

Cellular and Molecular Processes in Wound Healing



Abstract
:1. Introduction
2. Issues Related to the Study and Management of Wound Healing
3. Major Events in Wound Healing
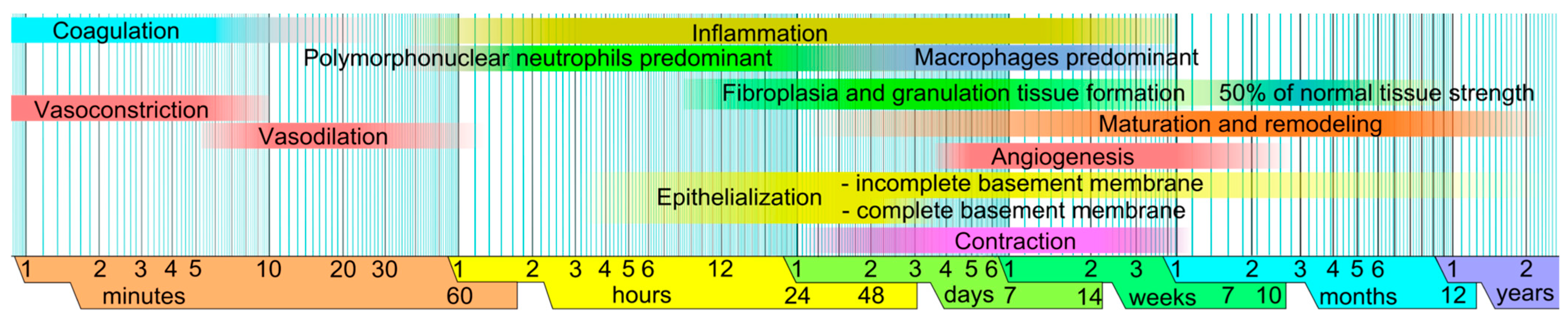
4. Recent Advances in Understanding the Key Cells Involved in Wound Healing
4.1. Phase: Hemostasis
Platelets
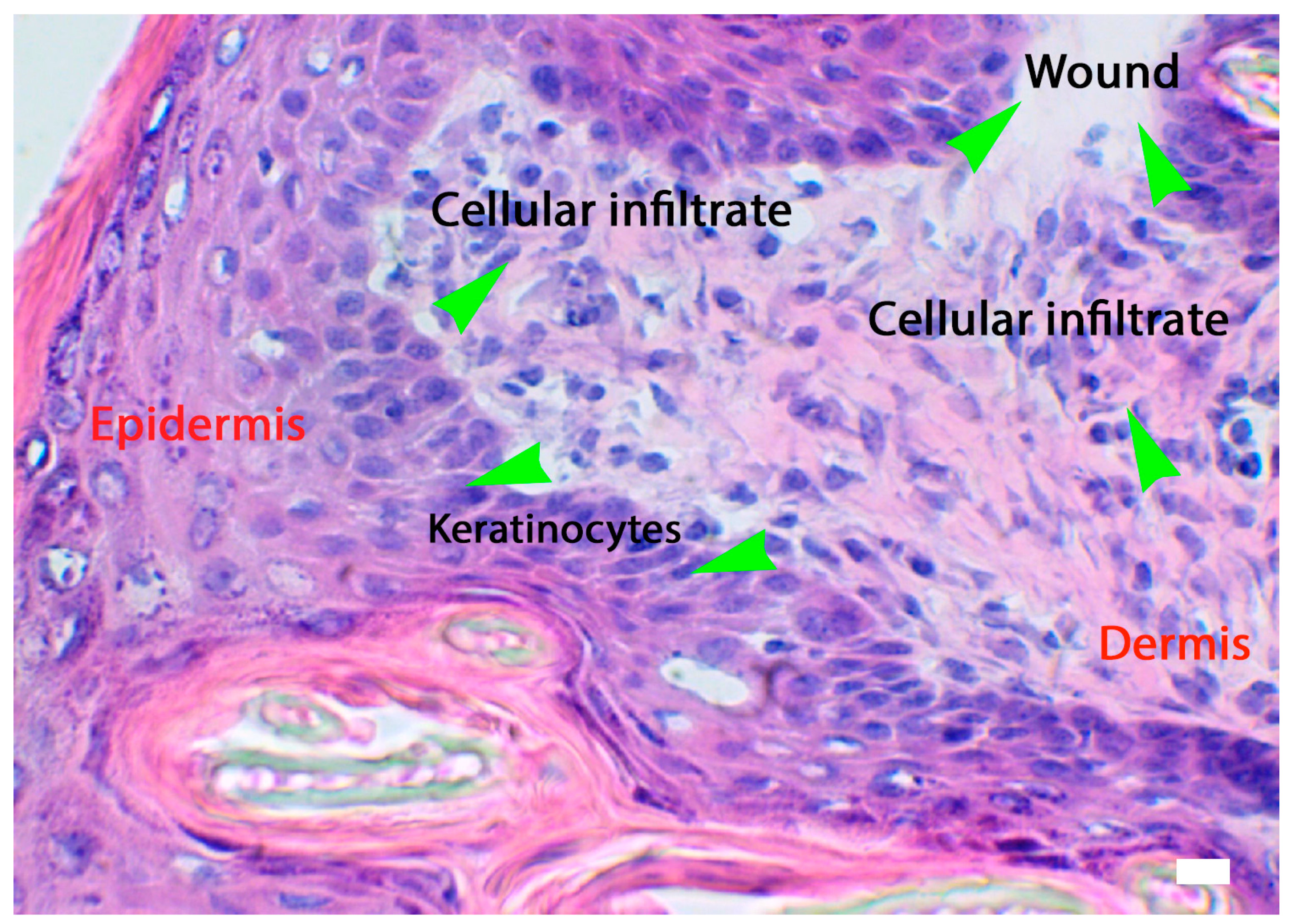
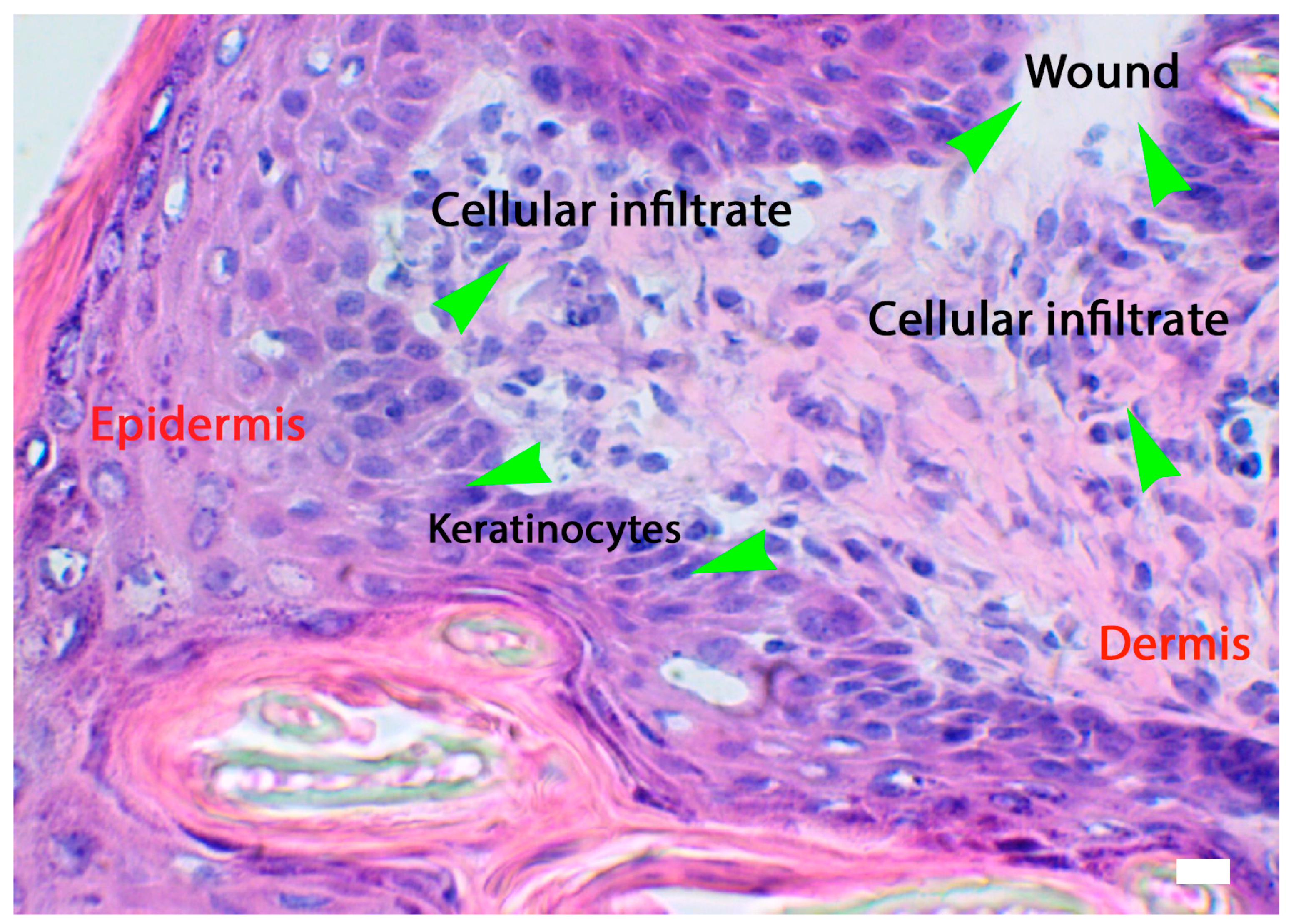
4.2. Phase: Inflammatory
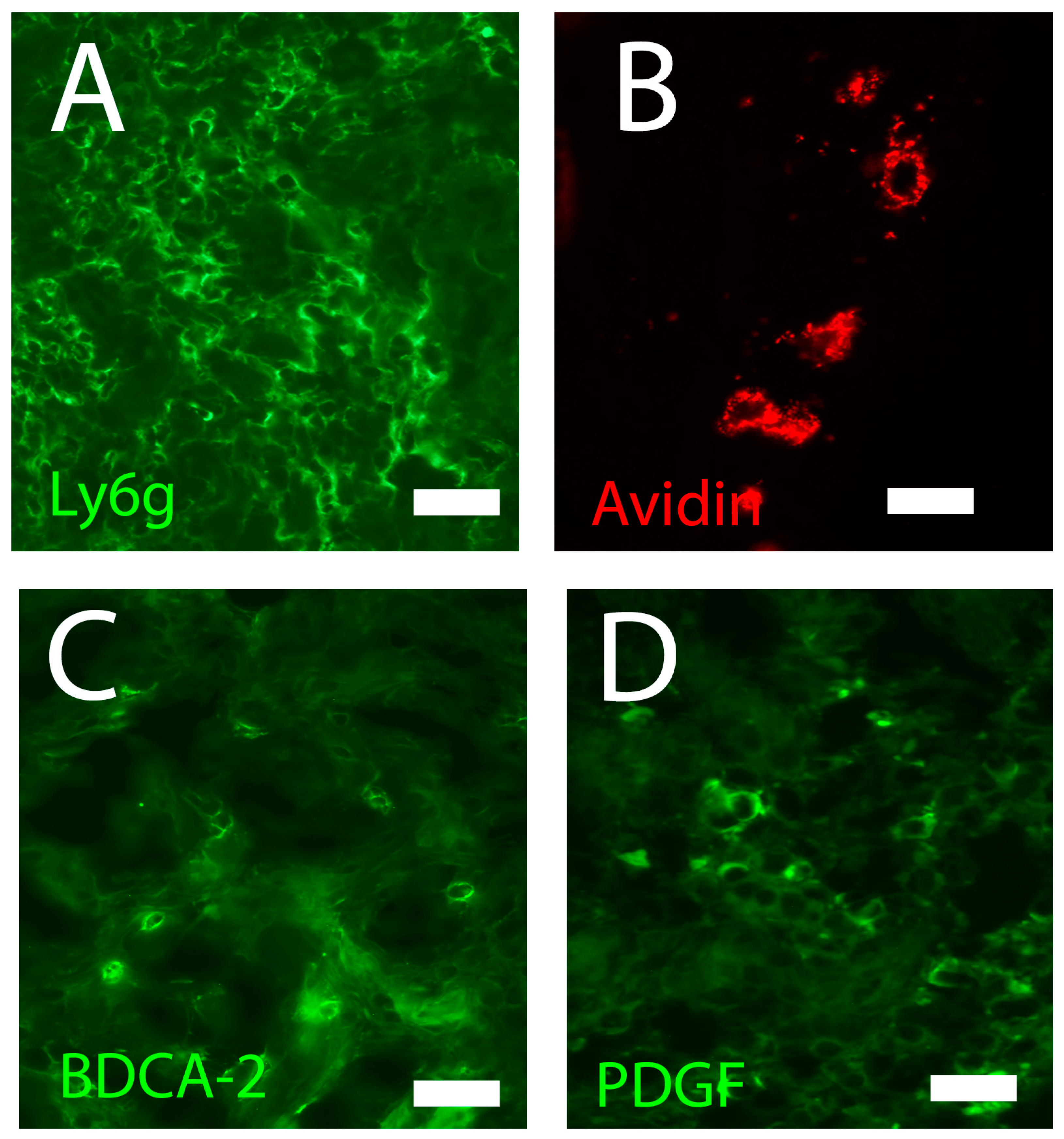
4.2.1. Mast Cells
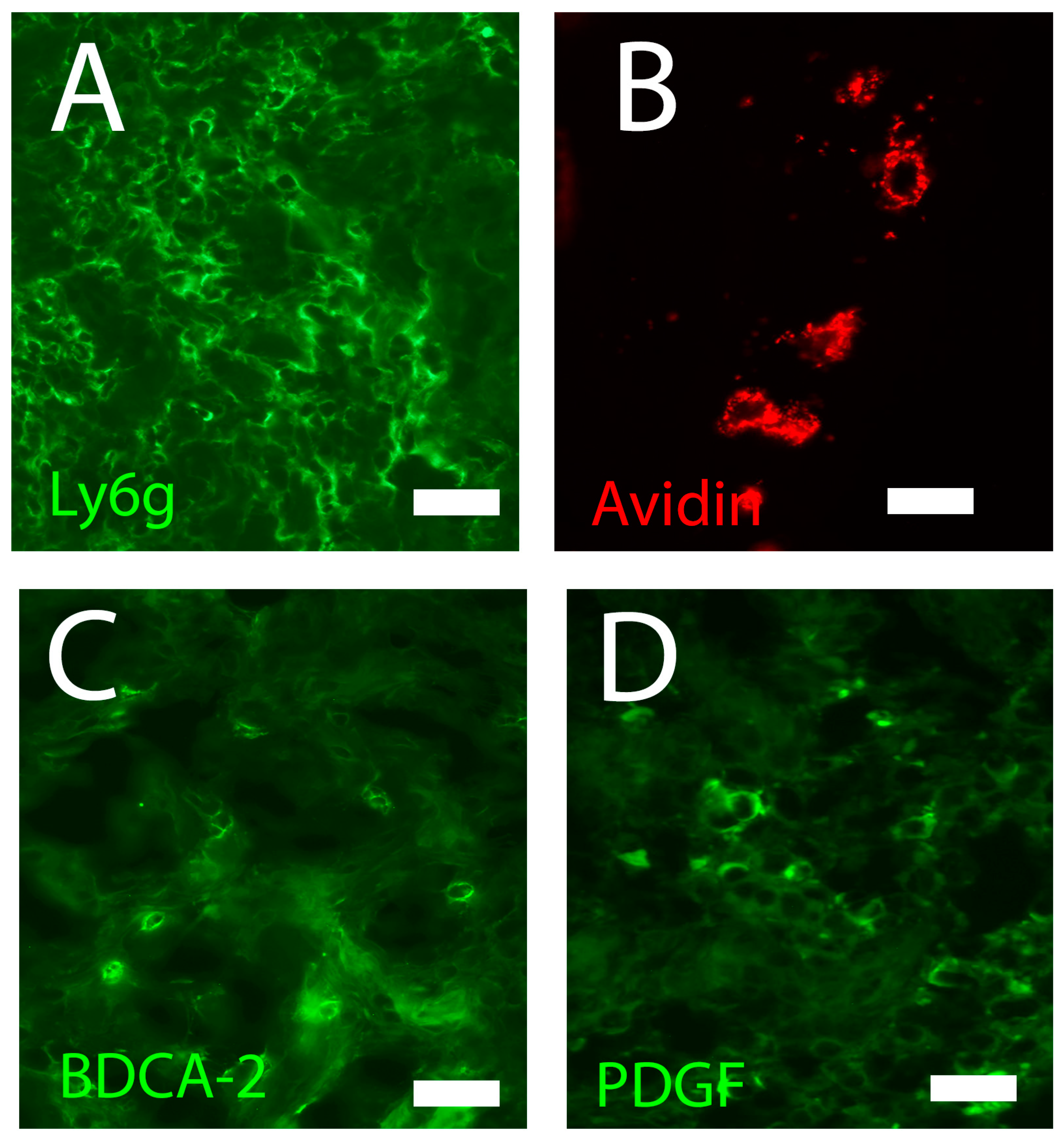
4.2.2. Neutrophils
4.2.3. Macrophages
4.2.4. Dendritic Cells
4.2.5. Plasmacytoid Dendritic Cells
4.2.6. Lymphocytes
4.3. Phase: Proliferation
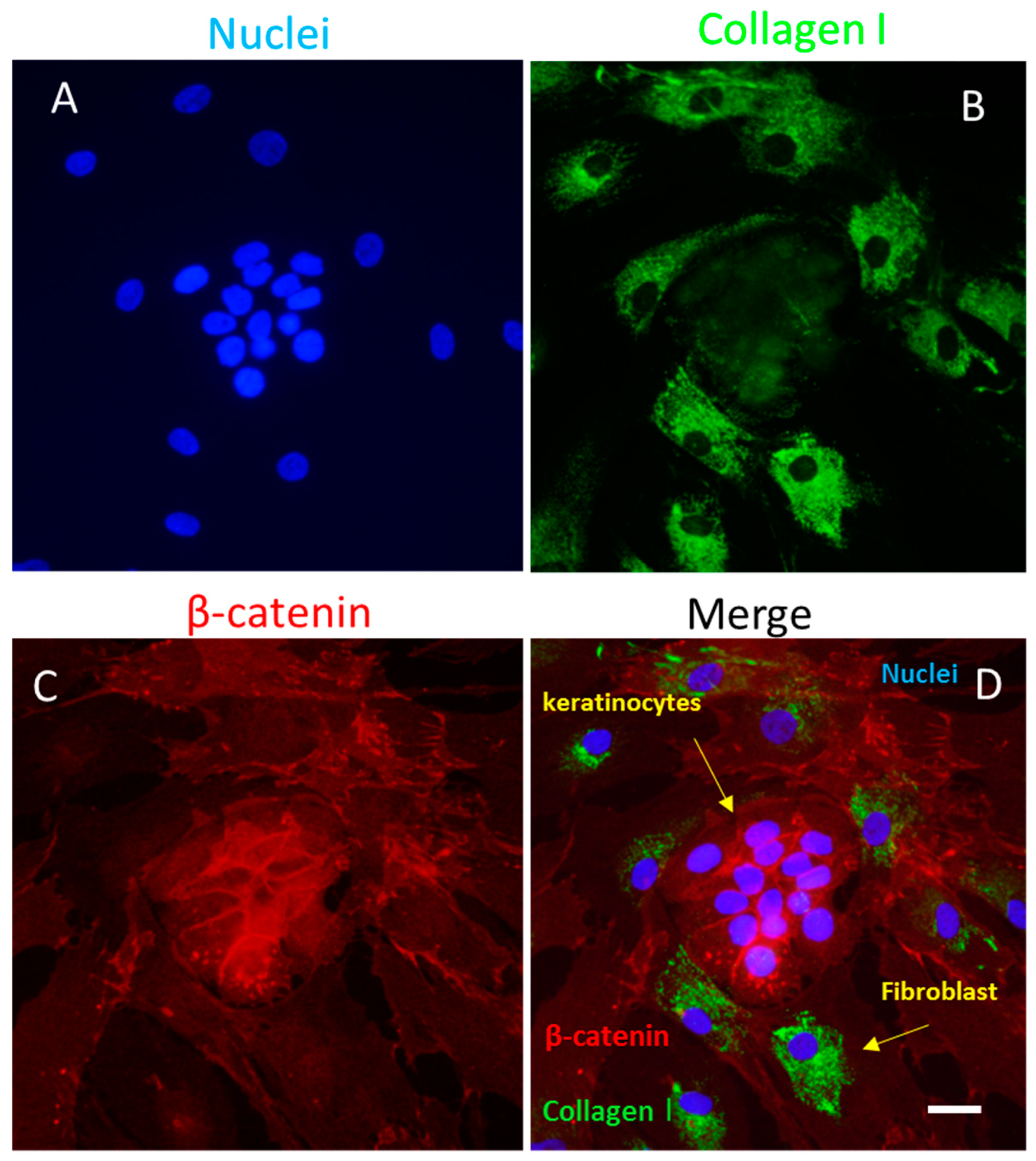
4.3.1. Keratinocytes
4.3.2. Endothelial Cells
4.3.3. Pericytes
4.3.4. Fibroblasts
4.4. Phases: Maturation and Remodeling
4.4.1. Myofibroblasts
4.4.2. The Central Role of Mast Cells in Wound Healing: A Hypothesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wound Healing | ||||
|---|---|---|---|---|
| Phases | Time | Cells | Main Bioactive Factors Secreted by Cells Involved in Wound Healing | Functions |
| Hemostasis | A few minutes | Platelets | CYTOKINES: TNFalpha GROWTH FACTORS: PDGF, TGFbeta, TGFalpha, FGF, IGF1, VEGF CHEMOKINES: CXCL8, CXCL1, CXCL2 | Initiation of inflammatory responses, angiogenesis |
| Inflammatory | 3–12 min to 3 days | Mast Cells | BIOGENIC AMMINES: Histamine CYTOKINES: TNFalpha, IL4, IL6, IL8 GROWTH FACTORS: VEGF, FGF | Vasodilation, inflammatory response, production of ECM |
| Inflammatory | 3–12 min to 3 days | Neutrophils | CYTOKINES: IL1beta, IL6, IL8, TNFalpha CHEMOKINES: CXCL1, CXCL2, CXCL8 GROWTH FACTORS: IGF, VEGF | Inflammatory response, keratinocyte proliferation, fibroblast proliferation, angiogenesis, collagen synthesis, endothelial cell activation |
| Inflammatory | 3–12 min to 3 days | Macrophages | CYTOKINES: IFNgamma, IL1beta, IL6, IL8, IL10, TNFalpha CHEMOKINES: RANTES GROWTH FACTORS: EGF, FGF, IGF, PGDF, TGFbeta, VEGF | Inflammatory response fibroblast proliferation, fibroblast chemotaxis, angiogenesis, ECM deposition |
