
Role of Fork-Head Box Genes in Breast Cancer: From Drug Resistance to Therapeutic Targets
 , , , , and
, , , , and
Abstract
1. Introduction
2. Role of Various Forkhead Boxes in Tumor Proliferation and Progression
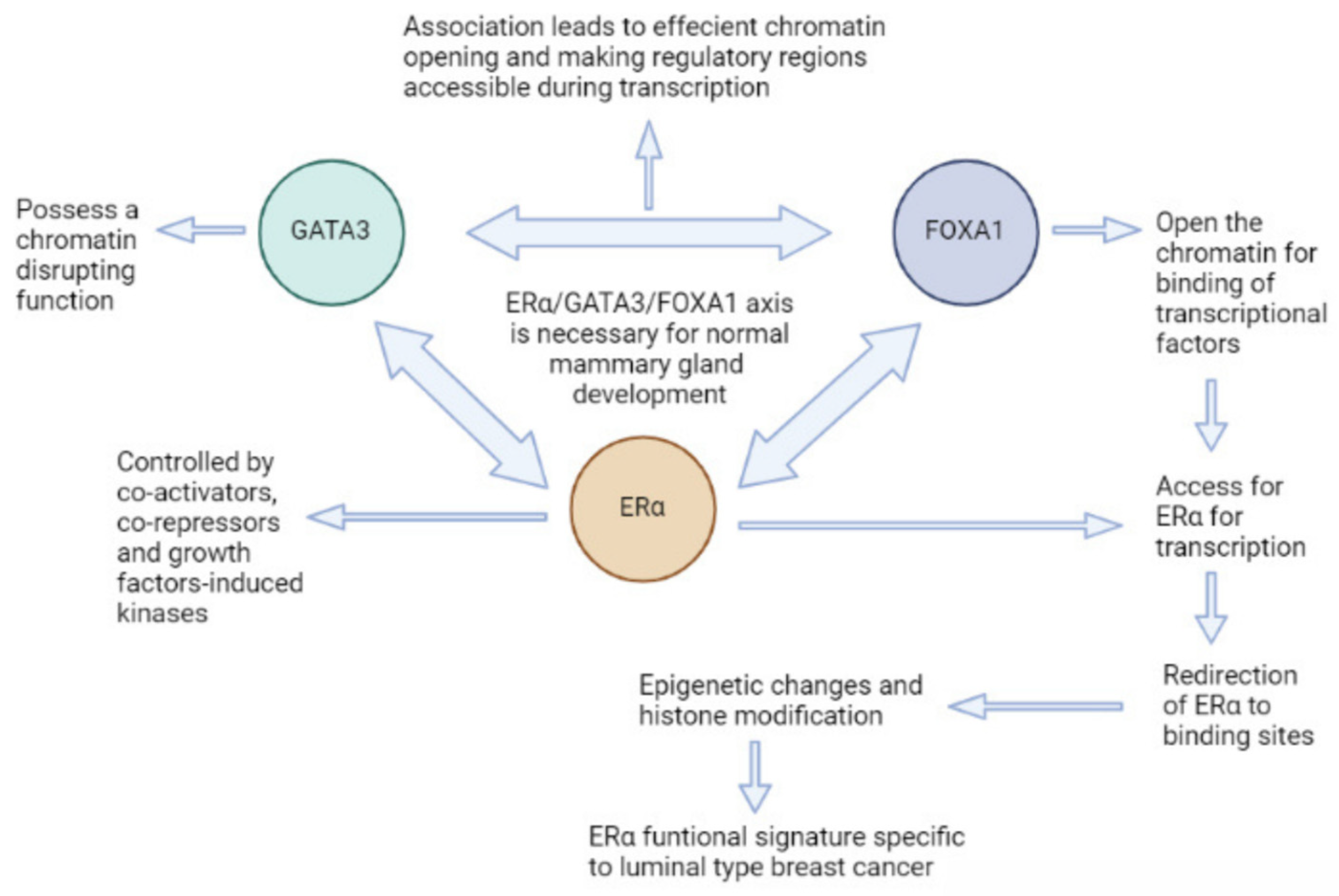
2.1. Forkhead Box A in Cancer
2.2. Forkhead Box M1 in Cancer
2.3. Forkhead Box O in Cancer
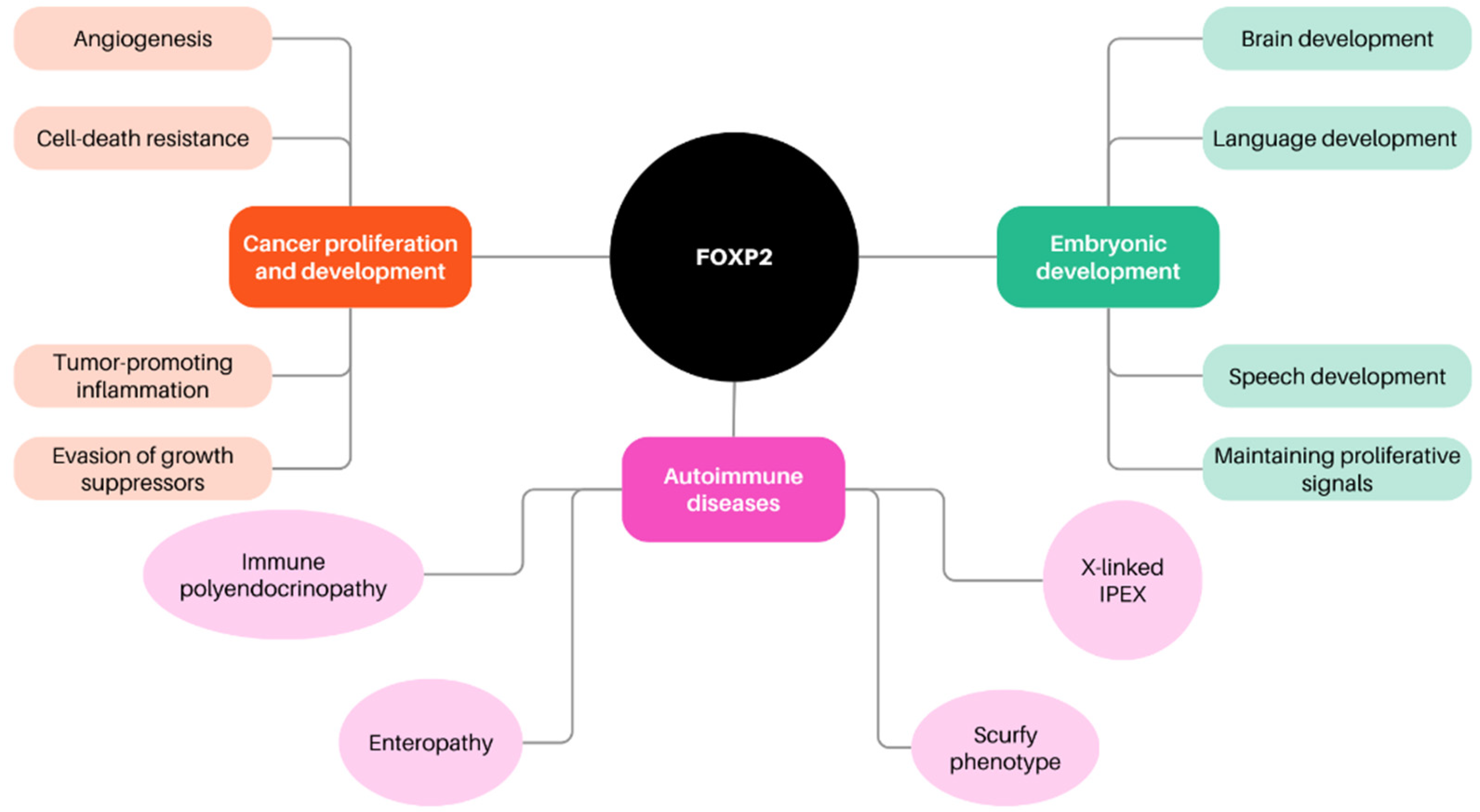
2.4. Forkhead Box P in Cancer
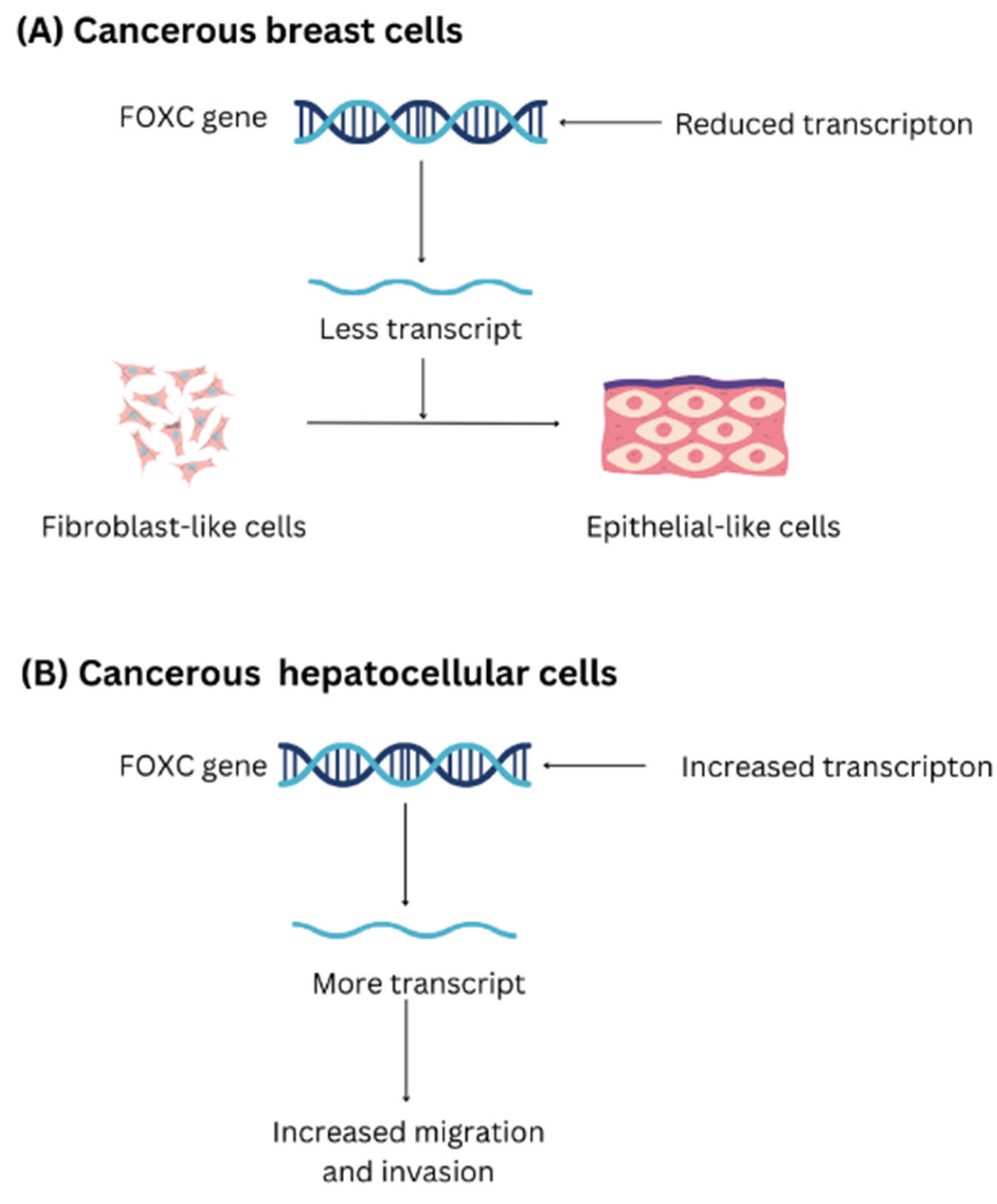
2.5. Forkhead Box C in Cancer
2.6. FOX Proteins in Cancer Drug Resistance
2.7. Negative Regulation of FOX Proteins by miRNAs
2.8. FOX Family and Stem Cells
2.9. Members of the FOX Family as Drug Targets for Cancer Therapies
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carlsson, P.; Mahlapuu, M. Forkhead transcription factors: Key players in development and metabolism. Dev. Biol. 2002, 250, 1–23. [Google Scholar] [CrossRef]
- Lam, E.W.F.; Brosens, J.J.; Gomes, A.R.; Koo, C.-Y. Forkhead box proteins: Tuning forks for transcriptional harmony. Nat. Rev. Cancer 2013, 13, 482. [Google Scholar] [CrossRef]
- Coffer, P.J.; Burgering, B.M.T. Forkhead-box transcription factors and their role in the immune system. Nat. Rev. Immunol. 2004, 4, 889. [Google Scholar] [CrossRef]
- Jackson, B.C.; Carpenter, C.; Nebert, D.W.; Vasiliou, V. Update of human and mouse forkhead box [FOX] gene families. Hum. Genom. 2010, 4, 345–352. [Google Scholar] [CrossRef]
- Choi, E.J.; Seo, E.J.; Kim, D.K.; Lee, S.I.; Kwon, Y.W.; Jang, I.H.; Kim, K.H.; Suh, D.S.; Kim, J.H. FOXP1 functions as an oncogene in promoting cancer stem cell-like characteristics in ovarian cancer cells. Oncotarget 2016, 7, 3506–3519. [Google Scholar] [CrossRef]
- Lam, E.W.-F.; Gomes, A.R. Forkhead box transcription factors in cancer initiation, progression and chemotherapeutic drug response. Front. Oncol. 2014, 4, 305. [Google Scholar] [CrossRef]
- Russell, E.G.; Cotter, T.G. Chapter Six—New insight into the role of reactive oxygen species [ROS] in cellular signal-transduction processes. Int. Rev. Cell Mol. Biol. 2015, 319, 221–254. [Google Scholar]
- Bach, D.H.; Long, N.P.; Luu, T.T.; Anh, N.H.; Kwon, S.W.; Lee, S.K. The Dominant Role of Forkhead Box Proteins in Cancer. Int. J. Mol. Sci. 2018, 19, 3279. [Google Scholar] [CrossRef]
- Ali, S.; Coombes, R.C. Endocrine-responsive breast cancer and strategies for combating resistance. Nat. Rev. Cancer 2002, 2, 101–112. [Google Scholar] [CrossRef]
- Nakshatri, H.; Badve, S. FOXA1 as a therapeutic target for breast cancer. Expert Opin. Ther. Targets 2007, 11, 507–514. [Google Scholar] [CrossRef]
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef]
- Friedman, J.R.; Kaestner, K.H. The FOXA family of transcription factors in development and metabolism. Cell Mol. Life Sci. 2006, 63, 2317–2328. [Google Scholar] [CrossRef]
- Jägle, S.; Busch, H.; Freihen, V.; Beyes, S.; Schrempp, M.; Boerries, M.; Hecht, A. SNAIL1-mediated downregulation of FOXA proteins facilitates the inactivation of transcriptional enhancer elements at key epithelial genes in colorectal cancer cells. PLoS Genet. 2017, 13, e1007109. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, Y.; Liu, Y.; Kao, L.-P.; Wang, X.; Skerry, B.; Li, Z. FOXA1 defines cancer cell specificity. Sci. Adv. 2016, 2, e1501473. [Google Scholar] [CrossRef]
- Lin, L.; Miller, C.T.; Contreras, J.I.; Prescott, M.S.; Dagenais, S.L.; Wu, R.; Yee, J.; Orringer, M.B.; Misek, D.E.; Hanash, S.M.; et al. The hepatocyte nuclear factor 3 alpha gene, HNF3alpha [FOXA1], on chromosome band 14q13 is amplified and overexpressed in esophageal and lung adenocarcinomas. Cancer Res. 2002, 62, 5273. [Google Scholar]
- DeGraff, D.J.; Clark, P.E.; Cates, J.M.; Yamashita, H.; Robinson, V.L.; Yu, X.; Smolkin, M.E.; Chang, S.S.; Cookson, M.S.; Herrick, M.K.; et al. Loss of the urothelial differentiation marker FOXA1 is associated with high grade, late stage bladder cancer and increased tumor proliferation. PLoS ONE 2012, 7, e36669. [Google Scholar] [CrossRef]
