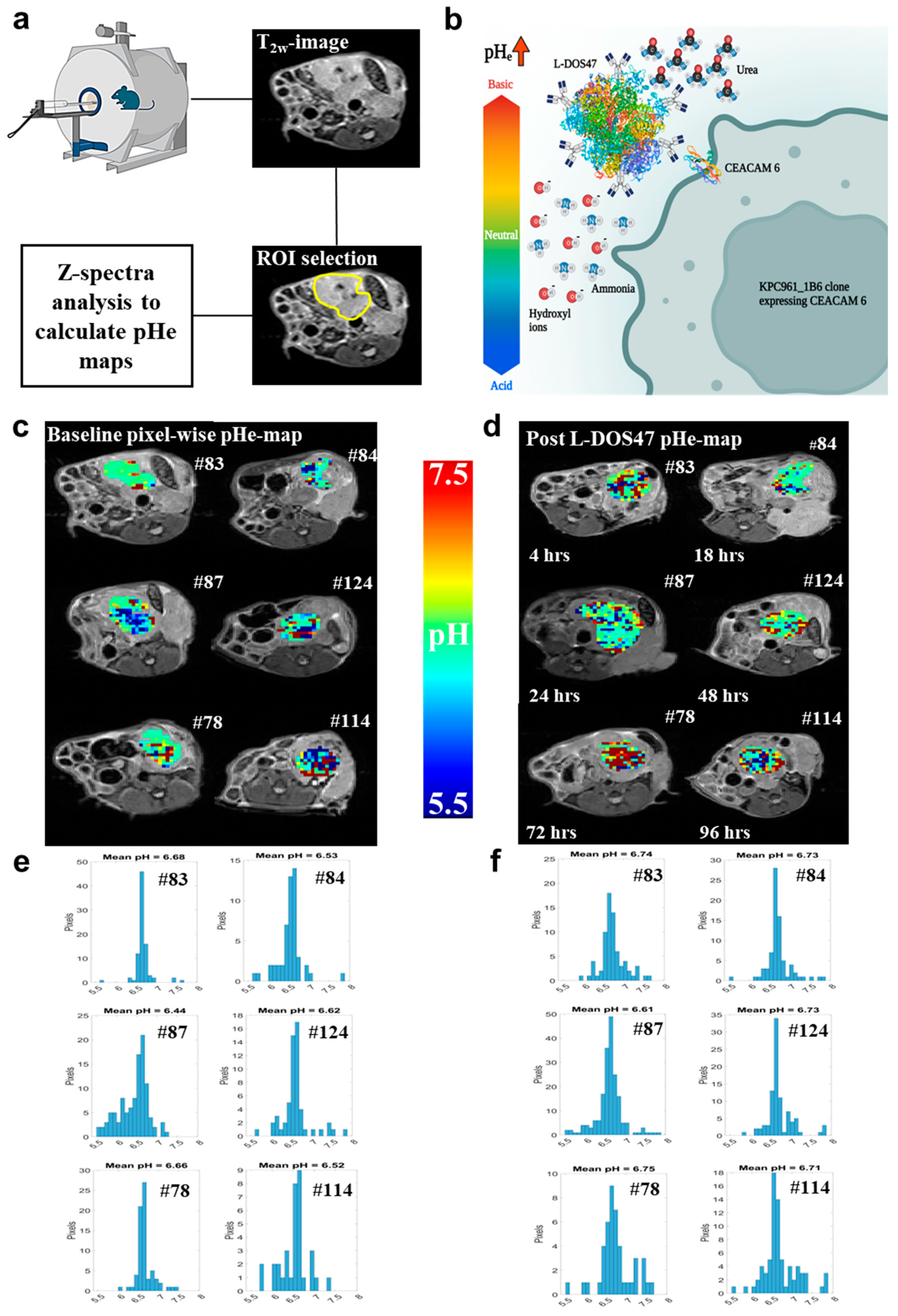
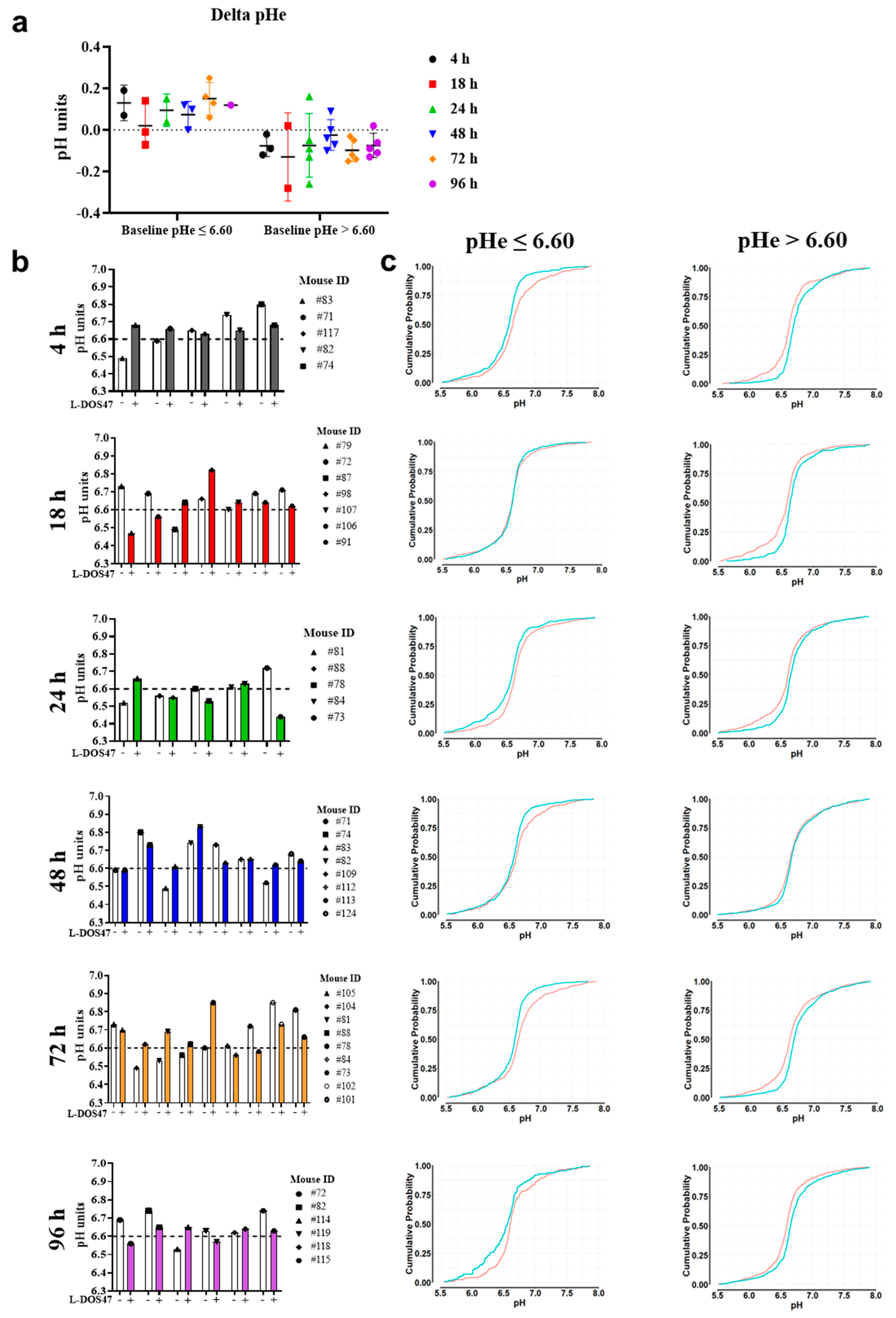
3.2. L-DOS47 Increases Tumor Extracellular pH in Acidic Tumors
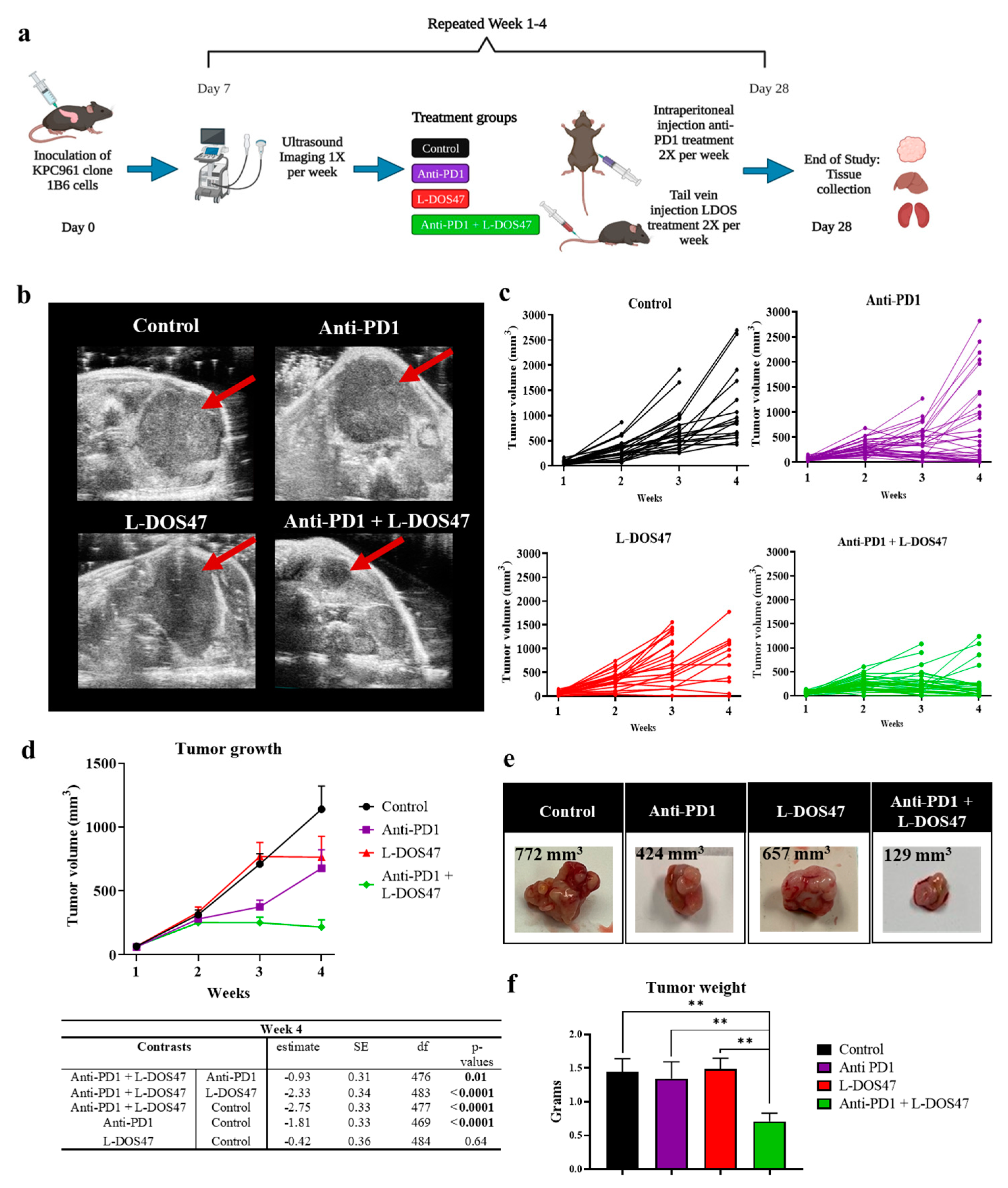
3.3. L-DOS47 Has a Synergistic Effect on Anti-PD1 Therapy in Reducing Tumor Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gatenby, R.A.; Gillies, R.J. Why do cancers have high aerobic glycolysis? Nat. Rev. Cancer 2004, 4, 891–899. [Google Scholar] [CrossRef]
- Corbet, C.; Feron, O. Tumour acidosis: From the passenger to the driver’s seat. Nat. Rev. Cancer 2017, 17, 577–593. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Estrella, V.; Beatty, M.; Abrahams, D.; El-Kenawi, A.; Russell, S.; Ibrahim-Hashim, A.; Longo, D.L.; Reshetnyak, Y.K.; Moshnikova, A.; et al. T-cells produce acidic niches in lymph nodes to suppress their own effector functions. Nat. Commun. 2020, 11, 4113. [Google Scholar] [CrossRef] [PubMed]
- Damgaci, S.; Ibrahim-Hashim, A.; Enriquez-Navas, P.M.; Pilon-Thomas, S.; Guvenis, A.; Gillies, R.J. Hypoxia and acidosis: Immune suppressors and therapeutic targets. Immunology 2018, 154, 354–362. [Google Scholar] [CrossRef]
- Bohn, T.; Rapp, S.; Luther, N.; Klein, M.; Bruehl, T.-J.; Kojima, N.; Lopez, P.A.; Hahlbrock, J.; Muth, S.; Endo, S.; et al. Tumor immunoevasion via acidosis-dependent induction of regulatory tumor-associated macrophages. Nat. Immunol. 2018, 19, 1319–1329. [Google Scholar] [CrossRef]
- Davern, M.; Donlon, N.E.; O’connell, F.; Gaughan, C.; O’donovan, C.; Habash, M.; Sheppard, A.D.; MacLean, M.; Dunne, M.R.; Moore, J.; et al. Acidosis significantly alters immune checkpoint expression profiles of T cells from oesophageal adenocarcinoma patients. Cancer Immunol. Immunother. 2023, 72, 55–71. [Google Scholar] [CrossRef]
