

A2AR Expression and Immunosuppressive Environment Independent of KRAS and GNAS Mutations in Pseudomyxoma Peritonei
 ,
,  , ,
, ,  , ,
, , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Patients
2.2. NGS, Immunohistochemistry, and Transcriptional Analysis
2.3. Prognosis and Statistical Analysis
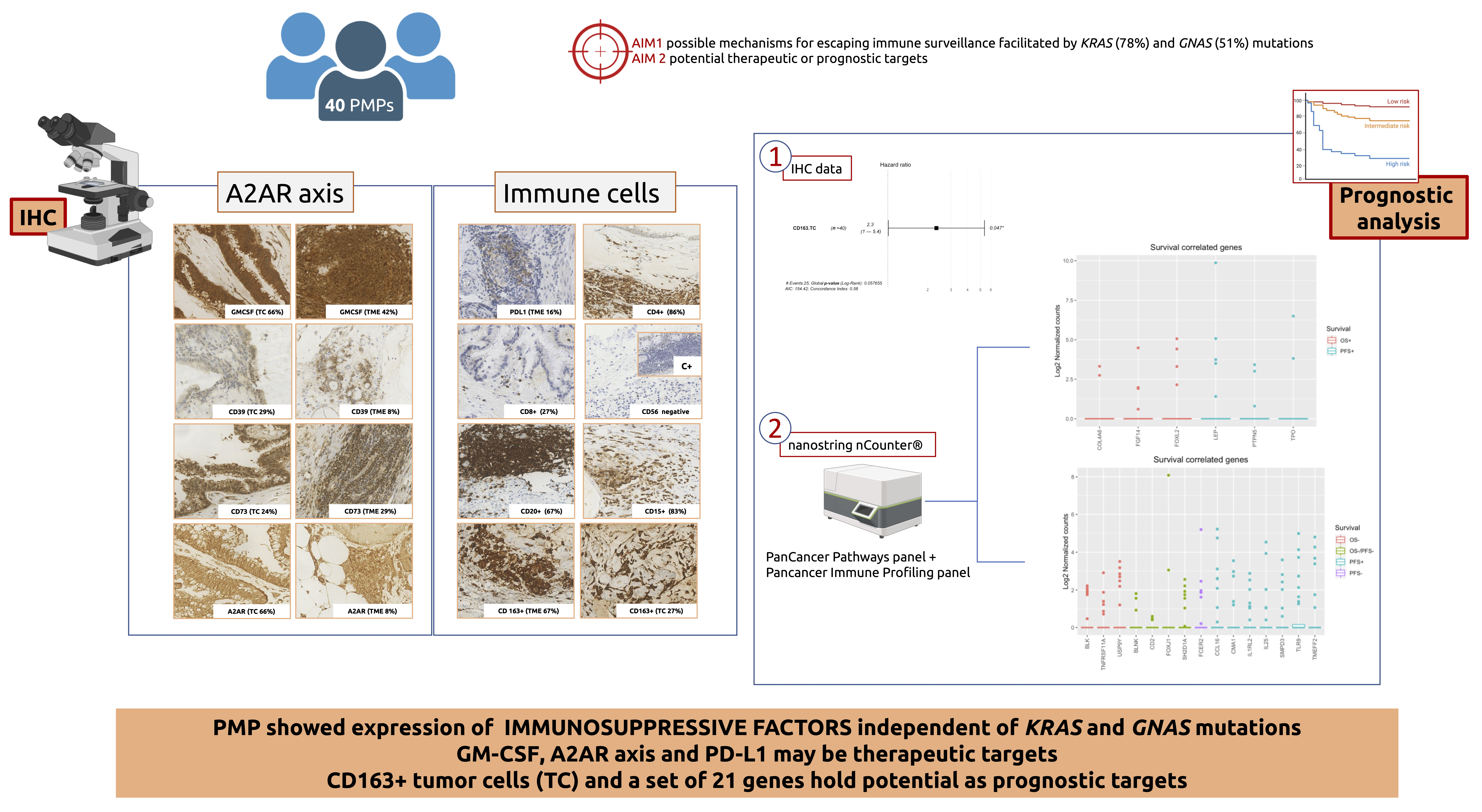
3. Results
3.1. KRAS and GNAS Status
3.2. Immunohistochemical Analysis
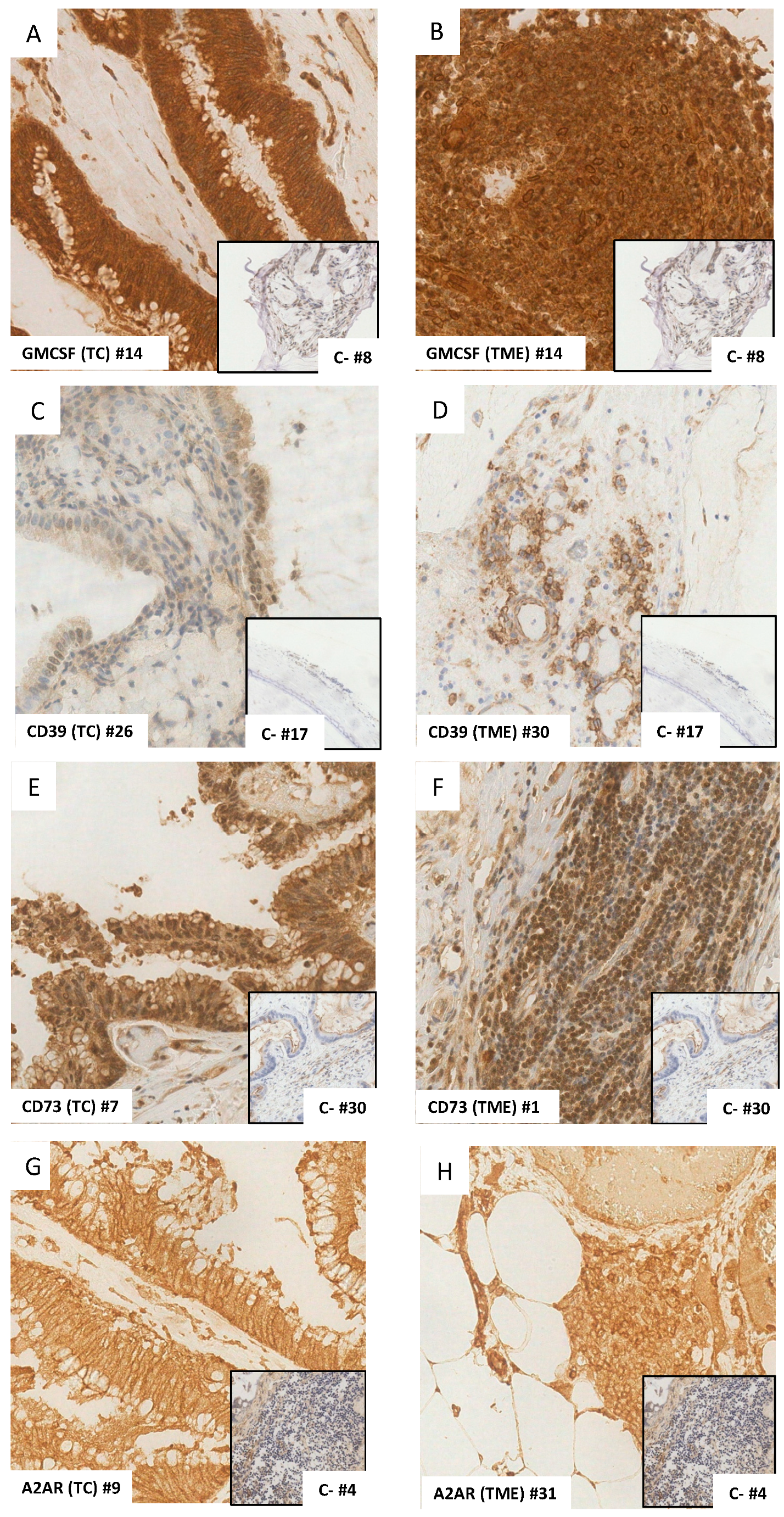
3.2.1. GM-CSF
3.2.2. CD39
3.2.3. CD73
3.2.4. A2AR
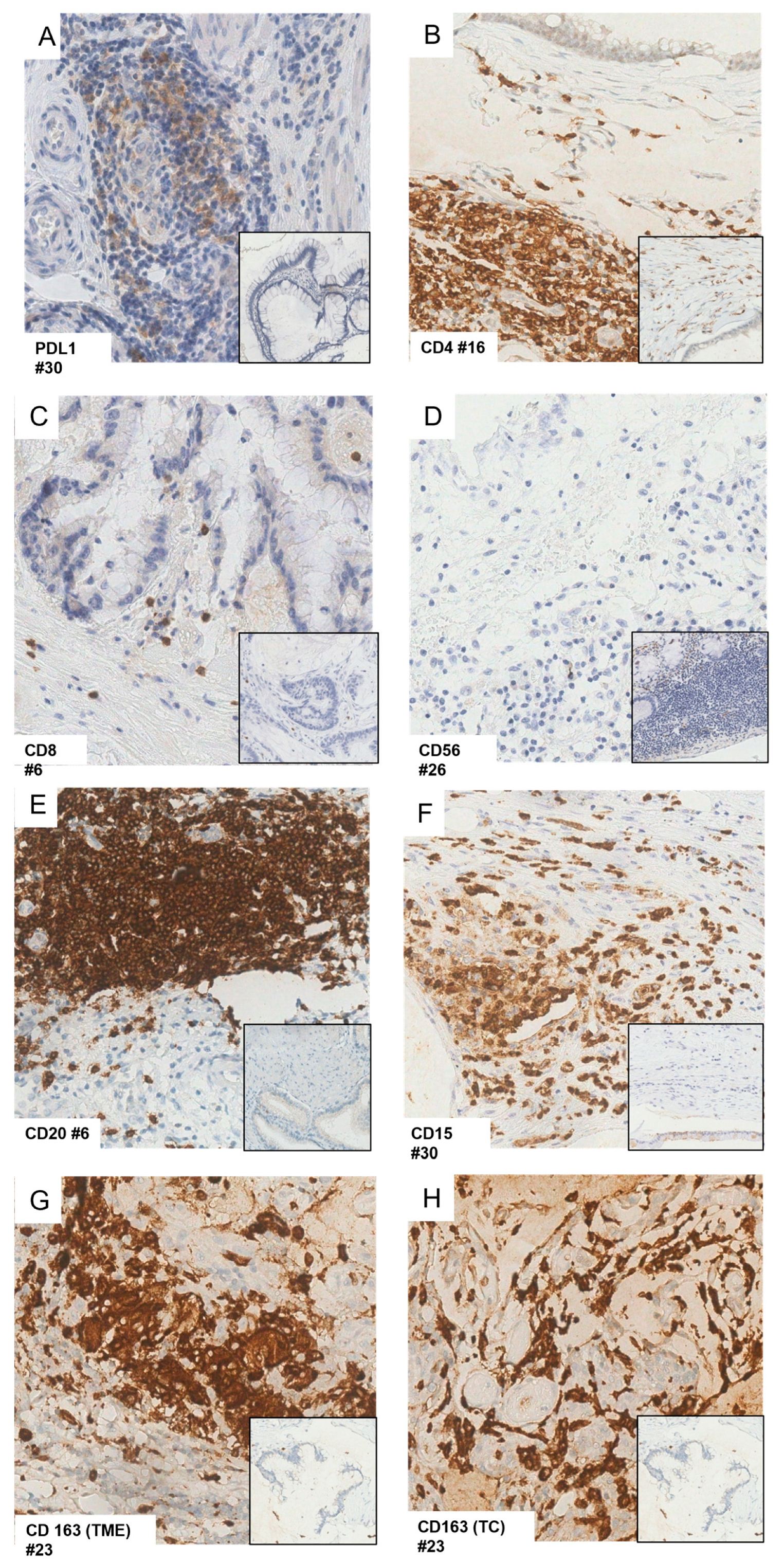
3.2.5. PD-L1
3.2.6. T Cells
3.2.7. Natural Killer (NK) Cells
3.2.8. B Cells
3.2.9. Granulocytic Myeloid-Derived Suppressor Cells
3.2.10. M2 Macrophages
3.3. Transcriptional Analysis
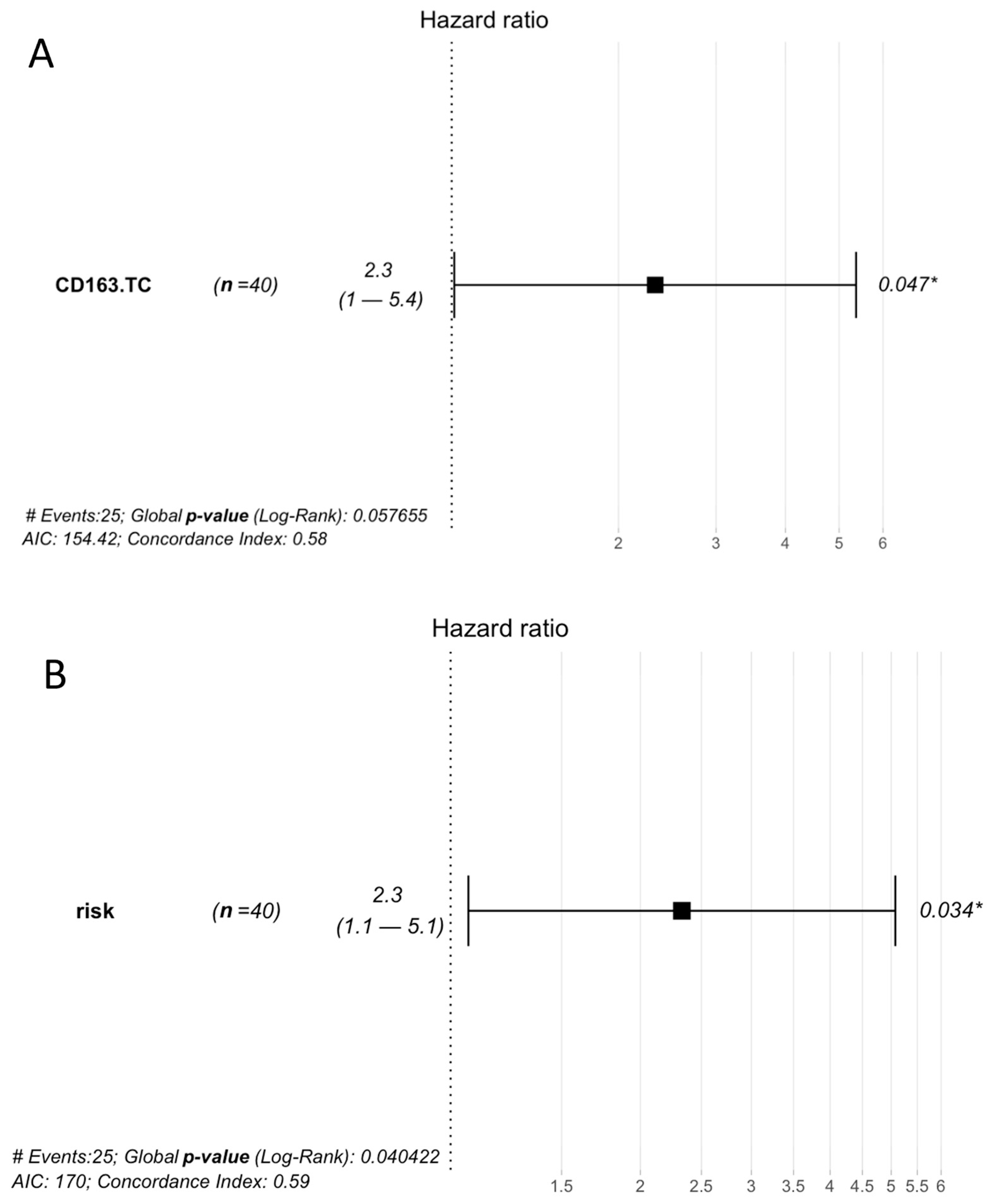
3.4. Prognostic Analysis
3.5. Risk Classification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Moran, B.J.; Cecil, T.D. The etiology, clinical presentation, and management of pseudomyxoma peritonei. Surg. Oncol. Clin. N. Am. 2003, 12, 585–603. [Google Scholar] [CrossRef]
- Carr, N.J.; McCarthy, W.F.; Sobin, L.H. Epithelial noncarcinoid tumors and tumor-like lesions of the appendix. A clinicopathologic study of 184 patients with a multivariate analysis of prognostic factors. Cancer 1995, 75, 757–768. [Google Scholar] [CrossRef]
- Sugarbaker, P.H. New standard of care for appendiceal epithelial neoplasms and pseudomyxoma peritonei syndrome? Lancet Oncol. 2006, 7, 69–76. [Google Scholar] [CrossRef]
- Baratti, D.; Milito, P.; Kusamura, S.; Martin Roman, L.; Guaglio, M.; Deraco, M. Systemic metastases from low-grade and high-grade pseudomyxoma peritonei: Treatments and outcomes. Eur. J. Surg. Oncol. 2022, 48, 1590–1597. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, E.M.; Feldman, R.; Mapow, B.L.; Mackovick, L.T.; Ward, K.M.; Morano, W.F.; Rubin, R.R.; Bowne, W.B. Appendix-derived Pseudomyxoma Peritonei (PMP): Molecular Profiling Toward Treatment of a Rare Malignancy. Am. J. Clin. Oncol. 2018, 41, 777–783. [Google Scholar] [CrossRef]