- Mirosevich, J.; Gao, N.; Gupta, A.; Shappell, S.B.; Jove, R.; Matusik, R.J. Expression and role of Foxa proteins in prostate cancer. Prostate 2006, 66, 1013–1028. [Google Scholar] [CrossRef]
- Lacroix, M.; Leclercq, G. About GATA3, HNF3A, and XBP1, three genes co-expressed with the oestrogen receptor-α gene [ESR1] in breast cancer. Mol. Cell. Endocrinol. 2004, 219, 1–7. [Google Scholar] [CrossRef]
- Oh, D.S.; Troester, M.A.; Usary, J.; Hu, Z.; He, X.; Fan, C.; Wu, J.; Carey, L.A.; Perou, C.M. Estrogen-regulated genes predict survival in hormone receptor-positive breast cancers. J. Clin. Oncol. 2006, 24, 1656–1664. [Google Scholar] [CrossRef]
- Carroll, J.S.; Brown, M. Estrogen receptor target gene: An evolving concept. Mol. Endocrinol. 2006, 20, 1707–1714. [Google Scholar] [CrossRef]
- Xia, W.; Bacus, S.; Hegde, P.; Husain, I.; Strum, J.; Liu, L.; Paulazzo, G.; Lyass, L.; Trusk, P.; Hill, J.; et al. A model of acquired autoresistance to a potent ErbB2 tyrosine kinase inhibitor and a therapeutic strategy to prevent its onset in breast cancer. Proc. Natl. Acad. Sci. USA 2006, 103, 7795–7800. [Google Scholar] [CrossRef] [PubMed]
- Raychaudhuri, P.; Park, H.J. FOXM1: A master regulator of tumor metastasis. Cancer Res. 2011, 71, 4329. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, D.; Yu, Q.; Li, L.; Wu, P. Prognostic value of FOXM1 in solid tumors: A systematic review and meta-analysis. Oncotarget 2017, 8, 32298–32308. [Google Scholar] [CrossRef]
- Park, H.J.; Carr, J.R.; Wang, Z.; Nogueira, V.; Hay, N.; Tyner, A.L.; Lau, L.F.; Costa, R.H.; Raychaudhuri, P. FOXM1, a critical regulator of oxidative stress during oncogenesis. EMBO J. 2009, 28, 2908. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Gusarova, G.; Wang, Z.; Carr, J.R.; Li, J.; Kim, K.H.; Qiu, J.; Park, Y.D.; Williamson, P.R.; Hay, N.; et al. Deregulation of FOXM1B leads to tumour metastasis. EMBO Mol. Med. 2011, 3, 21. [Google Scholar] [CrossRef]
- Wang, I.C.; Chen, Y.-J.; Hughes, D.; Petrovic, V.; Major, M.L.; Park, H.J.; Tan, Y.; Ackerson, T.; Costa, R.H. Forkhead box M1 regulates the transcriptional network of genes essential for mitotic progression and genes encoding the SCF [Skp2-Cks1] ubiquitin ligase. Mol. Cell Biol. 2005, 25, 10875. [Google Scholar] [CrossRef]
- Radhakrishnan, S.K.; Gartel, A.L. FOXM1: The Achilles’ heel of cancer? Nat. Rev. Cancer 2008, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- O’Regan, R.M.; Nahta, R. Targeting Forkhead Box M1 Transcription Factor in Breast Cancer. Biochem. Pharmacol. 2018, 154, 407–413. [Google Scholar] [CrossRef]
- Wonsey, D.R.; Follettie, M.T. Loss of the forkhead transcription factor FoxM1 causes centrosome amplification and mitotic catastrophe. Cancer Res. 2005, 65, 5181–5189. [Google Scholar] [CrossRef]
- Bektas, N.; Haaf, A.T.; Veeck, J.; Wild, P.J.; Lüscher-Firzlaff, J.; Hartmann, A.; Knüchel, R.; Dahl, E. Tight correlation between expression of the Forkhead transcription factor FOXM1 and HER2 in human breast cancer. BMC Cancer 2008, 8, 42. [Google Scholar] [CrossRef]
- Madureira, P.A.; Varshochi, R.; Constantinidou, D.; Francis, R.E.; Coombes, R.C.; Yao, K.-M.; Lam, E.W.-F. The Forkhead box M1 protein regulates the transcription of the estrogen receptor alpha in breast cancer cells. J. Biol. Chem. 2006, 281, 25167–25176. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.A.; Ross-Innes, C.S.; Beraldi, D.; Carroll, J.S.; Balasubramanian, S. Genome-wide mapping of FOXM1 binding reveals co-binding with estrogen receptor alpha in breast cancer cells. Genome Biol. 2013, 14, R6. [Google Scholar] [CrossRef] [PubMed]