| Inflammatory | 3–12 min to 3 days | Dendritic cells, plasmacytoid dendritic cells | GROWTH FACTORS: TGFbeta CYTOKINES: IFN gamma | Inflammatory response |
| Inflammatory | 3–12 min to 3 days | Lymphocytes | CYTOKINES: IFNgamma, IL2, IL4, IL10 CHEMOKINES: MCP, RANTES, MIP, Lymphotactin | Inflammatory response, decrease in collagen synthesis, synthesis of MMPs |
| Proliferation | 3 days to 12 days | Keratinocytes | CYTOKINES: IL1, IL6, IL8, IL10, IL18, IL20, TNFalpha GROWTH FACTORS: TGFbeta, VEGF, EGF, PGDF, CHEMOKINES: RANTES, MCP or MIP-1 | Proliferation of keratinocytes, angiogenesis, proliferation of keratinocytes, inflammatory response |
| Proliferation | 3 days to 12 days | Endothelial cells | GROWTH FACTORS: FGF, IGF, TGFbeta, PGDF, VEGF | Proliferation of fibroblasts and keratinocytes, differentiation of keratinocytes, angiogenesis |
| Maturation or remodeling | 12 days to 6 months | Fibroblasts | CHEMOKINES: CXCL1, CX3CL1, CCL2 CYTOKINES: IL6, IL8, IL12 GROWTH FACTORS: FGF, IGF, KGF, VEGF | Chemotaxis of inflammatory cells, proliferation of fibroblasts, fibroblast differentiation |
5. Molecular Events in Wound Healing
5.1. Growth Factors, Cytokines, and Other Substances
5.2. Genetic Activation in Wound Healing
6. Malfunctioning of Processes Related to Wound Healing
6.1. Chronic Wounds
6.2. Wound Healing in Conditions of Hyperglycaemia: Chronic Diabetic Wounds
7. Fibrosis: Hypertrophic Scarring and Keloids Associated with Wound-Healing Phases
7.1. Hypertrophic Scarring
7.2. Keloids
8. Impairment of Wound Healing and Recent Therapeutic Strategies
9. Future Perspective and Current Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Denomination | Acronym |
| Activator Protein 1 | AP1 |
| Angiopoietin 1 | ANG1 |
| c-Jun N-Terminal Kinase | JNK |
| Ca2+/Calmodulin-dependent Protein Kinase | CaMK |