- Pengelly, R.J.; Rowaiye, B.; Pickard, K.; Moran, B.; Dayal, S.; Tapper, W.; Mirnezami, A.; Cecil, T.; Mohamed, F.; Carr, N.; et al. Analysis of Mutation and Loss of Heterozygosity by Whole-Exome Sequencing Yields Insights into Pseudomyxoma Peritonei. J. Mol. Diagn. 2018, 20, 635–642. [Google Scholar] [CrossRef]
- Pietrantonio, F.; Berenato, R.; Maggi, C.; Caporale, M.; Milione, M.; Perrone, F.; Tamborini, E.; Baratti, D.; Kusamura, S.; Mariani, L.; et al. GNAS mutations as prognostic biomarker in patients with relapsed peritoneal pseudomyxoma receiving metronomic capecitabine and bevacizumab: A clinical and translational study. J. Transl. Med. 2016, 14, 125. [Google Scholar] [CrossRef]
- Pietrantonio, F.; Perrone, F.; Mennitto, A.; Gleeson, E.M.; Milione, M.; Tamborini, E.; Busico, A.; Settanni, G.; Berenato, R.; Caporale, M.; et al. Toward the molecular dissection of peritoneal pseudomyxoma. Ann. Oncol. 2016, 27, 2097–2103. [Google Scholar] [CrossRef]
- Kuracha, M.R.; Thomas, P.; Loggie, B.W.; Govindarajan, V. Bilateral blockade of MEK- and PI3K-mediated pathways downstream of mutant KRAS as a treatment approach for peritoneal mucinous malignancies. PLoS ONE 2017, 12, e0179510. [Google Scholar] [CrossRef] [PubMed]
- Dilly, A.K.; Honick, B.D.; Lee, Y.J.; Guo, Z.S.; Zeh, H.J.; Bartlett, D.L.; Choudry, H.A. Targeting G-protein coupled receptor-related signaling pathway in a murine xenograft model of appendiceal pseudomyxoma peritonei. Oncotarget 2017, 8, 106888–106900. [Google Scholar] [CrossRef] [PubMed]
- Petanidis, S.; Anestakis, D.; Argyraki, M.; Hadzopoulou-Cladaras, M.; Salifoglou, A. Differential expression of IL-17, 22 and 23 in the progression of colorectal cancer in patients with K-ras mutation: Ras signal inhibition and crosstalk with GM-CSF and IFN-gamma. PLoS ONE 2013, 8, e73616. [Google Scholar] [CrossRef]