- Horimoto, Y.; Hartman, J.; Millour, J.; Pollock, S.; Olmos, Y.; Ho, K.K.; Coombes, R.C.; Poutanen, M.; Mäkelä, S.I.; El-Bahrawy, M.; et al. ERbeta1 represses FOXM1 expression through targeting ERalpha to control cell proliferation in breast cancer. Am. J. Pathol. 2011, 179, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.F.; Zeng; Liang, W.Q.; Chen, C.F.; Sun, S.M.; Lin, H.Y. FoxM1 is a promising candidate target in the treatment of breast cancer. Oncotarget 2018, 9, 842–852. [Google Scholar] [CrossRef]
- Lee, J.J.; Lee, H.J.; Son, B.H.; Kim, S.B.; Ahn, J.-H.; Ahn, S.D.; Cho, E.Y.; Gong, G. Expression of FOXM1 and related proteins in breast cancer molecular subtypes. Int. J. Exp. Pathol. 2016, 97, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Coomans de Brachène, A.; Demoulin, J.B. FOXO transcription factors in cancer development and therapy. Cell Mol. Life Sci. 2016, 73, 1159–1172. [Google Scholar] [CrossRef]
- Huang, H.; Tindall, D.J. Dynamic FOXO transcription factors. J. Cell Sci. 2007, 120, 2479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gan, B.; Liu, D.; Paik, J.H. FOXO family members in cancer. Cancer Biol. Ther. 2011, 12, 253–259. [Google Scholar] [CrossRef]
- Greer, E.L.; Brunet, A. FOXO transcription factors in ageing and cancer. Acta Physiol. 2007, 192, 19–28. [Google Scholar] [CrossRef]
- Kloet, D.E.A.; Burgering, B.M.T. The PKB/FOXO switch in aging and cancer. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 1926–1937. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z.; Shang, Y.C.; Hou, J. Clever cancer strategies with FOXO transcription factors. Cell Cycle 2008, 7, 3829–3839. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Chauhan, A.S.; Zhuang, L.; Gan, B. FOXO transcription factors in cancer metabolism. Semin. Cancer Biol. 2018, 50, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Sin, C.; Li, H.; Crawford, D.A. Transcriptional regulation by FOXP1, FOXP2, and FOXP4 dimerization. J. Mol. Neurosci. 2015, 55, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Meerschaut, I.; Rochefort, D.; Revençu, N.; Pètre, J.; Corsello, C.; Rouleau, G.A.; Hamdan, F.F.; Michaud, J.L.; Morton, J.; Radley, J.; et al. FOXP1-related intellectual disability syndrome: A recognisable entity. J. Med. Genet. 2017, 54, 613. [Google Scholar] [CrossRef]
- Tao, J.-H.; Cheng, M.; Tang, J.-P.; Liu, Q.; Pan, F.; Li, X.-P. FOXP3, regulatory T cell, and autoimmune diseases. Inflammation 2017, 40, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Ladoire, S.; Mignot, G.; Apetoh, L.; Ghiringhelli, F. Human FOXP3 and cancer. Oncogene 2010, 29, 4121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Duan, N.; Song, T.; Li, Z.; Zhang, C.; Chen, X. The emerging roles of forkhead box [FOX] proteins in osteosarcoma. J. Cancer 2017, 8, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Koon, H.B.; Ippolito, G.C.; Banham, A.H.; Tucker, P.W. FOXP1: A potential therapeutic target in cancer. Expert Opin. Ther. Targets 2007, 11, 955–965. [Google Scholar] [CrossRef]
- Szylberg, L.; Karbownik, D.; Marszalek, A. The role of FOXP3 in human cancers. Anticancer. Res. 2016, 36, 3789–3794. [Google Scholar]
- Kume, T. The cooperative roles of FOXC1 and FOXC2 in cardiovascular development. Adv. Exp. Med. Biol. 2009, 665, 63–77. [Google Scholar]
- Papanicolaou, K.N.; Izumiya, Y.; Walsh, K. Forkhead transcription factors and cardiovascular biology. Circ. Res. 2008, 102, 16–31. [Google Scholar] [CrossRef]
- Elian, F.A.; Yan, E.; Walter, M.A. FOXC1, the new player in the cancer sandbox. Oncotarget 2018, 9, 8165–8178. [Google Scholar] [CrossRef]
- Han, B.; Bhowmick, N.; Qu, Y.; Chung, S.; Giuliano, A.E.; Cui, X. FOXC1: An emerging marker and therapeutic target for cancer. Oncogene 2017, 36, 3957. [Google Scholar] [CrossRef]