| Calcitonin-Gene-Related Peptide | CGRP |
| Caseinphosphopeptides | CPP |
| Checkpoint Kinase 1 | CHEK1 |
| Chemokine (C-C motif) Ligand | CCL |
| Chemokine (C-C motif) Receptor | CXCR |
| Chemokine (C-X-C motif) Ligand | CXCL |
| Chronic Wound | CW |
| Collagen Alpha Chain | COL4A |
| Connective Tissue Chemokine Activating Peptide | CTAP |
| Connective Tissue Growth Factor | CTGF |
| Cyclin B1 | CCNB1 |
| Cyclin-dependent Kinase 1 | CDK1 |
| Damage-associated Molecular Patterns | DAMPS |
| Dendritic Cells | DCs |
| Diabetic Foot Ulcers | DFUs |
| Differentially Expressed Genes | DEGs |
| Dopachrome Tautomerase | DCT |
| Endothelial Progenitor Cells | EPC |
| Epidermal Growth Factor | EGF |
| Epidermal Stem Cells | ESCs |
| ETS-related Gene | ERG |
| Epithelial–Mesenchymal Transition | EMT |
| Exendin-4 | Exe4 |
| Extracellular Matrix | ECM |
| Fibroblast Growth Factor | FGF |
| Glucagon-like Peptide-1 | GLP-1 |
| Growth-related Oncogene | GRO |
| Hepatocyte Growth Factor | HGF |
| Hydrogen Peroxide | H2O2 |
| IL | Interleukin |
| Insulin-like Growth Factor | IGF |
| Interferon | IFN |
| Interferon-inducible T-cell alpha chemoattractant | ITAC |
| Interferon-g-induced Monokine | MIG |
| Interferon-g-inducible Protein 10 | IP10 |
| Keratinocyte Growth Factor | KGF |
| Light-emitting Diodes | LED |
| Low-level Light Therapy | LLLT |
| Macrophage Inflammatory Protein | MIP |
| Macrophage-derived Chemokine | MDC |
| Mast Cells | MCs |
| Matrix Metalloproteinases | MMPs |
| Mesenchymal Stem Cells | MSCs |
| Monocyte Chemoattractant Proteins | MCP |
| Metal–Organic Frameworks | MOFs |
| Mitogen-activated Protein Kinase | MAPK |
| Monokine-induced Gamma Interferon | MIG |
| Natural Killer | NK |
| Nerve Growth Factor | NGF |
| Neurokinin A | NKA |
| Neuropeptide Y | NPY |
| Neutrophil-activating Peptide 2 | NAP2 |
| Non-Steroidal Anti-Inflammatory Drugs | NSAIDs |
| Octenidine | OCT |
| Pathogen-associated Molecular Patterns | PAMPs |
