

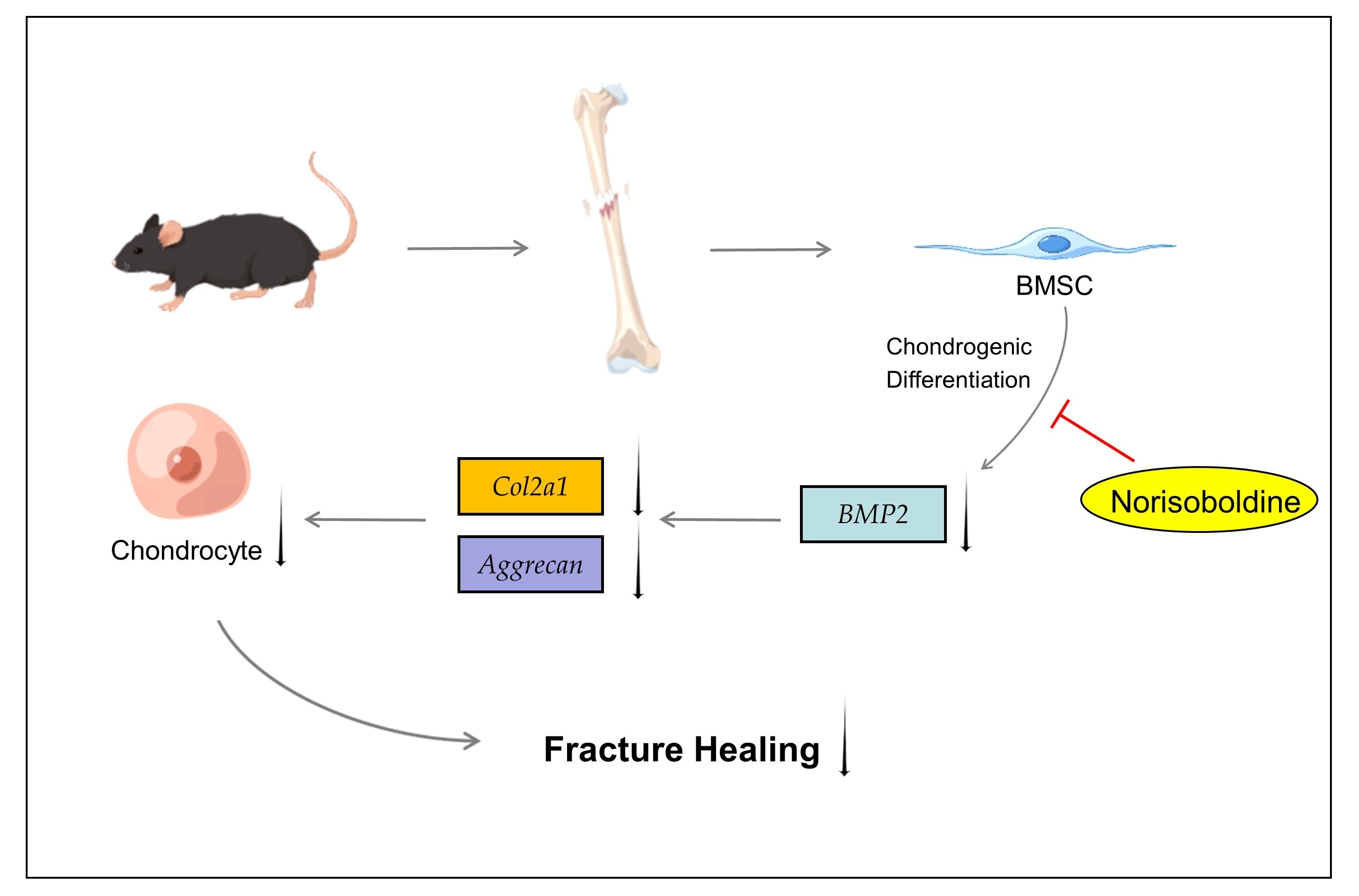
Norisoboldine, a Natural Isoquinoline Alkaloid, Inhibits Diaphyseal Fracture Healing in Mice by Alleviating Cartilage Formation

 , ,
, ,  and
and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Tibia Fracture Model
2.3. BMSC Cell Cultures
2.4. Chondrogenic Induction of BMSCs In Vitro
2.5. Micro-Computed Tomography (µCT) Analysis
2.6. Histology
2.7. Wound Healing Assay
2.8. Alcian Blue Staining
2.9. Immunofluorescent (IF) Staining
2.10. RNA Extraction and Quantitative PCR
2.11. Statistical Analysis
3. Results
3.1. NOR Inhibits Bone Regeneration by Delaying Callus Formation
3.2. NOR Decreases Cartilage Formation by Inhibiting Differentiation of BMSC to Chondrocyte
3.3. NOR Decreases Bone Ossification by Inhibiting Osterix (OSX)–Collage I (COL1) Signaling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Einhorn, T.A.; Gerstenfeld, L.C. Fracture healing: Mechanisms and interventions. Nat. Rev. Rheumatol. 2015, 11, 45–54. [Google Scholar] [CrossRef]
- Gerstenfeld, L.C.; Cho, T.J.; Kon, T.; Aizawa, T.; Tsay, A.; Fitch, J.; Barnes, G.L.; Graves, D.T.; Einhorn, T.A. Impaired fracture healing in the absence of TNF-alpha signaling: The role of TNF-alpha in endochondral cartilage resorption. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2003, 18, 1584–1592. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, Y.; Zhou, Z.; Yang, J.; Jia, Y.; Wu, H.; Dong, H.; Leng, X. Deer antler extract promotes tibia fracture healing in mice by activating BMP-2/SMAD4 signaling pathway. J. Orthop. Surg. Res. 2022, 17, 468. [Google Scholar] [CrossRef] [PubMed]
- Gerstenfeld, L.C.; Cullinane, D.M.; Barnes, G.L.; Graves, D.T.; Einhorn, T.A. Fracture healing as a post-natal developmental process: Molecular, spatial, and temporal aspects of its regulation. J. Cell. Biochem. 2003, 88, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.F.; Lv, Q.; Xia, Y.; Yue, M.F.; Shi, C.; Xia, Y.F.; Chou, G.X.; Wang, Z.T.; Dai, Y. Norisoboldine, an Anti-Arthritis Alkaloid Isolated from Radix Linderae, Attenuates Osteoclast Differentiation and Inflammatory Bone Erosion in an Aryl Hydrocarbon Receptor-Dependent Manner. Int. J. Biol. Sci. 2015, 11, 1113–1126. [Google Scholar] [CrossRef]
- Luo, Y.; Liu, M.; Xia, Y.; Dai, Y.; Chou, G.; Wang, Z. Therapeutic effect of norisoboldine, an alkaloid isolated from Radix Linderae, on collagen-induced arthritis in mice. Phytomed. Int. J. Phytother. Phytopharm. 2010, 17, 726–731. [Google Scholar] [CrossRef]
- Chen, Q.; Shao, X.; He, Y.; Lu, E.; Zhu, L.; Tang, W. Norisoboldine Attenuates Sepsis-Induced Acute Lung Injury by Modulating Macrophage Polarization via PKM2/HIF-1alpha/PGC-1alpha Pathway. Biol. Pharm. Bull. 2021, 44, 1536–1547. [Google Scholar] [CrossRef]