- Pylayeva-Gupta, Y.; Lee, K.E.; Hajdu, C.H.; Miller, G.; Bar-Sagi, D. Oncogenic Kras-induced GM-CSF production promotes the development of pancreatic neoplasia. Cancer Cell 2012, 21, 836–847. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Schmetterer, K.G.; Goldhahn, K.; Ziegler, L.S.; Gerner, M.C.; Schmidt, R.L.J.; Themanns, M.; Zebedin-Brandl, E.; Trapin, D.; Leitner, J.; Pickl, W.F.; et al. Overexpression of PDE4A Acts as Checkpoint Inhibitor Against cAMP-Mediated Immunosuppression in vitro. Front. Immunol. 2019, 10, 1790. [Google Scholar] [CrossRef] [PubMed]
- Antonioli, L.; Blandizzi, C.; Pacher, P.; Hasko, G. Immunity, inflammation and cancer: A leading role for adenosine. Nat. Rev. Cancer 2013, 13, 842–857. [Google Scholar] [CrossRef] [PubMed]
- Bullen, J.W.; Tchernyshyov, I.; Holewinski, R.J.; DeVine, L.; Wu, F.; Venkatraman, V.; Kass, D.L.; Cole, R.N.; Van Eyk, J.; Semenza, G.L. Protein kinase A-dependent phosphorylation stimulates the transcriptional activity of hypoxia-inducible factor 1. Sci. Signal 2016, 9, ra56. [Google Scholar] [CrossRef]
- Bruzzese, L.; Fromonot, J.; By, Y.; Durand-Gorde, J.M.; Condo, J.; Kipson, N.; Guieu, R.; Fenouillet, E.; Ruf, J. NF-kappaB enhances hypoxia-driven T-cell immunosuppression via upregulation of adenosine A(2A) receptors. Cell. Signal. 2014, 26, 1060–1067. [Google Scholar] [CrossRef]
- Cekic, C.; Linden, J. Purinergic regulation of the immune system. Nat. Rev. Immunol. 2016, 16, 177–192. [Google Scholar] [CrossRef]
- Ohta, A.; Gorelik, E.; Prasad, S.J.; Ronchese, F.; Lukashev, D.; Wong, M.K.; Huang, X.; Caldwell, S.; Liu, K.; Smith, P.; et al. A2A adenosine receptor protects tumors from antitumor T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13132–13137. [Google Scholar] [CrossRef]
- Vijayan, D.; Young, A.; Teng, M.W.L.; Smyth, M.J. Targeting immunosuppressive adenosine in cancer. Nat. Rev. Cancer 2017, 17, 709–724. [Google Scholar] [CrossRef]