| Pattern Recognition Receptors | PRR |
| Peroxisome Proliferator-activated Receptors | PPARs |
| Photodynamic Therapy | PDT |
| Plasmacytoid Dendritic Cells | PDCs |
| Platelet-derived Growth Factor | PDGF |
| Polyhexamethylene Biguanide | PHMB |
| Poly(butyl cyanoacrylate) | PBCA |
| Povidone–Iodine | PVP-1 |
| Protein Gene Product 9.5 | PGP 9.5 |
| Protein Kinase C | PKC |
| Reactive Oxygen Species | ROS |
| Regulated Upon Activation Normal T-Cell Expressed and Secreted | RANTES |
| Signal Transducer and Activator of Transcription 3 | STAT 3 |
| Smooth Muscle Actin | SMA |
| Sodium Hypochlorite | NaOCl |
| Stromal-Derived Growth Factor | SDF |
| Substance P | SP |
| Thrombospondin | TSP |
| Tissue Inhibitor of Metalloproteinases | TIMPs |
| Toll-like Receptor | TLR |
| Transforming Growth Factor | TGF |
| Tumor Necrosis Factor | TNF |
| Type 2 Diabetes Mellitus | T2DM |
| Tyrosinase | TYR |
| Tyrosinase-related protein 1 | TYRP1 |
| Vascular Endothelial Growth Factor | VEGF |
| Vasoactive Intestinal Peptide | VIP |
| Von Willebrand Factor | vWF |
| Wound Healing | WH |
References
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K. Human Wounds and Its Burden: An Updated Compendium of Estimates. Adv. Wound Care 2019, 8, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bacci, S. Cellular Mechanisms and Therapies in Wound Healing: Looking toward the Future. Biomedicines 2021, 9, 1611. [Google Scholar] [CrossRef]
- Gupta, S.; Andersen, C.; Black, J.; Fife, C.; Lantis, J.I.; Niezgoda, J.; Snyder, R.; Sumpio, B.; Tettelbach, W.; Treadwell, T.; et al. Management of Chronic Wounds: Diagnosis, Preparation, Treatment, and Follow-up. Wounds Compend. Clin. Res. Pract. 2017, 29, S19–S36. [Google Scholar]
- Babalska, Z.; Korbecka-Paczkowska, M.; Karpiński, T.M. Wound Antiseptics and European Guidelines for Antiseptic Application in Wound Treatment. Pharmaceuticals 2021, 14, 1253. [Google Scholar] [CrossRef]