- Gao, S.; Li, W.; Lin, G.; Liu, G.; Deng, W.; Zhai, C.; Bian, C.; He, G.; Hu, Z. Norisoboldine, an alkaloid from Radix linderae, inhibits NFAT activation and attenuates 2,4-dinitrofluorobenzene-induced dermatitis in mice. Immunopharmacol. Immunotoxicol. 2016, 38, 327–333. [Google Scholar] [CrossRef]
- Lu, Q.; Tong, B.; Luo, Y.; Sha, L.; Chou, G.; Wang, Z.; Xia, Y.; Dai, Y. Norisoboldine suppresses VEGF-induced endothelial cell migration via the cAMP-PKA-NF-kappaB/Notch1 pathway. PLoS ONE 2013, 8, e81220. [Google Scholar] [CrossRef]
- Wei, Z.F.; Tong, B.; Xia, Y.F.; Lu, Q.; Chou, G.X.; Wang, Z.T.; Dai, Y. Norisoboldine suppresses osteoclast differentiation through preventing the accumulation of TRAF6-TAK1 complexes and activation of MAPKs/NF-kappaB/c-Fos/NFATc1 Pathways. PLoS ONE 2013, 8, e59171. [Google Scholar]
- Tong, B.; Dou, Y.; Wang, T.; Yu, J.; Wu, X.; Lu, Q.; Chou, G.; Wang, Z.; Kong, L.; Dai, Y.; et al. Norisoboldine ameliorates collagen-induced arthritis through regulating the balance between Th17 and regulatory T cells in gut-associated lymphoid tissues. Toxicol. Appl. Pharmacol. 2015, 282, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Wang, F.; Song, J.; Lu, Q.; Zhao, P.; Xia, Y.; Chou, G.; Wang, Z.; Dai, Y. Norisoboldine inhibits the production of interleukin-6 in fibroblast-like synoviocytes from adjuvant arthritis rats through PKC/MAPK/NF-kappaB-p65/CREB pathways. J. Cell. Biochem. 2012, 113, 2785–2795. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Lu, S.; Gao, X.; Luo, Y.; Tong, B.; Wei, Z.; Lu, T.; Xia, Y.; Chou, G.; Wang, Z. Norisoboldine, an alkaloid compound isolated from Radix Linderae, inhibits synovial angiogenesis in adjuvant-induced arthritis rats by moderating Notch1 pathway-related endothelial tip cell phenotype. Exp. Biol. Med. 2012, 237, 919–932. [Google Scholar] [CrossRef]
- Xing, D.; Li, Q.; Lin, G.; Lin, H.; Kang, W.; Zhang, M.; Ding, R.; Li, N. The protective effects of propofol against renal ischemia-reperfusion injury are potentiated by norisoboldine treatment via inhibition of oxidative stress pathways. J. Biochem. Mol. Toxicol. 2022, 36, e22937. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Kim, J.W.; Lee, M.N.; Oh, S.H.; Piao, X.; Wang, Z.; Kwon, S.H.; Kim, O.S.; Koh, J.T. Isolation of High Purity Mouse Mesenchymal Stem Cells through Depleting Macrophages Using Liposomal Clodronate. Tissue Eng. Regen. Med. 2022, 19, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Zhang, Z.; Xu, Z.; Ouyang, H.; Wang, L.; Greenblatt, M.; Chen, X.; Zou, W. H3K36 methyltransferase NSD1 regulates chondrocyte differentiation for skeletal development and fracture repair. Bone Res. 2021, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Jiang, W.; Zhou, N.; Liao, J.; Yang, M.; Hu, N.; Liang, X.; Xu, W.; Chen, H.; Liu, W.; et al. Sox9 augments BMP2-induced chondrogenic differentiation by downregulating Smad7 in mesenchymal stem cells (MSCs). Genes Dis. 2017, 4, 229–239. [Google Scholar] [CrossRef]