- Ray, P.S.; Wang, J.; Qu, Y.; Sim, M.-S.; Shamonki, J.; Bagaria, S.P.; Ye, X.; Liu, B.; Elashoff, D.; Hoon, D.S.; et al. FOXC1 is a potential prognostic biomarker with functional significance in basal-like breast cancer. Cancer Res. 2010, 70, 3870. [Google Scholar] [CrossRef]
- Xia, L.; Huang, W.; Tian, D.; Zhu, H.; Qi, X.; Chen, Z.; Zhang, Y.; Hu, H.; Fan, D.; Nie, Y.; et al. Overexpression of forkhead box C1 promotes tumor metastasis and indicates poor prognosis in hepatocellular carcinoma. Hepatology 2012, 57, 610–624. [Google Scholar] [CrossRef]
- Wang, T.; Zheng, L.; Wang, Q.; Hu, Y.W. Emerging roles and mechanisms of FOXC2 in cancer. Clin. Chim. Acta 2018, 479, 84–93. [Google Scholar] [CrossRef]
- Yao, S.; Fan, L.Y.; Lam, E.W. The FOXO3-FOXM1 axis: A key cancer drug target and a modulator of cancer drug resistance. Semin. Cancer Biol. 2018, 50, 77–89. [Google Scholar] [CrossRef]
- Bach, D.-H.; Lee, S.K. Long noncoding RNAs in cancer cells. Cancer Lett. 2018, 419, 152–166. [Google Scholar] [CrossRef]
- Duc-Hiep, B.; Lee, S.K. The potential impacts of tylophora alkaloids and their derivatives in modulating inflammation, viral infections, and cancer. Curr. Med. Chem. 2018, 26, 4709–4725. [Google Scholar]
- Khongkow, P.; Karunarathna, U.; Khongkow, M.; Gong, C.; Gomes, A.R.; Yague, E.; Monteiro, L.J.; Kongsema, M.; Zona, S.; Man, E.P.; et al. FOXM1 targets NBS1 to regulate DNA damage-induced senescence and epirubicin resistance. Oncogene 2014, 33, 4144–4155. [Google Scholar] [CrossRef]
- Nestal de Moraes, G.; Delbue, D.; Silva, K.L.; Robaina, M.C.; Khongkow, P.; Gomes, A.R.; Zona, S.; Crocamo, S.; Mencalha, A.L.; Magalhaes, L.M.; et al. FOXM1 targets XIAP and Survivin to modulate breast cancer survival and chemoresistance. Cell. Signal. 2015, 27, 2496–2505. [Google Scholar] [CrossRef] [PubMed]
- Khongkow, P.; Gomes, A.R.; Gong, C.; Man, E.P.; Tsang, J.W.; Zhao, F.; Monteiro, L.J.; Coombes, R.C.; Medema, R.H.; Khoo, U.S.; et al. Paclitaxel targets FOXM1 to regulate KIF20A in mitotic catastrophe and breast cancer paclitaxel resistance. Oncogene 2016, 35, 990–1002. [Google Scholar] [CrossRef]
- Yu-Rice, Y.; Jin, Y.; Han, B.; Qu, Y.; Johnson, J.; Watanabe, T.; Cheng, L.; Deng, N.; Tanaka, H.; Gao, B.; et al. FOXC1 is involved in ERalpha silencing by counteracting GATA3 binding and is implicated in endocrine resistance. Oncogene 2016, 35, 5400–5411. [Google Scholar] [CrossRef]
- Meng, F.; Speyer, C.L.; Zhang, B.; Zhao, Y.; Chen, W.; Gorski, D.H.; Miller, F.R.; Wu, G. PDGFRalpha and beta play critical roles in mediating FOXQ1-driven breast cancer stemness and chemoresistance. Cancer Res. 2015, 75, 584–593. [Google Scholar] [CrossRef]
- Zhao, Y.F.; Zhao, J.Y.; Yue, H.; Hu, K.S.; Shen, H.; Guo, Z.G.; Su, X.J. FOXD1 promotes breast cancer proliferation and chemotherapeutic drug resistance by targeting p27. Biochem. Biophys. Res. Commun. 2015, 456, 232–237. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Nakayama, Y.; Yamaguchi, N. Down-regulation of Forkhead box protein A1 [FOXA1] leads to cancer stem cell-like properties in tamoxifen-resistant breast cancer cells through induction of interleukin-6. J. Biol. Chem. 2017, 292, 8136–8148. [Google Scholar] [CrossRef]
- Cai, J.; Tian, A.X.; Wang, Q.S.; Kong, P.Z.; Du, X.; Li, X.Q.; Feng, Y.M. FOXF2 suppresses the FOXC2-mediated epithelial-mesenchymal transition and multidrug resistance of basal-like breast cancer. Cancer Lett. 2015, 367, 129–137. [Google Scholar] [CrossRef]
- Xie, T.; Geng, J.; Wang, Y.; Wang, L.; Huang, M.; Chen, J.; Zhang, K.; Xue, L.; Liu, X.; Mao, X.; et al. FOXM1 evokes 5-fluorouracil resistance in colorectal cancer depending on ABCC10. Oncotarget 2017, 8, 8574–8589. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Zhu, Q.; Li, Z.; Peng, Y.; Yu, X.; Yuan, B.; Liu, Y.; Liu, Y.; Yin, L.; Peng, Y.; et al. The FOXM1-ABCC5 axis contributes to paclitaxel resistance in nasopharyngeal carcinoma cells. Cell Death Dis. 2017, 8, e2659. [Google Scholar] [CrossRef]