- Takeo, M.; Lee, W.; Ito, M. Wound Healing and Skin Regeneration. Cold Spring Harb. Perspect. Med. 2015, 5, a023267. [Google Scholar] [CrossRef] [PubMed]
- Tyavambiza, C.; Meyer, M.; Meyer, S. Cellular and Molecular Events of Wound Healing and the Potential of Silver Based Nanoformulations as Wound Healing Agents. Bioengineering 2022, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Cañedo-Dorantes, L.; Cañedo-Ayala, M. Skin Acute Wound Healing: A Comprehensive Review. Int. J. Inflamm. 2019, 2019, 3706315. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef]
- Olson, T.S.; Ley, K. Chemokines and chemokine receptors in leukocyte trafficking. Am. J. Physiol. Integr. Comp. Physiol. 2002, 283, R7–R28. [Google Scholar] [CrossRef]
- Yoshida, M.; Okubo, N.; Chosa, N.; Hasegawa, T.; Ibi, M.; Kamo, M.; Kyakumoto, S.; Ishisaki, A. TGF-β-Operated Growth Inhibition and Translineage Commitment into Smooth Muscle Cells of Periodontal Ligament-Derived Endothelial Progenitor Cells through Smad- and p38 MAPK-Dependent Signals. Int. J. Biol. Sci. 2012, 8, 1062–1074. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Dong, F.; Jia, Y.; Du, H.; Dong, N.; Xu, Y.; Wang, S.; Wu, H.; Liu, Z.; Li, W. Notch Signal Regulates Corneal Endothelial-to-Mesenchymal Transition. Am. J. Pathol. 2013, 183, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.V.; Soulika, A.M. The Dynamics of the Skin’s Immune System. Int. J. Mol. Sci. 2019, 20, 1811. [Google Scholar] [CrossRef]
- Gold, M.H.; McGuire, M.; Mustoe, T.; Pusic, A.; Sachdev, M.; Waibel, J.; Murcia, C. International Advisory Panel on Scar Management Updated international clinical recommendations on scar management: Part 2—Algorithms for scar prevention and treatment. Dermatol. Surg. 2014, 40, 825–831. [Google Scholar] [PubMed]
- Wikimedia Commons contributors, “File:Wound healing phases.svg”. Wikimedia Commons. Available online: https://commons.wikimedia.org/w/index.php?title=File:Wound_healing_phases.svg&oldid=782098460 (accessed on 1 June 2023).