- Carr, N.J.; Bibeau, F.; Bradley, R.F.; Dartigues, P.; Feakins, R.M.; Geisinger, K.R.; Gui, X.; Isaac, S.; Milione, M.; Misdraji, J.; et al. The histopathological classification, diagnosis and differential diagnosis of mucinous appendiceal neoplasms, appendiceal adenocarcinomas and pseudomyxoma peritonei. Histopathology 2017, 71, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Milione, M.; Miceli, R.; Barretta, F.; Pellegrinelli, A.; Spaggiari, P.; Tagliabue, G.; Centonze, G.; Paolino, C.; Mangogna, A.; Kankava, K.; et al. Microenvironment and tumor inflammatory features improve prognostic prediction in gastro-entero-pancreatic neuroendocrine neoplasms. J. Pathol. Clin. Res. 2019, 5, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Su, L.; Yang, W.; Zhang, J.; Wang, Y.; Wang, C.; Yu, Y.; Yang, L.; Zhou, Z. Development and validation of prognostic nomograms for pseudomyxoma peritonei patients after surgery: A population-based study. Medicine 2020, 99, e20963. [Google Scholar] [CrossRef]
- Yan, W.L.; Shen, K.Y.; Tien, C.Y.; Chen, Y.A.; Liu, S.J. Recent progress in GM-CSF-based cancer immunotherapy. Immunotherapy 2017, 9, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Graef, S.; Mussai, F.; Thomas, A.; Wali, N.; Yenidunya, B.G.; Yuan, C.; Morrow, B.; Zhang, J.; Korangy, F.; et al. Tumor-Derived GM-CSF Promotes Granulocyte Immunosuppression in Mesothelioma Patients. Clin. Cancer Res. 2018, 24, 2859–2872. [Google Scholar] [CrossRef]
- Sek, K.; Molck, C.; Stewart, G.D.; Kats, L.; Darcy, P.K.; Beavis, P.A. Targeting Adenosine Receptor Signaling in Cancer Immunotherapy. Int. J. Mol. Sci. 2018, 19, 3837. [Google Scholar] [CrossRef]
- Hatfield, S.; Veszeleiova, K.; Steingold, J.; Sethuraman, J.; Sitkovsky, M. Mechanistic Justifications of Systemic Therapeutic Oxygenation of Tumors to Weaken the Hypoxia Inducible Factor 1alpha-Mediated Immunosuppression. Adv. Exp. Med. Biol. 2019, 1136, 113–121. [Google Scholar] [CrossRef]
- Hatfield, S.M.; Sitkovsky, M. A2A adenosine receptor antagonists to weaken the hypoxia-HIF-1alpha driven immunosuppression and improve immunotherapies of cancer. Curr. Opin. Pharmacol. 2016, 29, 90–96. [Google Scholar] [CrossRef]