- Monteiro, L.J.; Khongkow, P.; Kongsema, M.; Morris, J.R.; Man, C.; Weekes, D.; Koo, C.Y.; Gomes, A.R.; Pinto, P.H.; Varghese, V.; et al. The Forkhead Box M1 protein regulates BRIP1 expression and DNA damage repair in epirubicin treatment. Oncogene 2013, 32, 4634–4645. [Google Scholar] [CrossRef]
- Kwok, J.M.; Peck, B.; Monteiro, L.J.; Schwenen, H.D.; Millour, J.; Coombes, R.C.; Myatt, S.S.; Lam, E.W. FOXM1 confers acquired cisplatin resistance in breast cancer cells. Mol. Cancer Res. 2010, 8, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.C.; Kashida, Y.; Kulp, S.K.; Wang, D.; Brueggemeier, R.W.; Shapiro, C.L.; Chen, C.S. Sensitizing estrogen receptor-negative breast cancer cells to tamoxifen with OSU-03012, a novel celecoxib-derived phosphoinositide-dependent protein kinase-1/Akt signaling inhibitor. Mol. Cancer Ther. 2008, 7, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Nestal de Moraes, G.; Bella, L.; Zona, S.; Burton, M.J.; Lam, E.W. Insights into a critical role of the FOXO3a-FOXM1 axis in DNA damage response and genotoxic drug resistance. Curr. Drug Targets 2016, 17, 164–177. [Google Scholar] [CrossRef]
- Bullock, M. FOXO factors and breast cancer: Outfoxing endocrine resistance. Endocr. Relat. Cancer 2016, 23, R113–R130. [Google Scholar] [CrossRef] [PubMed]
- Weng, W.; Okugawa, Y.; Toden, S.; Toiyama, Y.; Kusunoki, M.; Goel, A. FOXM1 and FOXQ1 are promising prognostic biomarkers and novel targets of tumor-suppressive miR-342 in human colorectal cancer. Clin. Cancer Res. 2016, 22, 4947–4957. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yu, X.; Bai, Q. miR-204 inhibits invasion and epithelial-mesenchymal transition by targeting FOXM1 in esophageal cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 12775–12783. [Google Scholar]
- Song, G.Q.; Zhao, Y. MicroRNA-211, a direct negative regulator of CDC25B expression, inhibits triple-negative breast cancer cells’ growth and migration. Tumour Biol. 2015, 36, 5001–5009. [Google Scholar] [CrossRef]
- Yang, X.W.; Shen, G.Z.; Cao, L.Q.; Jiang, X.F.; Peng, H.P.; Shen, G.; Chen, D.; Xue, P. MicroRNA-1269 promotes proliferation in human hepatocellular carcinoma via downregulation of FOXO1. BMC Cancer 2014, 14, 909. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Yang, T.; Yuan, S.; Wang, R.; Pan, Z.; Yang, Y.; Huang, G.; Gu, F.; Jiang, B.; et al. Double-negative feedback loop between microRNA-422a and forkhead box [FOX]G1/Q1/E1 regulates hepatocellular carcinoma tumor growth and metastasis. Hepatology 2015, 61, 561–573. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, Z.; Qiu, N.; Ling, L.; Jia, X.; Song, Y.; Li, H.; Li, J.; Lyu, H.; Liu, H.; et al. Disruption of FOXO3a-miRNA feedback inhibition of IGF2/IGF-1R/IRS1 signaling confers Herceptin resistance in HER2-positive breast cancer. Nat. Commun. 2021, 12, 2699. [Google Scholar] [CrossRef]
- Huang, X.; Gao, L.; Wang, S.; McManaman, J.L.; Thor, A.D.; Yang, X.; Esteva, F.J.; Liu, B. Heterotrimerization of the growth factor receptors erbB2, erbB3, and insulin-like growth factor-i receptor in breast cancer cells resistant to herceptin. Cancer Res. 2010, 70, 1204–1214. [Google Scholar] [CrossRef]
- Guertin, D.A.; Stevens, D.M.; Thoreen, C.C.; Burds, A.A.; Kalaany, N.Y.; Moffat, J.; Brown, M.; Fitzgerald, K.J.; Sabatini, D.M. Ablation in mice of the mTORC components raptor, rictor, or mLST8 reveals that mTORC2 is required for signalling to Akt-FOXO and PKCalpha, but not S6K1. Dev. Cell 2006, 11, 859–871. [Google Scholar]
- Ding, M.; Bruick, R.K.; Yu, Y. Secreted IGFBP5 mediates mTORC1-dependent feedback inhibition of IGF-1 signalling. Nat. Cell Biol. 2016, 18, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Hamurcu, Z.; Sener, E.F.; Taheri, S.; Nalbantoglu, U.; Kokcu, N.D.; Tahtasakal, R.; Cınar, V.; Guler, A.; Ozkul, Y.; Dönmez-Altuntas, H.; et al. MicroRNA profiling identifies Forkhead box transcription factor M1 [FOXM1] regulated miR-186 and miR-200b alterations in triple negative breast cancer. Cell. Signal. 2021, 83, 109979. [Google Scholar] [CrossRef] [PubMed]