- Qing, C. The molecular biology in wound healing & non-healing wound. Chin. J. Traumatol. 2017, 20, 189–193. [Google Scholar] [CrossRef]
- Etulain, J. Platelets in wound healing and regenerative medicine. Platelets 2018, 29, 556–568. [Google Scholar] [CrossRef]
- Bacci, S. Fine Regulation during Wound Healing by Mast Cells, a Physiological Role Not Yet Clarified. Int. J. Mol. Sci. 2022, 23, 1820. [Google Scholar] [CrossRef]
- Brandt, E.; Petersen, F.; Ludwig, A.; Ehlert, J.; Bock, L.; Flad, H. The β-thromboglobulins and platelet factor 4: Blood platelet-derived CXC chemokines with divergent roles in early neutrophil regulation. J. Leukoc. Biol. 2000, 67, 471–478. [Google Scholar] [CrossRef]
- Gillitzer, R.; Goebeler, M. Chemokines in cutaneous wound healing. J. Leukoc. Biol. 2001, 69, 513–521. [Google Scholar] [CrossRef]
- Murdoch, C.; Finn, A. Chemokine receptors and their role in inflammation and infectious diseases. Blood 2000, 95, 3032–3043. [Google Scholar] [CrossRef]
- Yamamoto, T.; Eckes, B.; Mauch, C.; Hartmann, K.; Krieg, T. Monocyte Chemoattractant Protein-1 Enhances Gene Expression and Synthesis of Matrix Metalloproteinase-1 in Human Fibroblasts by an Autocrine IL-1α Loop. J. Immunol. 2000, 164, 6174–6179. [Google Scholar] [CrossRef] [PubMed]
- Krzyszczyk, P.; Schloss, R.; Palmer, A.; Berthiaume, F. The Role of Macrophages in Acute and Chronic Wound Healing and Interventions to Promote Pro-wound Healing Phenotypes. Front. Physiol. 2018, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Bacci, S.; Defraia, B.; Cinci, L.; Calosi, L.; Guasti, D.; Pieri, L.; Lotti, V.; Bonelli, A.; Romagnoli, P. Immunohistochemical analysis of dendritic cells in skin lesions: Correlations with survival time. Forensic Sci. Int. 2014, 244, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Chen, Y.; Zhang, Y.; Wei, A.; Zhou, J.; Li, Q.; Guo, L. MiR-21/PTEN Axis Promotes Skin Wound Healing by Dendritic Cells Enhancement. J. Cell. Biochem. 2017, 118, 3511–3519. [Google Scholar] [CrossRef] [PubMed]
- Bordon, Y. pDCs play off scratch. Nat. Rev. Immunol. 2010, 11, 5. [Google Scholar] [CrossRef]
- Nosbaum, A.; Prevel, N.; Truong, H.-A.; Mehta, P.; Ettinger, M.; Scharschmidt, T.C.; Ali, N.H.; Pauli, M.L.; Abbas, A.K.; Rosenblum, M.D. Cutting Edge: Regulatory T Cells Facilitate Cutaneous Wound Healing. J. Immunol. 2016, 196, 2010–2014. [Google Scholar] [CrossRef]
- Giantulli, S.; Tortorella, E.; Brasili, F.; Scarpa, S.; Cerroni, B.; Paradossi, G.; Bedini, A.; Morrone, S.; Silvestri, I.; Domenici, F. Effect of 1-MHz ultrasound on the proinflammatory interleukin-6 secretion in human keratinocytes. Sci. Rep. 2021, 11, 19033. [Google Scholar] [CrossRef]
- Piipponen, M.; Li, D.; Landén, N.X. The Immune Functions of Keratinocytes in Skin Wound Healing. Int. J. Mol. Sci. 2020, 21, 8790. [Google Scholar] [CrossRef]
- Bacci, S. The Epidermis in Microgravity and Unloading Conditions and Their Effects on Wound Healing. Front. Bioeng. Biotechnol. 2022. [CrossRef]
- Rennekampff, H.O.; Hansbrough, J.F.; Kiessig, V.; Dore, C.; Sticherling, M.; Schroder, J.M. Bioactive interleukin-8 is expressed in wounds and enhances wound healing. J. Surg. Res. 2000, 93, 41–54. [Google Scholar] [CrossRef]
- Engelhardt, E.; Toksoy, A.; Goebeler, M.; Debus, S.; Bröcker, E.-B.; Gillitzer, R. Chemokines IL-8, GROα, MCP-1, IP-10, and Mig Are Sequentially and Differentially Expressed during Phase-Specific Infiltration of Leukocyte Subsets in Human Wound Healing. Am. J. Pathol. 1998, 153, 1849–1860. [Google Scholar] [CrossRef]
- Kulke, R.; Bornscheuer, E.; Schlüter, C.; Bartels, J.; Röwert, J.; Sticherling, M.; Christophers, E. The CXC Receptor 2 Is Overexpressed in Psoriatic Epidermis. J. Investig. Dermatol. 1998, 110, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Goebeler, M.; Yoshimura, T.; Toksoy, A.; Ritter, U.; Bröcker, E.-B.; Gillitzer, R. The Chemokine Repertoire of Human Dermal Microvascular Endothelial Cells and Its Regulation by Inflammatory Cytokines. J. Investig. Dermatol. 1997, 108, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Strieter, R.M.; Polverini, P.J.; Kunkel, S.L.; Arenberg, D.A.; Burdick, M.D.; Kasper, J.; Dzuiba, J.; Van Damme, J.; Walz, A.; Marriott, D.; et al. The Functional Role of the ELR Motif in CXC Chemokine-mediated Angiogenesis. J. Biol. Chem. 1995, 270, 27348–27357. [Google Scholar] [CrossRef]
- Belperio, J.; Keane, M.P.; Arenberg, D.; Addison, C.L.; E Ehlert, J.; Burdick, M.D.; Strieter, R.M. CXC chemokines in angiogenesis. J. Leukoc. Biol. 2000, 68, 1–8. [Google Scholar] [CrossRef]
- Randolph, G.J.; Furie, M.B. A soluble gradient of endogenous monocyte chemoattractant protein-1 promotes the transendothelial migration of monocytes in vitro. Perspect. Surg. 1995, 155, 3610–3618. [Google Scholar] [CrossRef]
- Morbidelli, L.; Genah, S.; Cialdai, F. Effect of Microgravity on Endothelial Cell Function, Angiogenesis, and Vessel Remodeling During Wound Healing. Front. Bioeng. Biotechnol. 2021, 9, 720091. [Google Scholar] [CrossRef]
- Johnson, K.E.; Wilgus, T.A.; Perez-Amodio, S.; Rubio, N.; Vila, O.F.; Navarro-Requena, C.; Castaño, O.; Sanchez-Ferrero, A.; Marti-Munoz, J.; Alsina-Giber, M.; et al. Vascular Endothelial Growth Factor and Angiogenesis in the Regulation of Cutaneous Wound Repair. Adv. Wound Care 2014, 3, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J.; Satish, L.; Yates, C.C.; Wells, A. Pericytes: A newly recognized player in wound healing. Wound Repair Regen. 2016, 24, 204–214. [Google Scholar] [CrossRef]
- Cialdai, F.; Risaliti, C.; Monici, M. Role of fibroblasts in wound healing and tissue remodeling on Earth and in space. Front. Bioeng. Biotechnol. 2022, 10, 958381. [Google Scholar] [CrossRef]
- Monika, P.; Waiker, P.V.; Chandraprabha, M.N.; Rangarajan, A.; Murthy, K.N.C. Myofibroblast progeny in wound biology and wound healing studies. Wound Repair Regen. 2021, 29, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Schultz, G.S.; Chin, G.A.; Moldawer, L.; Diegelman, R.F. Principles of wound healing. In Mechanisms of Vascular Disease: A Reference Book for Vascular Specialists; Fitridge, R., Thompson, M., Eds.; University of Adelaide Press: Adelaide, Australia, 2011. Available online: https://www.ncbi.nlm.nih.gov/books/NBK534261 (accessed on 1 June 2023).