- Augustin, R.C.; Leone, R.D.; Naing, A.; Fong, L.; Bao, R.; Luke, J.J. Next steps for clinical translation of adenosine pathway inhibition in cancer immunotherapy. J. Immunother. Cancer 2022, 10, e004089. [Google Scholar] [CrossRef]
- Paijens, S.T.; Vledder, A.; de Bruyn, M.; Nijman, H.W. Tumor-infiltrating lymphocytes in the immunotherapy era. Cell Mol. Immunol. 2021, 18, 842–859. [Google Scholar] [CrossRef]
- de Visser, K.E.; Korets, L.V.; Coussens, L.M. De novo carcinogenesis promoted by chronic inflammation is B lymphocyte dependent. Cancer Cell 2005, 7, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.E.; Spata, M.; Bayne, L.J.; Buza, E.L.; Durham, A.C.; Allman, D.; Vonderheide, R.H.; Simon, M.C. Hif1a Deletion Reveals Pro-Neoplastic Function of B Cells in Pancreatic Neoplasia. Cancer Discov. 2016, 6, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Sun, Q.; Wang, J.; Pan, W.; Ren, X. Positive and negative functions of B lymphocytes in tumors. Oncotarget 2016, 7, 55828–55839. [Google Scholar] [CrossRef] [PubMed]
- Garvin, S.; Oda, H.; Arnesson, L.G.; Lindstrom, A.; Shabo, I. Tumor cell expression of CD163 is associated to postoperative radiotherapy and poor prognosis in patients with breast cancer treated with breast-conserving surgery. J. Cancer Res. Clin. Oncol. 2018, 144, 1253–1263. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | KRAS | GNAS | GM-CSF TC | GM-CSF TME | CD39 TC | CD39 TME | CD73 TC | CD73 TME | A2AR TC | A2AR TME | CD4 TME | CD8 TME | CD20 TME | CD56 TME | CD15 TME | CD163 TME | CD163TC | PDL1 TME |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | M | WT | 3 | 2 | 0 | 0 | 0 | 4 | 6 | 0 | 9 | 0 | 2 | 0 | 0 | 2 | 9 | 0 |
| 2 | M | M | 12 | 0 | 4 | 0 | 3 | 0 | 4 | 0 | 4 | 0 | 2 | 0 | 6 | 0 | 0 | 0 |
| 3 | WT | WT | 3 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 6 | 0 | 2 | 0 | 6 | 6 | 0 | 0 |
| 4 | WT | WT | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 4 | 0 | 2 | 0 | 3 | 2 | 0 | 0 |