- Legendre, F.; Ollitrault, D.; Gomez-Leduc, T.; Bouyoucef, M.; Hervieu, M.; Gruchy, N.; Mallein-Gerin, F.; Leclercq, S.; Demoor, M.; Galera, P. Enhanced chondrogenesis of bone marrow-derived stem cells by using a combinatory cell therapy strategy with BMP-2/TGF-beta1, hypoxia, and COL1A1/HtrA1 siRNAs. Sci. Rep. 2017, 7, 3406. [Google Scholar] [CrossRef]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef]
- Hu, D.P.; Ferro, F.; Yang, F.; Taylor, A.J.; Chang, W.; Miclau, T.; Marcucio, R.S.; Bahney, C.S. Cartilage to bone transformation during fracture healing is coordinated by the invading vasculature and induction of the core pluripotency genes. Development 2017, 144, 221–234. [Google Scholar] [CrossRef]
- Kodama, J.; Wilkinson, K.J.; Iwamoto, M.; Otsuru, S.; Enomoto-Iwamoto, M. The role of hypertrophic chondrocytes in regulation of the cartilage-to-bone transition in fracture healing. Bone Rep. 2022, 17, 101616. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, A.; Nakajima, A.; Nakajima, F.; Goto, K.; Yamazaki, M. Molecular basis for affected cartilage formation and bone union in fracture healing of the streptozotocin-induced diabetic rat. Bone 2008, 43, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Kayal, R.A.; Alblowi, J.; McKenzie, E.; Krothapalli, N.; Silkman, L.; Gerstenfeld, L.; Einhorn, T.A.; Graves, D.T. Diabetes causes the accelerated loss of cartilage during fracture repair which is reversed by insulin treatment. Bone 2009, 44, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Henneicke, H.; Zhang, Y.; Stoner, S.; Cheng, T.L.; Schindeler, A.; Chen, D.; Tuckermann, J.; Cooper, M.S.; Seibel, M.J.; et al. Disruption of glucocorticoid signaling in chondrocytes delays metaphyseal fracture healing but does not affect normal cartilage and bone development. Bone 2014, 69, 12–22. [Google Scholar] [CrossRef]
- Zhang, C.; Feinberg, D.; Alharbi, M.; Ding, Z.; Lu, C.; O’Connor, J.P.; Graves, D.T. Chondrocytes Promote Vascularization in Fracture Healing Through a FOXO1-Dependent Mechanism. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2019, 34, 547–556. [Google Scholar] [CrossRef]
- Bahney, C.S.; Zondervan, R.L.; Allison, P.; Theologis, A.; Ashley, J.W.; Ahn, J.; Miclau, T.; Marcucio, R.S.; Hankenson, K.D. Cellular biology of fracture healing. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2019, 37, 35–50. [Google Scholar] [CrossRef]
- Kitaori, T.; Ito, H.; Schwarz, E.M.; Tsutsumi, R.; Yoshitomi, H.; Oishi, S.; Nakano, M.; Fujii, N.; Nagasawa, T.; Nakamura, T. Stromal cell-derived factor 1/CXCR4 signaling is critical for the recruitment of mesenchymal stem cells to the fracture site during skeletal repair in a mouse model. Arthritis Rheum. 2009, 60, 813–823. [Google Scholar] [CrossRef]