- Palmieri, B.; Vadalà, M.; Laurino, C. Review of the molecular mechanisms in wound healing: New therapeutic targets? J. Wound Care 2017, 26, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-J.; Fan, M.; Gao, W. Identification of potential hub genes associated with skin wound healing based on time course bioinformatic analyses. BMC Surg. 2021, 21, 303. [Google Scholar] [CrossRef] [PubMed]
- Raziyeva, K.; Kim, Y.; Zharkinbekov, Z.; Kassymbek, K.; Jimi, S.; Saparov, A. Immunology of Acute and Chronic Wound Healing. Biomolecules 2021, 11, 700. [Google Scholar] [CrossRef] [PubMed]
- Falanga, V.; Isseroff, R.R.; Soulika, A.M.; Romanelli, M.; Margolis, D.; Kapp, S.; Granick, M.; Harding, K. Chronic wounds. Nat. Rev. Dis. Prim. 2022, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.; Nadkarni, S.; Lele, J.; Sakhalkar, S.; Mokashi, P.; Kaushik, K.S. Bioengineered Platforms for Chronic Wound Infection Studies: How Can We Make Them More Human-Relevant? Front. Bioeng. Biotechnol. 2019, 7, 418. [Google Scholar] [CrossRef]
- Grandi, V.; Corsi, A.; Pimpinelli, N.; Bacci, S. Cellular Mechanisms in Acute and Chronic Wounds after PDT Therapy: An Update. Biomedicines 2022, 10, 1624. [Google Scholar] [CrossRef]
- Tottoli, E.M.; Dorati, R.; Genta, I.; Chiesa, E.; Pisani, S.; Conti, B. Skin Wound Healing Process and New Emerging Technologies for Skin Wound Care and Regeneration. Pharmaceutics 2020, 12, 735. [Google Scholar] [CrossRef]
- Holl, J.; Kowalewski, C.; Zimek, Z.; Fiedor, P.; Kaminski, A.; Oldak, T.; Moniuszko, M.; Eljaszewicz, A. Chronic Diabetic Wounds and Their Treatment with Skin Substitutes. Cells 2021, 10, 655. [Google Scholar] [CrossRef]
- Andrews, J.P.; Marttala, J.; Macarak, E.; Rosenbloom, J.; Uitto, J. Keloids: The paradigm of skin fibrosis—Pathomechanisms and treatment. Matrix Biol. 2016, 51, 37–46. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, X.; Wang, W.; Zhou, G.; Zhang, W.; Gao, Z.; Wu, X.; Liu, W. Combined analyses of RNA-sequence and Hi-C along with GWAS loci—A novel approach to dissect keloid disorder genetic mechanism. PLOS Genet. 2022, 18, e1010168. [Google Scholar] [CrossRef]
- Lingzhi, Z.; Meirong, L.; Xiaobing, F. Biological approaches for hypertrophic scars. Int. Wound J. 2019, 17, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Rinkevich, Y. Scars or Regeneration?—Dermal Fibroblasts as Drivers of Diverse Skin Wound Responses. Int. J. Mol. Sci. 2020, 21, 617. [Google Scholar] [CrossRef] [PubMed]
- Berman, B.; Maderal, A.; Raphael, B. Keloids and Hypertrophic Scars: Pathophysiology, Classification, and Treatment. Dermatol. Surg. 2017, 43 (Suppl. S1), S3–S18. [Google Scholar] [CrossRef]
- Leszczynski, R.; da Silva, C.A.P.; Pinto, A.C.P.N.; Kuczynski, U.; da Silva, E.M. Laser therapy for treating hypertrophic and keloid scars. Cochrane Database Syst. Rev. 2022, 2022, CD011642. [Google Scholar] [CrossRef]
- Alghamdi, M.A.; Wallace, H.J.; Melton, P.E.; Moses, E.K.; Stevenson, A.; Al-Eitan, L.N.; Rea, S.; Duke, J.M.; Danielsen, P.L.; Prêle, C.M.; et al. Identification of Differentially Methylated CpG Sites in Fibroblasts from Keloid Scars. Biomedicines 2020, 8, 181. [Google Scholar] [CrossRef]
- Magni, G.; Banchelli, M.; Cherchi, F.; Coppi, E.; Fraccalvieri, M.; Rossi, M.; Tatini, F.; Pugliese, A.M.; Degl’innocenti, D.R.; Alfieri, D.; et al. Experimental Study on Blue Light Interaction with Human Keloid-Derived Fibroblasts. Biomedicines 2020, 8, 573. [Google Scholar] [CrossRef]
- Yang, R.; Wang, J.; Chen, X.; Shi, Y.; Xie, J. Epidermal Stem Cells in Wound Healing and Regeneration. Stem Cells Int. 2020, 2020, 9148310. [Google Scholar] [CrossRef]
- Ma, H.; Siu, W.-S.; Leung, P.-C. The Potential of MSC-Based Cell-Free Therapy in Wound Healing—A Thorough Literature Review. Int. J. Mol. Sci. 2023, 24, 9356. [Google Scholar] [CrossRef]
- Kua, J.E.H.; Siow, C.W.; Lim, W.K.; Masilamani, J.; Tjin, M.S.; Yeong, J.; Lim, T.K.H.; Phan, T.T.; Chua, A.W.C. Human Umbilical Cord Lining-Derived Epithelial Cells: A Potential Source of Non-Native Epithelial Cells That Accelerate Healing in a Porcine Cutaneous Wound Model. Int. J. Mol. Sci. 2022, 23, 8918. [Google Scholar] [CrossRef]
- Khalil, H.; Cullen, M.; Chambers, H.; McGrail, M. Medications affecting healing: An evidence-based analysis. Int. Wound J. 2017, 14, 1340–1345. [Google Scholar] [CrossRef] [PubMed]
- Bacci, S.; Laurino, A.; Manni, M.E.; Landucci, E.; Musilli, C.; De Siena, G.; Mocali, A.; Raimondi, L. The pro-healing effect of exendin-4 on wounds produced by abrasion in normoglycemic mice. Eur. J. Pharmacol. 2015, 764, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Paroli, G.; Murciano, N.; Mancini, C.; Soldaini, M.; Rijli, S.; DeSiena, G.; Bacci, S. The role of mast cells in cellular modifications evoked by Exendin-4 in treated wounds: A preclinical study. J. Wound Care 2022, 31, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Guarino, M.F.; Fernández-Nieto, D.; Montes, L.V.; Lobo, D.d.P. Methyl Aminolaevulinic Acid versus Aminolaevulinic Acid Photodynamic Therapy of Actinic Keratosis with Low Doses of Red-Light LED Illumination: Results of Long-Term Follow-Up. Biomedicines 2022, 10, 3218. [Google Scholar] [CrossRef] [PubMed]