| 5 | M | WT | 0 | 2 | 0 | 0 | 0 | 0 | 6 | 0 | 9 | 0 | 9 | 0 | 0 | 2 | 6 | 0 |
| 6 | M | WT | 9 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 4 | 4 | 6 | 0 | 3 | 4 | 0 | 0 |
| 7 | WT | WT | 9 | 2 | 3 | 0 | 6 | 0 | 9 | 0 | NE | NE | NE | NE | NE | NE | NE | NE |
| 8 | M | WT | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 4 | 0 | 2 | 0 | 6 | 4 | 0 | 0 |
| 9 | WT | M | 9 | 2 | 0 | 0 | 6 | 0 | 9 | 0 | 4 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| 10 | NE | NE | 2 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 2 | 0 | 2 | 0 | 2 | 6 | 0 | 0 |
| 11 | M | M | 4 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 4 | 0 | 4 | 0 | 0 | 2 | 0 | 0 |
| 12 | M | M | 6 | 4 | 0 | 0 | 0 | 0 | 9 | 0 | 6 | 2 | 9 | 0 | 4 | 6 | 6 | 0 |
| 13 | M | M | 12 | 2 | 0 | 0 | 0 | 4 | 12 | 0 | 6 | 0 | 2 | 0 | 3 | 0 | 0 | 0 |
| 14 | M | WT | 9 | 4 | 0 | 0 | 9 | 0 | 2 | 0 | 4 | 0 | 4 | 0 | 2 | 4 | 0 | 0 |
| 15 | M | M | 0 | 4 | 0 | 0 | 0 | 0 | 4 | 0 | 3 | 2 | 12 | 0 | 9 | 2 | 3 | 0 |
| 16 | M | M | 9 | 4 | 4 | 0 | 0 | 0 | 12 | 0 | 6 | 3 | 6 | 0 | 12 | 3 | 12 | 0 |
| 17 | M | M | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 4 | 0 | 0 | 0 | 2 | 4 | 0 | 0 |
| 18 | M | M | 6 | 0 | 0 | 2 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 |
| 19 | WT | WT | 6 | 4 | 4 | 0 | 0 | 0 | 9 | 0 | 9 | 0 | 6 | 0 | 6 | 4 | 0 | 0 |
| 20 | M | WT | 9 | 2 | 9 | 0 | 0 | 4 | 9 | 0 | 0 | 4 | 0 | 0 | 6 | 0 | 0 | POS |
| 21 | NE | NE | 0 | 4 | 0 | 0 | 0 | 2 | 0 | 3 | 4 | 0 | 0 | 0 | 6 | 4 | 0 | POS |
| 22 | M | WT | 4 | 0 | 2 | 0 | 0 | 0 | 4 | 0 | 4 | 2 | 4 | 0 | 0 | 0 | 0 | 0 |
| 23 | M | M | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 6 | 4 | 0 | 2 | 4 | 9 | 0 |
| 24 | M | M | 9 | 2 | 4 | 0 | 0 | 4 | 6 | 0 | 6 | 2 | 4 | 0 | 4 | 0 | 12 | POS |
| 25 | M | M | 4 | 9 | 0 | 0 | 0 | 0 | 6 | 0 | 4 | 0 | 0 | 0 | 6 | 2 | 0 | POS |
| 26 | M | M | 9 | 0 | 6 | 0 | 0 | 0 | 9 | 0 | 6 | 0 | 4 | 0 | 0 | 4 | 12 | 0 |
