

Intra-Articular Hyaluronic Acid in Osteoarthritis and Tendinopathies: Molecular and Clinical Approaches

 , ,
, ,  , ,
, ,  ,
,
Abstract
1. Introduction
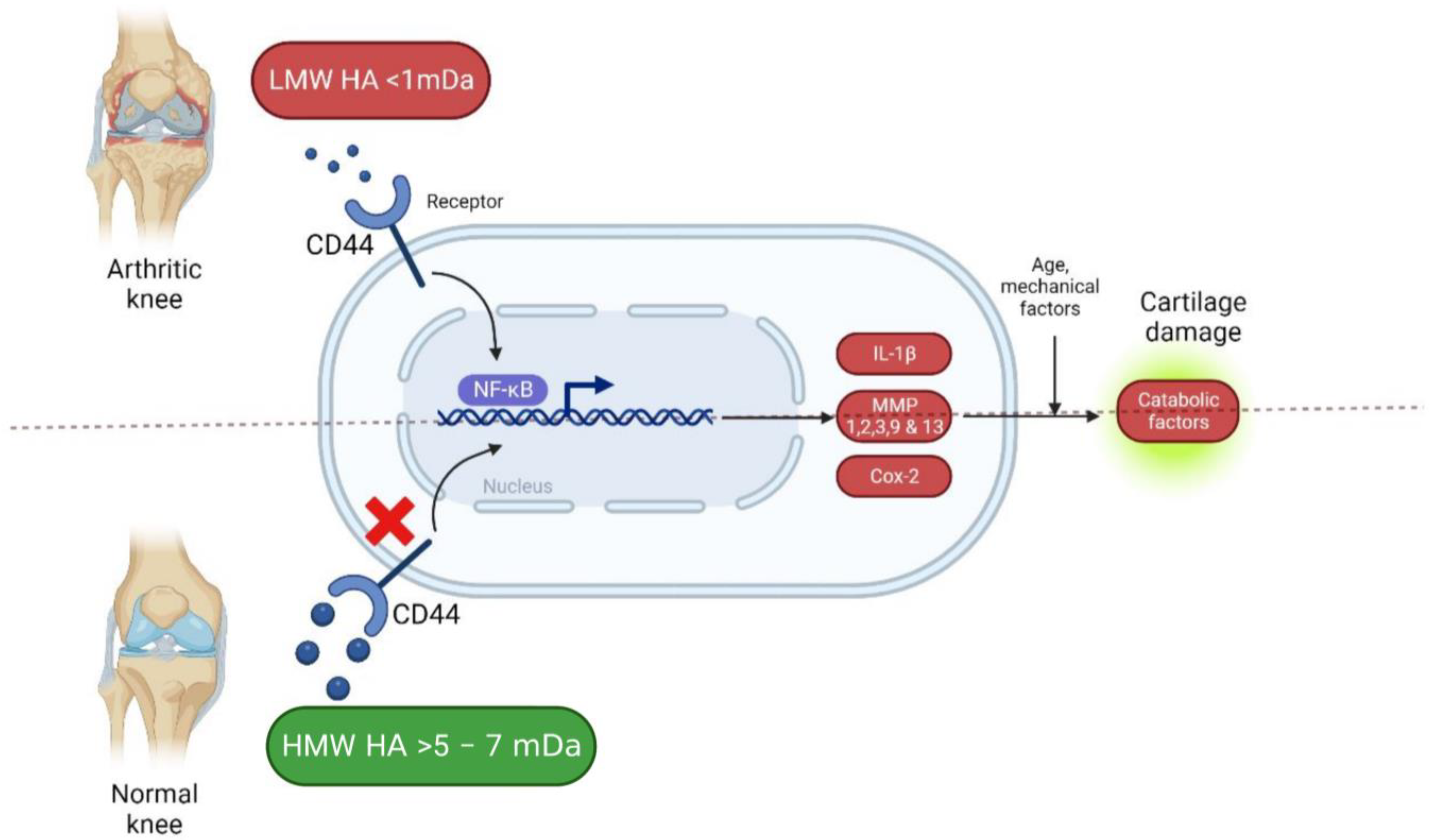
2. Etiopathogenesis of Osteoarthritis
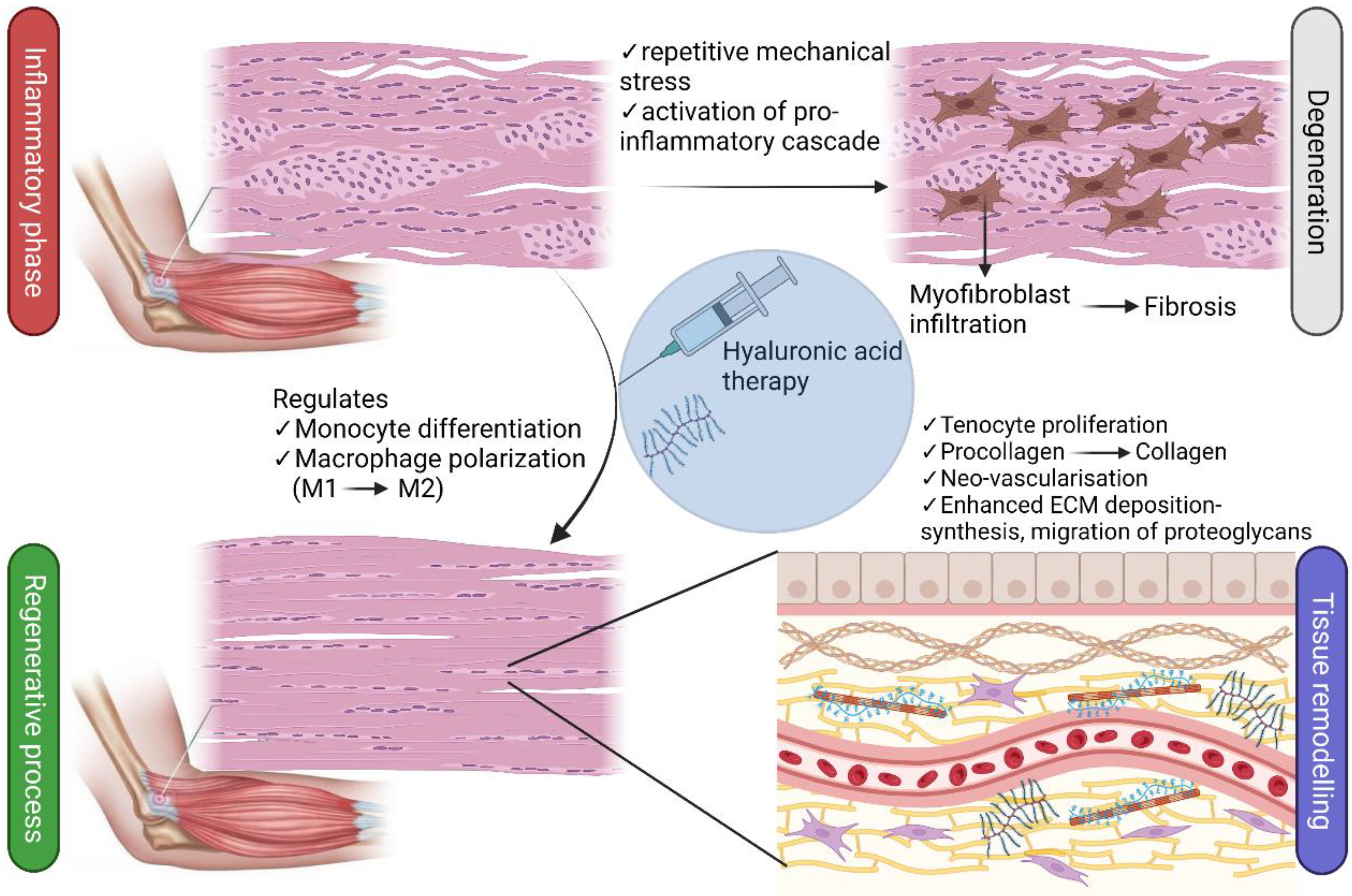
3. Etiopathogenesis of Tendinopathy
4. Physicochemical and Biological Properties of Hyaluronic Acid
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, R.C.; Lall, R.; Srivastava, A.; Sinha, A. Hyaluronic Acid: Molecular Mechanisms and Therapeutic Trajectory. Front. Vet. Sci. 2019, 6, 192. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, Y.; Li, J.; Du, G.; Chen, J. Microbial Production of Hyaluronic Acid: Current State, Challenges, and Perspectives. Microb. Cell Factories 2011, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Fallacara, A.; Baldini, E.; Manfredini, S.; Vertuani, S. Hyaluronic Acid in the Third Millennium. Polymers 2018, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Hermans, J.; Bierma-Zeinstra, S.M.A.; Bos, P.K.; Niesten, D.D.; Verhaar, J.A.N.; Reijman, M. The Effectiveness of High Molecular Weight Hyaluronic Acid for Knee Osteoarthritis in Patients in the Working Age: A Randomised Controlled Trial. BMC Musculoskelet. Disord. 2019, 20, 196. [Google Scholar] [CrossRef]
- Muneta, T.; Koga, H.; Ju, Y.-J.; Mochizuki, T.; Sekiya, I. Hyaluronan Injection Therapy for Athletic Patients with Patellar Tendinopathy. J. Orthop. Sci. 2012, 17, 425–431. [Google Scholar] [CrossRef]
- Liang, J.; Jiang, D.; Noble, P.W. Hyaluronan as a Therapeutic Target in Human Diseases. Adv. Drug Deliv. Rev. 2016, 97, 186–203. [Google Scholar] [CrossRef]
- Hunter, D.J.; March, L.; Chew, M. Osteoarthritis in 2020 and beyond: A Lancet Commission. Lancet 2020, 396, 1711–1712. [Google Scholar] [CrossRef]