- Pötzl, J.; Roser, D.; Bankel, L.; Hömberg, N.; Geishauser, A.; Brenner, C.D.; Weigand, M.; Röcken, M.; Mocikat, R. Reversal of tumor acidosis by systemic buffering reactivates NK cells to express IFN-γ and induces NK cell-dependent lymphoma control without other immunotherapies. Int. J. Cancer 2017, 140, 2125–2133. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.S.; Yunes, J.A.; Gillies, R.J.; Gatenby, R.A. The potential role of systemic buffers in reducing intratumoral extracellular pH and acid-mediated invasion. Cancer Res. 2009, 69, 2677–2684. [Google Scholar] [CrossRef]
- Robey, I.F.; Baggett, B.K.; Kirkpatrick, N.D.; Roe, D.J.; Dosescu, J.; Sloane, B.F.; Hashim, A.I.; Morse, D.L.; Raghunand, N.; Gatenby, R.A.; et al. Bicarbonate increases tumor pH and inhibits spontaneous metastases. Cancer Res. 2009, 69, 2260–2268. [Google Scholar] [CrossRef]
- Persi, E.; Duran-Frigola, M.; Damaghi, M.; Roush, W.R.; Aloy, P.; Cleveland, J.L.; Gillies, R.J.; Ruppin, E. Systems analysis of intracellular pH vulnerabilities for cancer therapy. Nat. Commun. 2018, 9, 2997. [Google Scholar] [CrossRef]
- Bellone, M.; Calcinotto, A.; Filipazzi, P.; De Milito, A.; Fais, S.; Rivoltini, L. The acidity of the tumor microenvironment is a mechanism of immune escape that can be overcome by proton pump inhibitors. OncoImmunology 2013, 2, e22058. [Google Scholar] [CrossRef] [PubMed]
- Calcinotto, A.; Filipazzi, P.; Grioni, M.; Iero, M.; De Milito, A.; Ricupito, A.; Cova, A.; Canese, R.; Jachetti, E.; Rossetti, M.; et al. Modulation of microenvironment acidity reverses anergy in human and murine tumor-infiltrating T lymphocytes. Cancer Res. 2012, 72, 2746–2756. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Perut, F.; Fagioli, F.; Del Prever, A.B.; Meazza, C.; Parafioriti, A.; Picci, P.; Gambarotti, M.; Avnet, S.; Baldini, N.; et al. Proton pump inhibitor chemosensitization in human osteosarcoma: From the bench to the patients’ bed. J. Transl. Med. 2013, 11, 268. [Google Scholar] [CrossRef]