| 27 | M | M | 6 | 2 | 0 | 0 | 0 | 2 | 9 | 0 | 12 | 3 | 4 | 0 | 4 | 4 | 12 | POS |
| 28 | M | WT | 12 | 2 | 9 | 0 | 0 | 0 | 9 | 0 | 4 | 0 | 0 | 0 | 6 | 0 | 0 | 0 |
| 29 | M | M | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 4 | 0 | 2 | 0 | 9 | 3 | 0 | 0 |
| 30 | M | WT | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 4 | 9 | 0 | 4 | 0 | 6 | 0 | 4 | POS |
| 31 | M | M | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 0 | 0 | 0 | 6 | 4 | 0 | 0 |
| 32 | M | M | 4 | 0 | 9 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 6 | 4 | 0 | 0 |
| 33 | M | WT | 4 | 0 | 9 | 0 | 0 | 0 | 6 | 0 | 3 | 0 | 3 | 0 | 6 | 2 | 0 | 0 |
| 34 | WT | WT | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 |
| 35 | M | WT | 12 | 0 | 0 | 4 | 0 | 4 | 9 | 0 | 9 | 0 | 2 | 0 | 6 | 4 | 0 | 0 |
| 36 | M | WT | 9 | 0 | 0 | 0 | 9 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 |
| 37 | WT | M | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 6 | 0 | 4 | 0 | 2 | 4 | 0 | 0 |
| 38 | M | M | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 6 | 0 | 0 | 0 |
| 38 CASES | 29 KRAS MUT | 7 KRAS WT | 19 GNAS MUT | 17 GNAS WT | ||
|---|---|---|---|---|---|---|
| GM-CSF+ | TC | 25 (66%) | 20 (69%) | 4 (57%) | 12 (63%) | 12 (70%) |
| TME | 16 (42%) | 12 (41%) | 3 (43%) | 8 (42%) | 7 (41%) | |
| CD39+ | TC | 11 (29%) | 9 (31%) | 2 (28%) | 5 (26%) | 6 (35%) |
| TME | 3 (8%) | 3 (10%) | 0 | 1 (5%) | 2 (12%) | |
| CD73+ | TC | 9 (24%) | 6 (31%) | 2 (28%) | 5 (26%) | 3 (18%) |
| TME | 11 (29%) | 8 (27%) | 2 (28%) | 6 (31%) | 4 (23%) | |
| A2AR+ | TC | 25 (66%) | 21 (72%) | 4 (57%) | 11 (58%) | 14 (82%) |
| TME | 3 (8%) | 3 (10%) | 0 | 1 (5%) | 2 (12%) | |
| PD-L1+ | TC | 0 | 0 | 0 | 0 | 0 |
| TME | 6 (16%) | 5 (17%) | 0 | 3 (16%) | 2 (12%) |
| 37 CASES | 29 KRAS MUT | 6 KRAS WT | 19 GNAS MUT | 16 GNAS WT | 24 A2AR POS | 13 A2AR Neg | ||
|---|---|---|---|---|---|---|---|---|
| CD4+ | TME | 32 (86%) | 25 (86%) | 5 (83%) | 17 (89%) | 13 (81%) | 21 (87%) | 11 (85%) |