- Global Burden of Disease Collaborative Network. Global Burden of Disease Study 2019 (GBD 2019) Results; Global Burden of Disease Collaborative Network: Seattle, WA, USA, 2019. [Google Scholar]
- Zhang, Y.; Jordan, J.M. Epidemiology of Osteoarthritis. Clin. Geriatr. Med. 2010, 26, 355–369. [Google Scholar] [CrossRef]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.J. Osteoarthritis: Toward a Comprehensive Understanding of Pathological Mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [CrossRef]
- Azzini, G.O.M.; Santos, G.S.; Visoni, S.B.C.; Azzini, V.O.M.; dos Santos, R.G.; Huber, S.C.; Lana, J.F. Metabolic Syndrome and Subchondral Bone Alterations: The Rise of Osteoarthritis—A Review. J. Clin. Orthop. Trauma 2020, 11, S849–S855. [Google Scholar] [CrossRef]
- Mora, J.C.; Przkora, R.; Cruz-Almeida, Y. Knee Osteoarthritis: Pathophysiology and Current Treatment Modalities. J. Pain Res. 2018, 11, 2189–2196. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef]
- Lana, J.F.; Macedo, A.; Ingrao, I.L.G.; Huber, S.C.; Santos, G.S.; Santana, M.H.A. Leukocyte-Rich PRP for Knee Osteoarthritis: Current Concepts. J. Clin. Orthop. Trauma 2019, 10, S179–S182. [Google Scholar] [CrossRef]
- Man, G.S.; Mologhianu, G. Osteoarthritis Pathogenesis—A Complex Process That Involves the Entire Joint. J. Med. Life 2014, 7, 37–41. [Google Scholar]
- Yuan, G.-H.; Tanaka, M.; Masuko-Hongo, K.; Shibakawa, A.; Kato, T.; Nishioka, K.; Nakamura, H. Characterization of Cells from Pannus-like Tissue over Articular Cartilage of Advanced Osteoarthritis. Osteoarthr. Cartil. 2004, 12, 38–45. [Google Scholar] [CrossRef]
- Sellam, J.; Berenbaum, F. The Role of Synovitis in Pathophysiology and Clinical Symptoms of Osteoarthritis. Nat. Rev. Rheumatol. 2010, 6, 625–635. [Google Scholar] [CrossRef]