- Hamurcu, Z.; Ashour, A.; Kahraman, N.; Ozpolat, B. FOXM1 regulates expression of eukaryotic elongation factor 2 kinase [EF2K] and promotes proliferation, invasion and tumorgenesis of human triple negative breast cancer cells. Oncotarget 2016, 7, 16619–16635. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, J.; Liu, Y.; Zhang, Z.; Duan, S. MicroRNA-937 inhibits cell proliferation and metastasis in gastric cancer cells by downregulating FOXL2. Cancer Biomark. 2017, 21, 105–116. [Google Scholar] [CrossRef]
- Zhang, L.; Zeng, D.; Chen, Y.; Li, N.; Lv, Y.; Li, Y.; Xu, X.; Xu, G. miR-937 contributes to the lung cancer cell proliferation by targeting INPP4B. Life Sci. 2016, 155, 110–115. [Google Scholar] [CrossRef]
- Han, X.; Guo, X.; Zhang, W.; Cong, Q. MicroRNA-937 inhibits the malignant phenotypes of breast cancer by directly targeting and downregulating forkhead box Q1. Oncol. Targets Ther. 2019, 12, 4813–4824, Retraction in Oncol. Targets Ther. 2021, 14, 4991–4992. [Google Scholar] [CrossRef]
- Wang, J.; Li, W.; Zhao, Y.; Kang, D.; Fu, W.; Zheng, X.; Pang, X.; Du, G. Members of FOX family could be drug targets of cancers. Pharmacol. Ther. 2018, 181, 183–196. [Google Scholar] [CrossRef]
- Li, Z.; Gadue, P.; Chen, K.; Jiao, Y.; Tuteja, G.; Schug, J.; Li, W.; Kaestner, K.H. Foxa2 and H2A.Z mediate nucleosome depletion during embryonic stem cell differentiation. Cell 2012, 151, 1608–1616. [Google Scholar] [CrossRef]
- Omatsu, Y.; Seike, M.; Sugiyama, T.; Kume, T.; Nagasawa, T. Foxc1 is a critical regulator of haematopoietic stem/progenitor cell niche formation. Nature 2014, 508, 536–540. [Google Scholar] [CrossRef]
- Lay, K.; Kume, T.; Fuchs, E. FOXC1 maintains the hair follicle stem cell niche and governs stem cell quiescence to preserve long-term tissue-regenerating potential. Proc. Natl. Acad. Sci. USA 2016, 113, E1506–E1515. [Google Scholar] [CrossRef]
- Lambers, E.; Arnone, B.; Fatima, A.; Qin, G.; Wasserstrom, J.A.; Kume, T. Foxc1 Regulates Early Cardiomyogenesis and Functional Properties of Embryonic Stem Cell Derived Cardiomyocytes. Stem Cells 2016, 34, 1487–1500. [Google Scholar] [CrossRef]
- Paranjape, A.N.; Soundararajan, R.; Werden, S.J.; Joseph, R.; Taube, J.H.; Liu, H.; Rodriguez-Canales, J.; Sphyris, N.; Wistuba, I.; Miura, N.; et al. Inhibition of FOXC2 restores epithelial phenotype and drug sensitivity in prostate cancer cells with stem-cell properties. Oncogene 2016, 35, 5963–5976. [Google Scholar] [CrossRef]
- Hollier, B.G.; Tinnirello, A.A.; Werden, S.J.; Evans, K.W.; Taube, J.H.; Sarkar, T.R.; Sphyris, N.; Shariati, M.; Kumar, S.V.; Battula, V.L.; et al. FOXC2 expression links epithelial-mesenchymal transition and stem cell properties in breast cancer. Cancer Res. 2013, 73, 1981–1992. [Google Scholar] [CrossRef]
- Liu, K.; Sun, Y.; Liu, D.; Ye, S. Inhibition of Wnt/beta-catenin signaling by IWR1 induces expression of Foxd3 to promote mouse epiblast stem cell self-renewal. Biochem. Biophys. Res. Commun. 2017, 490, 616–622. [Google Scholar] [CrossRef]
- Wei, H.J.; Nickoloff, J.A.; Chen, W.H.; Liu, H.Y.; Lo, W.C.; Chang, Y.T.; Yang, P.C.; Wu, C.W.; Williams, D.F.; Gelovani, J.G.; et al. FOXF1 mediates mesenchymal stem cell fusion-induced reprogramming of lung cancer cells. Oncotarget 2014, 5, 9514–9529. [Google Scholar] [CrossRef]
- Nik, A.M.; Reyahi, A.; Ponten, F.; Carlsson, P. Foxf2 in intestinal fibroblasts reduces numbers of Lgr5[+] stem cells and adenoma formation by inhibiting Wnt signaling. Gastroenterology 2013, 144, 1001–1011. [Google Scholar] [CrossRef]