- França, C.M.; Anders, J.J.; Lanzafame, R.J. Photobiomodulation in Wound Healing: What Are We Not Considering? Photomed. Laser Surg. 2016, 34, 51–52. [Google Scholar] [CrossRef]
- Cicchi, R.; Rossi, F.; Alfieri, D.; Bacci, S.; Tatini, F.; De Siena, G.; Paroli, G.; Pini, R.; Pavone, F.S. Observation of an improved healing process in superficial skin wounds after irradiation with a blue-LED haemostatic device. J. Biophotonics 2016, 9, 645–655. [Google Scholar] [CrossRef]
- Magni, G.; Tatini, F.; Bacci, S.; Paroli, G.; De Siena, G.; Cicchi, R.; Pavone, F.S.; Pini, R.; Rossi, F. Blue LED light modulates inflammatory infiltrate and improves the healing of superficial wounds. Photodermatol. Photoimmunol. Photomed. 2019, 36, 166–168. [Google Scholar] [CrossRef]
- Magni, G.; Tatini, F.; De Siena, G.; Pavone, F.S.; Alfieri, D.; Cicchi, R.; Rossi, M.; Murciano, N.; Paroli, G.; Vannucci, C.; et al. Blue-LED-Light Photobiomodulation of Inflammatory Responses and New Tissue Formation in Mouse-Skin Wounds. Life 2022, 12, 1564. [Google Scholar] [CrossRef]
- Nguyen, J.K.; Weedon, J.; Jakus, J.; Heilman, E.; Isseroff, R.R.; Siegel, D.M.; Jagdeo, J.R. A dose-ranging, parallel group, split-face, single-blind phase II study of light emitting diode-red light (LED-RL) for skin scarring prevention: Study protocol for a randomized controlled trial. Trials 2019, 20, 432. [Google Scholar] [CrossRef]
- Polak, A.; Kucio, C.; Kloth, L.; Paczula, M.; Hordynska, E.; Ickowicz, T.; Blaszczak, E.; Kucio, E.; Oleszczyk, K.; Ficek, K.; et al. A Randomized, Controlled Clinical Study to Assess the Effect of Anodal and Cathodal Electrical Stimulation on Periwound Skin Blood Flow and Pressure Ulcer Size Reduction in Persons with Neurological Injuries. J. Wound Ostomy Cont. Nurs. 2018, 64, 10–29. [Google Scholar] [CrossRef]
- Alkahtani, S.A.; Kunwar, P.S.; Jalilifar, M.; Rashidi, S.; Yadollahpour, A. Ultrasound-based Techniques as Alternative Treatments for Chronic Wounds: A Comprehensive Review of Clinical Applications. Cureus 2017, 9, e1952. [Google Scholar] [CrossRef] [PubMed]
- Karsli, P.B.; Gurcay, E.; Karaahmet, O.Z.; Cakci, A. High-Voltage Electrical Stimulation versus Ultrasound in the Treatment of Pressure Ulcers. Adv. Ski. Wound Care 2017, 30, 565–570. [Google Scholar] [CrossRef]
- Bianchi, E.; Vigani, B.; Ruggeri, M.; Del Favero, E.; Ricci, C.; Grisoli, P.; Ferraretto, A.; Rossi, S.; Viseras, C.; Sandri, G. Electrospun Scaffolds Based on Poly(butyl cyanoacrylate) for Tendon Tissue Engineering. Int. J. Mol. Sci. 2023, 24, 3172. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Guo, X.; Mo, F.; Sun, D. Different Dimensional Copper-Based Metal–Organic Frameworks with Enzyme-Mimetic Activity for Antibacterial Therapy. Int. J. Mol. Sci. 2023, 24, 3173. [Google Scholar] [CrossRef] [PubMed]
- Bąchor, U.; Junka, A.; Brożyna, M.; Mączyński, M. The In Vitro Impact of Isoxazole Derivatives on Pathogenic Biofilm and Cytotoxicity of Fibroblast Cell Line. Int. J. Mol. Sci. 2023, 24, 2997. [Google Scholar] [CrossRef]
- Di Lodovico, S.; Diban, F.; Di Fermo, P.; Petrini, M.; Fontana, A.; Di Giulio, M.; Piattelli, A.; D’ercole, S.; Cellini, L. Antimicrobial Combined Action of Graphene Oxide and Light Emitting Diodes for Chronic Wound Management. Int. J. Mol. Sci. 2022, 23, 6942. [Google Scholar] [CrossRef]
- Sousa, P.; Lopes, B.; Sousa, A.C.; Moreira, A.; Coelho, A.; Alvites, R.; Alves, N.; Geuna, S.; Maurício, A.C. Advancements and Insights in Exosome-Based Therapies for Wound Healing: A Comprehensive Systematic Review (2018–June 2023). Biomedicines 2023, 11, 2099. [Google Scholar] [CrossRef]
- Cialdai, F.; Bacci, S.; Zizi, V.; Norfini, A.; Balsamo, M.; Ciccone, V.; Morbidelli, L.; Calosi, L.; Risaliti, C.; Vanhelden, L.; et al. Optimization of an Ex-Vivo Human Skin/Vein Model for Long-Term Wound Healing Studies: Ground Preparatory Activities for the ‘Suture in Space’ Experiment Onboard the International Space Station. Int. J. Mol. Sci. 2022, 23, 14123. [Google Scholar] [CrossRef]
- Wang, B.; Nolan, R.; Marshall, H. COVID-19 Immunisation, Willingness to Be Vaccinated and Vaccination Strategies to Improve Vaccine Uptake in Australia. Vaccines 2021, 9, 1467. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Guarino, M.; Hernández-Bule, M.L.; Bacci, S. Cellular and Molecular Processes in Wound Healing. Biomedicines 2023, 11, 2526. https://doi.org/10.3390/biomedicines11092526
Fernández-Guarino M, Hernández-Bule ML, Bacci S. Cellular and Molecular Processes in Wound Healing. Biomedicines. 2023; 11(9):2526. https://doi.org/10.3390/biomedicines11092526
Chicago/Turabian StyleFernández-Guarino, Montserrat, Maria Luisa Hernández-Bule, and Stefano Bacci. 2023. "Cellular and Molecular Processes in Wound Healing" Biomedicines 11, no. 9: 2526. https://doi.org/10.3390/biomedicines11092526
APA StyleFernández-Guarino, M., Hernández-Bule, M. L., & Bacci, S. (2023). Cellular and Molecular Processes in Wound Healing. Biomedicines, 11(9), 2526. https://doi.org/10.3390/biomedicines11092526