- Li, X.; Gao, Z.; Wang, J. Single percutaneous injection of stromal cell-derived factor-1 induces bone repair in mouse closed tibial fracture model. Orthopedics 2011, 34, 450. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Inzana, J.A.; Mirando, A.J.; Ren, Y.; Liu, Z.; Shen, J.; O’Keefe, R.J.; Awad, H.A.; Hilton, M.J. NOTCH signaling in skeletal progenitors is critical for fracture repair. J. Clin. Investig. 2016, 126, 1471–1481. [Google Scholar] [CrossRef]
- Shu, B.; Zhang, M.; Xie, R.; Wang, M.; Jin, H.; Hou, W.; Tang, D.; Harris, S.E.; Mishina, Y.; O’Keefe, R.J.; et al. BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development. J. Cell Sci. 2011, 124 Pt 20, 3428–3440. [Google Scholar] [CrossRef]
- Zhou, N.; Li, Q.; Lin, X.; Hu, N.; Liao, J.Y.; Lin, L.B.; Zhao, C.; Hu, Z.M.; Liang, X.; Xu, W.; et al. BMP2 induces chondrogenic differentiation, osteogenic differentiation and endochondral ossification in stem cells. Cell Tissue Res. 2016, 366, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Wang, K.; Qiao, S.M.; Dai, Y.; Wei, Z.F. Norisoboldine, a natural aryl hydrocarbon receptor agonist, alleviates TNBS-induced colitis in mice, by inhibiting the activation of NLRP3 inflammasome. Chin. J. Nat. Med. 2018, 16, 161–174. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| Aggrecan | CCTGCTACTTCATCGACCCC | AGATGCTGTTGACTCGAACCT |
| Bmp2 | GGGACCCGCTGTCTTCTAGT | TCAACTCAAATTCGCTGAGGAC |
| Gapdh | AGGTCGGTGTGAACGGATTTG | TGTAGACCATGTAGTTGAGGTCA |
| Col2a1 | GGGAATGTCCTCTGCGATGAC | GAAGGGGATCTCGGGGTTG |
| Alk2 | GTGGAAGATTACAAGCCACCA | GGGTCTGAGAACCATCTGTTAGG |
| Il-6 | TAGTCCTTCCTACCCCAATTTCC | TTGGTCCTTAGCCACTCCTTC |
| Il-1β | GCAACTGTTCCTGAACTCAACT | ATCTTTTGGGGTCCGTCAACT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, W.; Shen, M.; Sun, K.; Li, S.; Miao, J.; Wang, J.; Xu, J.; Wen, P.; Zhang, Q. Norisoboldine, a Natural Isoquinoline Alkaloid, Inhibits Diaphyseal Fracture Healing in Mice by Alleviating Cartilage Formation. Biomedicines 2023, 11, 2031. https://doi.org/10.3390/biomedicines11072031
Yan W, Shen M, Sun K, Li S, Miao J, Wang J, Xu J, Wen P, Zhang Q. Norisoboldine, a Natural Isoquinoline Alkaloid, Inhibits Diaphyseal Fracture Healing in Mice by Alleviating Cartilage Formation. Biomedicines. 2023; 11(7):2031. https://doi.org/10.3390/biomedicines11072031
Chicago/Turabian StyleYan, Wenliang, Meng Shen, Kainong Sun, Shiming Li, Jingyuan Miao, Jun Wang, Jiayang Xu, Pengcheng Wen, and Qian Zhang. 2023. "Norisoboldine, a Natural Isoquinoline Alkaloid, Inhibits Diaphyseal Fracture Healing in Mice by Alleviating Cartilage Formation" Biomedicines 11, no. 7: 2031. https://doi.org/10.3390/biomedicines11072031
APA StyleYan, W., Shen, M., Sun, K., Li, S., Miao, J., Wang, J., Xu, J., Wen, P., & Zhang, Q. (2023). Norisoboldine, a Natural Isoquinoline Alkaloid, Inhibits Diaphyseal Fracture Healing in Mice by Alleviating Cartilage Formation. Biomedicines, 11(7), 2031. https://doi.org/10.3390/biomedicines11072031