- Bao, B.; Wang, Z.; Ali, S.; Kong, D.; Banerjee, S.; Ahmad, A.; Li, Y.; Azmi, A.S.; Miele, L.; Sarkar, F.H. Over-expression of FoxM1 leads to epithelial-mesenchymal transition and cancer stem cell phenotype in pancreatic cancer cells. J. Cell Biochem. 2011, 112, 2296–2306. [Google Scholar] [CrossRef]
- Song, I.S.; Jeong, Y.J.; Jeong, S.H.; Heo, H.J.; Kim, H.K.; Bae, K.B.; Park, Y.H.; Kim, S.U.; Kim, J.M.; Kim, N.; et al. FOXM1-Induced PRX3 Regulates Stemness and Survival of Colon Cancer Cells via Maintenance of Mitochondrial Function. Gastroenterology 2015, 149, 1006–1016. e1009. [Google Scholar] [CrossRef]
- Jiang, L.; Wu, X.; Wang, P.; Wen, T.; Yu, C.; Wei, L.; Chen, H. Targeting FoxM1 by thiostrepton inhibits growth and induces apoptosis of laryngeal squamous cell carcinoma. J. Cancer Res. Clin. Oncol. 2015, 141, 971–981. [Google Scholar] [CrossRef]
- Radhakrishnan, S.K.; Bhat, U.G.; Hughes, D.E.; Wang, I.C.; Costa, R.H.; Gartel, A.L. Identification of a chemical inhibitor of the oncogenic transcription factor forkhead box M1. Cancer Res. 2006, 66, 9731–9735. [Google Scholar] [CrossRef]
- Gusarova, G.A.; Wang, I.C.; Major, M.L.; Kalinichenko, V.V.; Ackerson, T.; Petrovic, V.; Costa, R.H. A cell-penetrating ARF peptide inhibitor of FoxM1 in mouse hepatocellular carcinoma treatment. J. Clin. Investig. 2007, 117, 99–111. [Google Scholar] [CrossRef]
- Hill, R.; Kalathur, R.K.R.; Callejas, S.; Colaço, L.; Brandão, R.; Serelde, B.; Cebriá, A.; Blanco-Aparicio, C.; Pastor, J.; Futschik, M.; et al. A novel phosphatidylinositol 3-kinase [PI3K] inhibitor directs a potent FOXO-dependent, p53- independent cell cycle arrest phenotype characterized by the differential induction of a subset of FOXO-regulated genes. Breast Cancer Res. 2014, 16, 482. [Google Scholar] [CrossRef]
- Gomez-Gutierrez, J.G.; Souza, V.; Hao, H.Y.; Montes de Oca-Luna, R.; Dong, Y.B.; Zhou, H.S.; McMasters, K.M. Adenovirus-mediated gene transfer of FKHRL1 triple mutant efficiently induces apoptosis in melanoma cells. Cancer Biol. Ther. 2006, 5, 875–883. [Google Scholar] [CrossRef]
- Kim, S.H.; Kaschula, C.H.; Priedigkeit, N.; Lee, A.V.; Singh, S.V. Forkhead Box Q1 Is a Novel Target of Breast Cancer Stem Cell Inhibition by Diallyl Trisulfide. J. Biol. Chem. 2016, 291, 13495–13508. [Google Scholar] [CrossRef]
- Liu, H.; Song, Y.; Qiu, H.; Liu, Y.; Luo, K.; Yi, Y.; Jiang, G.; Lu, M.; Zhang, Z.; Yin, J.; et al. Downregulation of FOXO3a by DNMT1 promotes breast cancer stem cell properties and tumorigenesis. Cell Death Differ. 2020, 27, 966–983. [Google Scholar] [CrossRef]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; Van De Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Laganiere, J.; Deblois, G.; Lefebvre, C.; Bataille, A.R.; Robert, F.; Giguere, V. From the cover: Location analysis of estrogen receptor alpha target promoters reveals that foxa1 defines a domain of the estrogen response. Proc. Natl. Acad. Sci. USA 2005, 102, 11651–11656. [Google Scholar] [CrossRef]
- Zaret, K.S.; Carroll, J.S. Pioneer transcription factors: Establishing competence for gene expression. Genes. Dev. 2011, 25, 2227–2241. [Google Scholar] [CrossRef]
- Carroll, J.S.; Liu, X.S.; Brodsky, A.S.; Li, W.; Meyer, C.A.; Szary, A.J.; Eeckhoute, J.; Shao, W.; Hestermann, E.V.; Geistlinger, T.R.; et al. Chromosome-wide mapping of estrogen receptor binding reveals long-range regulation requiring the forkhead protein FoxA1. Cell 2005, 122, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Augello, M.A.; Hickey, T.E.; Knudsen, K.E. FOXA1: Master of steroid receptor function in cancer. EMBO J. 2011, 30, 3885–3894. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.L.; Men, J.R.; Liu, H.Y.; Liu, M.Y.; Zhang, H.S. FOXM1 facilitates breast cancer cell stemness and migration in YAP1-dependent manner. Arch. Biochem. Biophys. 2020, 685, 108349. [Google Scholar] [CrossRef]