- Wang, B.-Y.; Zhang, J.; Wang, J.-L.; Sun, S.; Wang, Z.-H.; Wang, L.-P.; Zhang, Q.-L.; Lv, F.-F.; Cao, E.-Y.; Shao, Z.-M.; et al. Intermittent high dose proton pump inhibitor enhances the antitumor effects of chemotherapy in metastatic breast cancer. J. Exp. Clin. Cancer Res. 2015, 34, 85. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Thomas, S.; Kodumudi, K.N.; El-Kenawi, A.E.; Russell, S.; Weber, A.M.; Luddy, K.; Damaghi, M.; Wojtkowiak, J.W.; Mule, J.J.; Ibrahim-Hashim, A.; et al. Neutralization of Tumor Acidity Improves Antitumor Responses to Immunotherapy. Cancer Res. 2016, 76, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Gillies, R.J.; Ibrahim-Hashim, A.; Ordway, B.; Gatenby, R.A. Back to basic: Trials and tribulations of alkalizing agents in cancer. Front. Oncol. 2022, 12, 981718. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, N.; Arabzadeh, A. Carcinoembryonic antigen-related cell adhesion molecules (CEACAMs) in cancer progression and metastasis. Cancer Metastasis Rev. 2013, 32, 643–671. [Google Scholar] [CrossRef]
- Burgos, M.; Cavero-Redondo, I.; Álvarez-Bueno, C.; Galán-Moya, E.M.; Pandiella, A.; Amir, E.; Ocaña, A. Prognostic value of the immune target CEACAM6 in cancer: A meta-analysis. Ther. Adv. Med. Oncol. 2022, 14, 17588359211072621. [Google Scholar] [CrossRef]
- Tian, B.; Wong, W.Y.; Hegmann, E.; Gaspar, K.; Kumar, P.; Chao, H. Production and characterization of a camelid single domain antibody-urease enzyme conjugate for the treatment of cancer. Bioconjug. Chem. 2015, 26, 1144–1155. [Google Scholar] [CrossRef]
- Piha-Paul, S.; Simon, G.; Belani, C.P.; Chao, H.; Gaspar, K.; Lee, B.; Dowlati, A. A Phase 1, Open-Label, Dose-Escalation Study of L-DOS47 in Combination with Pemetrexed Plus Carboplatin in Patients with Stage IV Recurrent or Metastatic Nonsquamous NSCLC. JTO Clin. Res. Rep. 2022, 3, 100408. [Google Scholar] [CrossRef]
- An, F.; Zheng, C.; Zhang, G.; Zhou, L.; Wu, Y.; Hou, Z.; Zhou, Z.; Chen, K.; Zhan, Q. Carcinoembryonic Antigen Related Cell Adhesion Molecule 6 Promotes Carcinogenesis of Gastric Cancer and Anti-CEACAM6 Fluorescent Probe Can Diagnose the Precancerous Lesions. Front. Oncol. 2021, 11, 643669. [Google Scholar] [CrossRef]