- Lee, J.H.; Ort, T.; Ma, K.; Picha, K.; Carton, J.; Marsters, P.A.; Lohmander, L.S.; Baribaud, F.; Song, X.-Y.R.; Blake, S. Resistin Is Elevated Following Traumatic Joint Injury and Causes Matrix Degradation and Release of Inflammatory Cytokines from Articular Cartilage in Vitro. Osteoarthr. Cartil. 2009, 17, 613–620. [Google Scholar] [CrossRef]
- Presle, N.; Pottie, P.; Dumond, H.; Guillaume, C.; Lapicque, F.; Pallu, S.; Mainard, D.; Netter, P.; Terlain, B. Differential Distribution of Adipokines between Serum and Synovial Fluid in Patients with Osteoarthritis. Contribution of Joint Tissues to Their Articular Production. Osteoarthr. Cartil. 2006, 14, 690–695. [Google Scholar] [CrossRef]
- Hasegawa, M.; Segawa, T.; Maeda, M.; Yoshida, T.; Sudo, A. Thrombin-Cleaved Osteopontin Levels in Synovial Fluid Correlate with Disease Severity of Knee Osteoarthritis. J. Rheumatol. 2011, 38, 129–134. [Google Scholar] [CrossRef]
- Krasnokutsky, S.; Attur, M.; Palmer, G.; Samuels, J.; Abramson, S.B. Current Concepts in the Pathogenesis of Osteoarthritis. Osteoarthr. Cartil. 2008, 16, S1–S3. [Google Scholar] [CrossRef]
- Ashraf, S.; Walsh, D.A. Angiogenesis in Osteoarthritis. Curr. Opin. Rheumatol. 2008, 20, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Marcu, K.B. Cartilage Homeostasis in Health and Rheumatic Diseases. Arthritis Res. 2009, 11, 224. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, S.; Huang, J.; Guo, W.; Chen, J.; Zhang, L.; Zhao, B.; Peng, J.; Wang, A.; Wang, Y.; et al. The ECM-Cell Interaction of Cartilage Extracellular Matrix on Chondrocytes. BioMed Res. Int. 2014, 2014, e648459. [Google Scholar] [CrossRef] [PubMed]
- Heijink, A.; Gomoll, A.H.; Madry, H.; Drobnič, M.; Filardo, G.; Espregueira-Mendes, J.; Van Dijk, C.N. Biomechanical Considerations in the Pathogenesis of Osteoarthritis of the Knee. Knee Surg. Sport. Traumatol. Arthrosc. 2012, 20, 423–435. [Google Scholar] [CrossRef]