| CD8+ | TME | 10 (27%) | 9 (31%) | 1 (17%) | 7 (37%) | 3 (19%) | 9 (37%) | 1 (8%) |
| CD56 | TME | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| CD20+ | TME | 25 (67%) | 20 (69%) | 4 (67%) | 12 (63%) | 12 (75%) | 18 (75%) | 7 (54%) |
| CD15+ | TME | 31 (84%) | 24 (83%) | 5 (83%) | 16 (84%) | 13 (81%) | 19 (79%) | 12 (92%) |
| CD163+ | TME | 25 (67%) | 19 (65%) | 4 (67%) | 13 (68%) | 9 (56%) | 16 (67%) | 9 (69%) |
| TC | 10 (27%) | 10 (34%) | 0 | 7 (37%) | 3 (18%) | 9 (37%) | 1 (8%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kusamura, S.; Busico, A.; Conca, E.; Capone, I.; Agnelli, L.; Lorenzini, D.; Brich, S.; Angelini, M.; Volpi, C.C.; Trupia, D.V.; et al. A2AR Expression and Immunosuppressive Environment Independent of KRAS and GNAS Mutations in Pseudomyxoma Peritonei. Biomedicines 2023, 11, 2049. https://doi.org/10.3390/biomedicines11072049
Kusamura S, Busico A, Conca E, Capone I, Agnelli L, Lorenzini D, Brich S, Angelini M, Volpi CC, Trupia DV, et al. A2AR Expression and Immunosuppressive Environment Independent of KRAS and GNAS Mutations in Pseudomyxoma Peritonei. Biomedicines. 2023; 11(7):2049. https://doi.org/10.3390/biomedicines11072049
Chicago/Turabian StyleKusamura, Shigeki, Adele Busico, Elena Conca, Iolanda Capone, Luca Agnelli, Daniele Lorenzini, Silvia Brich, Marta Angelini, Chiara Costanza Volpi, Desirè Viola Trupia, and et al. 2023. "A2AR Expression and Immunosuppressive Environment Independent of KRAS and GNAS Mutations in Pseudomyxoma Peritonei" Biomedicines 11, no. 7: 2049. https://doi.org/10.3390/biomedicines11072049
APA StyleKusamura, S., Busico, A., Conca, E., Capone, I., Agnelli, L., Lorenzini, D., Brich, S., Angelini, M., Volpi, C. C., Trupia, D. V., Lagano, V., Torelli, T., Gloghini, A., Baratti, D., Guaglio, M., Milione, M., Deraco, M., & Perrone, F. (2023). A2AR Expression and Immunosuppressive Environment Independent of KRAS and GNAS Mutations in Pseudomyxoma Peritonei. Biomedicines, 11(7), 2049. https://doi.org/10.3390/biomedicines11072049