- Kurlinkus, B.; Ger, M.; Kaupinis, A.; Jasiunas, E.; Valius, M.; Sileikis, A. CEACAM6′s Role as a Chemoresistance and Prognostic Biomarker for Pancreatic Cancer: A Comparison of CEACAM6′s Diagnostic and Prognostic Capabilities with Those of CA19-9 and CEA. Life 2021, 11, 542. [Google Scholar] [CrossRef]
- Balachandran, V.P.; Beatty, G.L.; Dougan, S.K. Broadening the Impact of Immunotherapy to Pancreatic Cancer: Challenges and Opportunities. Gastroenterology 2019, 156, 2056–2072. [Google Scholar] [CrossRef]
- Longo, D.L.; Dastrù, W.; Digilio, G.; Keupp, J.; Langereis, S.; Lanzardo, S.; Prestigio, S.; Steinbach, O.; Terreno, E.; Uggeri, F.; et al. Iopamidol as a responsive MRI-chemical exchange saturation transfer contrast agent for pH mapping of kidneys: In vivo studies in mice at 7 T. Magn. Reson. Med. 2011, 65, 202–211. [Google Scholar] [CrossRef]
- Anemone, A.; Consolino, L.; Conti, L.; Irrera, P.; Hsu, M.Y.; Villano, D.; Dastrù, W.; Porporato, P.E.; Cavallo, F.; Longo, D.L. Tumour acidosis evaluated in vivo by MRI-CEST pH imaging reveals breast cancer metastatic potential. Br. J. Cancer 2021, 124, 207–216. [Google Scholar] [CrossRef]
- Jardim-Perassi, B.V.; Irrera, P.; Lau, J.Y.C.; Budzevich, M.; Whelan, C.J.; Abrahams, D.; Ruiz, E.; Ibrahim-Hashim, A.; Erturk, S.D.; Longo, D.L.; et al. Intraperitoneal Delivery of Iopamidol to Assess Extracellular pH of Orthotopic Pancreatic Tumor Model by CEST-MRI. Contrast Media Mol. Imaging 2023, 2023, 1944970. [Google Scholar] [CrossRef] [PubMed]
- Irrera, P.; Roberto, M.; Consolino, L.; Anemone, A.; Villano, D.; Navarro-Tableros, V.; Carella, A.; Dastrù, W.; Aime, S.; Longo, D.L. Effect of Esomeprazole Treatment on Extracellular Tumor pH in a Preclinical Model of Prostate Cancer by MRI-CEST Tumor pH Imaging. Metabolites 2022, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Anemone, A.; Consolino, L.; Conti, L.; Reineri, F.; Cavallo, F.; Aime, S.; Longo, D.L. In vivo evaluation of tumour acidosis for assessing the early metabolic response and onset of resistance to dichloroacetate by using magnetic resonance pH imaging. Int. J. Oncol. 