- Dilmac, S.; Kuscu, N.; Caner, A.; Yildirim, S.; Yoldas, B.; Farooqi, A.A.; Tanriover, G. SIRT1/FOXO Signaling Pathway in Breast Cancer Progression and Metastasis. Int. J. Mol. Sci. 2022, 23, 10227. [Google Scholar] [CrossRef]
- Liu, J.; Duan, Z.; Guo, W.; Zeng, L.; Wu, Y.; Chen, Y.; Tai, F.; Wang, Y.; Lin, Y.; Zhang, Q.; et al. Targeting the BRD4/FOXO3a/CDK6 axis sensitizes AKT inhibition in luminal breast cancer. Nat. Commun. 2018, 9, 5200. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Zeng, C.; Wang, S.; Han, J.; Zhu, L.; Zhao, H.; Zhang, Y.; Lu, J.; Xu, Y. Forkhead box P3 promotes breast cancer cell apoptosis by regulating programmed cell death 4 expression. Oncol. Lett. 2020, 20, 292. [Google Scholar] [CrossRef]
- Badve, S.; Nakshatri, H. Oestrogen-receptor-positive breast cancer: Towards bridging histopathological and molecular classifications. J. Clin. Pathol. 2009, 62, 6–12. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Hallmarks | FOXA | FOXC | FOXM | FOXO | FOXP |
|---|---|---|---|---|---|
| Invasion and metastasis | FOXA1, FOXA2, FOXA3 | FOXC1, FOXC2 | FOXM1 | FOXO1, FOXO3A, FOXO4 | FOXP1, FOXP3 |
| Immune destruction | X | X | X | FOXO1 | FOXP1, FOXP3, FOXP4 |
| Cellular energetics | FOXA2 | FOXC2 | FOXM1 | FOXO1, FOXO6, FOXO3A | X |
| Replicative immortality | FOXA1, FOXA2 | FOXC2 | FOXM1 | FOXO1, FOXO3A, FOXO4 | X |
| Evading growth suppressors | FOXA1, FOXA2 | X | FOXM1 | FOXO1, FOXO3A, FOXO4 | FOXP1, FOXP3 |
| Genome stability and mutation | FOXA1, FOXA2, FOXA3 | FOXC1, FOXC2 | FOXM1 | FOXO1, FOXO6, FOXO3A, FOXO4 | FOXP1, FOXP2, FOXP3, FOXP4 |
| Inducing angiogenesis | FOXA1, FOXA2 | FOXC1, FOXC2 | FOXM1 | FOXO1, FOXO3A | FOXP1, FOXP3 |
| Resisting cell death | FOXA1, FOXA2, FOXA3 | X | FOXM1 | FOXO1, FOXO6, FOXO3A, FOXO4 | FOXP1, FOXP3 |
| Sustaining proliferative signaling | FOXA1, FOXA2 | FOXC1, FOXC2 | FOXM1 | FOXO1, FOXO6, FOXO3A, FOXO4 | FOXP1, FOXP3 |
| Tumor proliferative inflammation | FOXA1, FOXA3 | FOXC1 | FOXM1 | FOXO1, FOXO6, FOXO3A, FOXO4 | FOXP3 |
| miRNA Class | Interaction with Forkhead Box | Associated Cancer | References |
|---|---|---|---|
| miR-342 | FOXM1, FOXQ1 | Colorectal cancer | [87] |
| miR-204 | FOXM1 | Esophageal cancer | [88] |
| miR-422a | FOXQ1, FOXG1, FOXE1 | Liver cancer | [91] |
| miRNAs | FOXO3A | HER2 Positive breast cancer | [92,93] |
| miR200b-5p | FOXM1 | Breast cancer | [97] |
| miR-186-5p | FOXM1 | Breast cancer | [97] |
| miRNA-937 | FOXQ1 | Stomach, lung, breast cancer | [98,99,100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadaf; Hazazi, A.; Alkhalil, S.S.; Alsaiari, A.A.; Gharib, A.F.; Alhuthali, H.M.; Rana, S.; Aloliqi, A.A.; Eisa, A.A.; Hasan, M.R.; et al. Role of Fork-Head Box Genes in Breast Cancer: From Drug Resistance to Therapeutic Targets. Biomedicines 2023, 11, 2159. https://doi.org/10.3390/biomedicines11082159
Sadaf, Hazazi A, Alkhalil SS, Alsaiari AA, Gharib AF, Alhuthali HM, Rana S, Aloliqi AA, Eisa AA, Hasan MR, et al. Role of Fork-Head Box Genes in Breast Cancer: From Drug Resistance to Therapeutic Targets. Biomedicines. 2023; 11(8):2159. https://doi.org/10.3390/biomedicines11082159
Chicago/Turabian StyleSadaf, Ali Hazazi, Samia S. Alkhalil, Ahad Amer Alsaiari, Amal F. Gharib, Hayaa M. Alhuthali, Shanika Rana, Abdulaziz A. Aloliqi, Alaa Abdulaziz Eisa, Mohammad Raghibul Hasan, and et al. 2023. "Role of Fork-Head Box Genes in Breast Cancer: From Drug Resistance to Therapeutic Targets" Biomedicines 11, no. 8: 2159. https://doi.org/10.3390/biomedicines11082159
APA StyleSadaf, Hazazi, A., Alkhalil, S. S., Alsaiari, A. A., Gharib, A. F., Alhuthali, H. M., Rana, S., Aloliqi, A. A., Eisa, A. A., Hasan, M. R., & Dev, K. (2023). Role of Fork-Head Box Genes in Breast Cancer: From Drug Resistance to Therapeutic Targets. Biomedicines, 11(8), 2159. https://doi.org/10.3390/biomedicines11082159