2017, 51, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.M.; Randtke, E.A.; Yoshimaru, E.S.; Howison, C.M.; Chalasani, P.; Klein, R.R.; Chambers, S.K.; Kuo, P.H.; Pagel, M.D. Clinical Translation of Tumor Acidosis Measurements with AcidoCEST MRI. Mol. Imaging Biol. 2017, 19, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Müller-Lutz, A.; Khalil, N.; Schmitt, B.; Jellus, V.; Pentang, G.; Oeltzschner, G.; Antoch, G.; Lanzman, R.S.; Wittsack, H.-J. Pilot study of Iopamidol-based quantitative pH imaging on a clinical 3T MR scanner. Magn. Reson. Mater. Phys. Biol. Med. 2014, 27, 477–485. [Google Scholar] [CrossRef]
- Tang, Y.; Xiao, G.; Shen, Z.; Zhuang, C.; Xie, Y.; Zhang, X.; Yang, Z.; Guan, J.; Shen, Y.; Chen, Y.; et al. Noninvasive Detection of Extracellular pH in Human Benign and Malignant Liver Tumors Using CEST MRI. Front. Oncol. 2020, 10, 578985. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.P.; Rachagani, S.; Souchek, J.J.; Mallya, K.; Johansson, S.L.; Batra, S.K. Novel Pancreatic Cancer Cell Lines Derived from Genetically Engineered Mouse Models of Spontaneous Pancreatic Adenocarcinoma: Applications in Diagnosis and Therapy. PLoS ONE 2013, 8, e80580. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Almhanna, K. Pancreatic cancer and immune checkpoint inhibitors—Still a long way to go. Transl. Gastroenterol. Hepatol. 2021, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Marabelle, A.; Le, D.T.; Ascierto, P.A.; Di Giacomo, A.M.; De Jesus-Acosta, A.; Delord, J.-P.; Geva, R.; Gottfried, M.; Penel, N.; Hansen, A.R.; et al. Efficacy of Pembrolizumab in Patients with Noncolorectal High Microsatellite Instability/Mismatch Repair–Deficient Cancer: Results from the Phase II KEYNOTE-158 Study. J. Clin. Oncol. 2020, 38, 1–10. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, E.M.; Oh, D.-Y.; Dhani, N.; Renouf, D.J.; Lee, M.A.; Sun, W.; Fisher, G.; Hezel, A.; Chang, S.-C.; Vlahovic, G.; et al. Durvalumab with or Without Tremelimumab for Patients with Metastatic Pancreatic Ductal Adenocarcinoma: A Phase 2 Randomized Clinical Trial. JAMA Oncol. 2019, 5, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, A.; Bogdanov, A.; Chubenko, V.; Volkov, N.; Moiseenko, F.; Moiseyenko, V. Tumor acidity: From hallmark of cancer to target of treatment. Front. Oncol. 2022, 12, 979154. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, S.N.; Poellmann, C.C.; Wisniewski, N.A.; Gaskin, A.A.; Shoemaker, J.S.; Poulson, J.M.; Dewhirst, M.W.; Klitzman, B. Urea as a recovery marker for quantitative assessment of tumor interstitial solutes with microdialysis. Cancer Res. 2001, 61, 7964–7970. [Google Scholar]
- Kabacaoglu, D.; Ciecielski, K.J.; Ruess, D.A.; Algül, H. Immune Checkpoint Inhibition for Pancreatic Ductal Adenocarcinoma: Current Limitations and Future Options. Front. Immunol. 2018, 9, 1878. [Google Scholar] [CrossRef]
- Feng, M.; Xiong, G.; Cao, Z.; Yang, G.; Zheng, S.; Song, X.; You, L.; Zheng, L.; Zhang, T.; Zhao, Y. PD-1/PD-L1 and immunotherapy for pancreatic cancer. Cancer Lett. 2017, 407, 57–65. [Google Scholar] [CrossRef]
- Mucileanu, A.; Chira, R.; Mircea, P.A. PD-1/PD-L1 expression in pancreatic cancer and its implication in novel therapies. Med. Pharm. Rep. 2021, 94, 402–410. [Google Scholar] [CrossRef]
- Pillai, S.R.; Damaghi, M.; Marunaka, Y.; Spugnini, E.P.; Fais, S.; Gillies, R.J. Causes, consequences, and therapy of tumors acidosis. Cancer Metastasis Rev. 2019, 38, 205–222. [Google Scholar] [CrossRef]
- Ibrahim-Hashim, A.; Cornnell, H.H.; Abrahams, D.; Lloyd, M.; Bui, M.; Gillies, R.J.; Gatenby, R.A. Systemic buffers inhibit carcinogenesis in TRAMP mice. J. Urol. 2012, 188, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Estrella, V.; Chen, T.; Lloyd, M.; Wojtkowiak, J.; Cornnell, H.H.; Ibrahim-Hashim, A.; Bailey, K.; Balagurunathan, Y.; Rothberg, J.M.; Sloane, B.F.; et al. Acidity generated by the tumor microenvironment drives local invasion. Cancer Res. 2013, 73, 1524–1535. [Google Scholar] [CrossRef] [PubMed]
- Pilot, C.; Mahipal, A.; Gillies, R.J. Buffer Therapy → Buffer Diet. J. Nutr. Food Sci. 2018, 8, 1–5. [Google Scholar] [CrossRef]
- Hamaguchi, R.; Narui, R.; Wada, H. Effects of Alkalization Therapy on Chemotherapy Outcomes in Metastatic or Recurrent Pancreatic Cancer. Anticancer Res. 2020, 40, 873–880. [Google Scholar] [CrossRef]
- Hamaguchi, R.; Narui, R.; Morikawa, H.; Wada, H. Improved Chemotherapy Outcomes of Patients with Small-cell Lung Cancer Treated with Combined Alkalization Therapy and Intravenous Vitamin C. Cancer Diagn. Progn. 2021, 1, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, R.; Isowa, M.; Narui, R.; Morikawa, H.; Wada, H. Clinical review of alkalization therapy in cancer treatment. Front. Oncol. 2022, 12, 1003588. [Google Scholar] [CrossRef]
- Ramlau, R.; Kowalski, D.; Szczylik, C.; Szczęsna, A.; Wiatr, E.; Demas, S.; Chao, H.; Roszkowski-Sliz, K. P2.06-006 Phase I/II Dose Escalation Study of L-DOS47 as a Monotherapy in Non-Squamous Non-Small Cell Lung Cancer Patients: Topic: Phase I/II Trials. J. Thorac. Oncol. 2017, 12, S1071–S1072. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day 13 | |||||
| Contrasts | Estimate | SE | df | p-Values | |
| Anti-PD1 + L-DOS47 | Anti-PD1 | −0.26 | 0.22 | 353 | 0.65 |
| Anti-PD1 + L-DOS47 | L-DOS47 | −0.66 | 0.25 | 405 | 0.04 |
| Anti-PD1 + L-DOS47 | Control | −0.74 | 0.24 | 363 | 0.01 |
| Anti-PD1 | Control | −0.48 | 0.25 | 334 | 0.21 |
| L-DOS47 | Control | −0.08 | 0.26 | 432 | 0.98 |
| Day 21 | |||||
| Contrasts | estimate | SE | df | p-values | |
| Anti-PD1 + L-DOS47 | Anti-PD1 | −0.60 | 0.22 | 340 | 0.03 |
| Anti-PD1 + L-DOS47 | L-DOS47 | −1.49 | 0.24 | 389 | <0.0001 |
| Anti-PD1 + L-DOS47 | Control | −1.75 | 0.24 | 347 | <0.0001 |
| Anti-PD1 | Control | −1.15 | 0.24 | 316 | <0.0001 |
| L-DOS47 | Control | −0.25 | 0.25 | 418 | 0.75 |
| Day 27 | |||||
| Contrasts | estimate | SE | df | p-values | |
| Anti-PD1 + L-DOS47 | Anti-PD1 | −0.93 | 0.31 | 476 | 0.01 |
| Anti-PD1 + L-DOS47 | L-DOS47 | −2.33 | 0.34 | 483 | <0.0001 |
| Anti-PD1 + L-DOS47 | Control | −2.75 | 0.33 | 477 | <0.0001 |
| Anti-PD1 | Control | −1.81 | 0.33 | 469 | <0.0001 |
| L-DOS47 | Control | −0.42 | 0.36 | 484 | 0.64 |
| Contrasts | Estimate | SE | df | p-Values | |
|---|---|---|---|---|---|
| Control | L-DOS47 | −0.04 | 0.28 | −0.15 | 0.99 |
| Control | Anti-PD1 | 0.10 | 0.27 | 0.37 | 0.98 |
| Control | Anti-PD1 + L-DOS47 | 0.73 | 0.24 | 3.05 | 0.01 |
| L-DOS47 | Anti-PD1 + L-DOS47 | 0.77 | 0.24 | 3.17 | 0.01 |
| Anti-PD1 | Anti-PD1 + L-DOS47 | 0.63 | 0.23 | 2.73 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jardim-Perassi, B.V.; Irrera, P.; Oluwatola, O.E.; Abrahams, D.; Estrella, V.C.; Ordway, B.; Byrne, S.R.; Ojeda, A.A.; Whelan, C.J.; Kim, J.; et al. L-DOS47 Elevates Pancreatic Cancer Tumor pH and Enhances Response to Immunotherapy. Biomedicines 2024, 12, 461. https://doi.org/10.3390/biomedicines12020461
Jardim-Perassi BV, Irrera P, Oluwatola OE, Abrahams D, Estrella VC, Ordway B, Byrne SR, Ojeda AA, Whelan CJ, Kim J, et al. L-DOS47 Elevates Pancreatic Cancer Tumor pH and Enhances Response to Immunotherapy. Biomedicines. 2024; 12(2):461. https://doi.org/10.3390/biomedicines12020461
Chicago/Turabian StyleJardim-Perassi, Bruna Victorasso, Pietro Irrera, Oluwaseyi E. Oluwatola, Dominique Abrahams, Veronica C. Estrella, Bryce Ordway, Samantha R. Byrne, Andrew A. Ojeda, Christopher J. Whelan, Jongphil Kim, and et al. 2024. "L-DOS47 Elevates Pancreatic Cancer Tumor pH and Enhances Response to Immunotherapy" Biomedicines 12, no. 2: 461. https://doi.org/10.3390/biomedicines12020461
APA StyleJardim-Perassi, B. V., Irrera, P., Oluwatola, O. E., Abrahams, D., Estrella, V. C., Ordway, B., Byrne, S. R., Ojeda, A. A., Whelan, C. J., Kim, J., Beatty, M. S., Damgaci-Erturk, S., Longo, D. L., Gaspar, K. J., Siegers, G. M., Centeno, B. A., Lau, J. Y. C., Pilon-Thomas, S. A., Ibrahim-Hashim, A., & Gillies, R. J. (2024). L-DOS47 Elevates Pancreatic Cancer Tumor pH and Enhances Response to Immunotherapy. Biomedicines, 12(2), 461. https://doi.org/10.3390/biomedicines12020461