- Wang, M.; Peng, Z.; Vasilev, K.; Ketheesan, N. Investigation of Wear Particles Generated in Human Knee Joints Using Atomic Force Microscopy. Tribol. Lett. 2013, 51, 161–170. [Google Scholar] [CrossRef]
- Stannus, O.; Jones, G.; Cicuttini, F.; Parameswaran, V.; Quinn, S.; Burgess, J.; Ding, C. Circulating Levels of IL-6 and TNF-α Are Associated with Knee Radiographic Osteoarthritis and Knee Cartilage Loss in Older Adults. Osteoarthr. Cartil. 2010, 18, 1441–1447. [Google Scholar] [CrossRef]
- Buckwalter, J.A.; Mankin, H.J.; Grodzinsky, A.J. Articular Cartilage and Osteoarthritis. Instr. Course Lect. 2005, 54, 465–480. [Google Scholar]
- Loiacono, C.; Palermi, S.; Massa, B.; Belviso, I.; Romano, V.; Di Gregorio, A.; Sirico, F.; Sacco, A.M. Tendinopathy: Pathophysiology, Therapeutic Options, and Role of Nutraceutics. A Narrative Literature Review. Medicina 2019, 55, 447. [Google Scholar] [CrossRef]
- Andarawis-Puri, N.; Flatow, E.L.; Soslowsky, L.J. Tendon Basic Science: Development, Repair, Regeneration, and Healing: Tendon Development, Injury, and Repair. J. Orthop. Res. 2015, 33, 780–784. [Google Scholar] [CrossRef]
- Abbah, S.A.; Spanoudes, K.; O’Brien, T.; Pandit, A.; Zeugolis, D.I. Assessment of Stem Cell Carriers for Tendon Tissue Engineering in Pre-Clinical Models. Stem. Cell Res. 2014, 5, 38. [Google Scholar] [CrossRef]
- Maffulli, N.; Wong, J.; Almekinders, L.C. Types and Epidemiology of Tendinopathy. Clin. Sport. Med. 2003, 22, 675–692. [Google Scholar] [CrossRef]
- Albers, I.S.; Zwerver, J.; Diercks, R.L.; Dekker, J.H.; Van den Akker-Scheek, I. Incidence and Prevalence of Lower Extremity Tendinopathy in a Dutch General Practice Population: A Cross Sectional Study. BMC Musculoskelet. Disord 2016, 17, 16. [Google Scholar] [CrossRef]
- Morton, S.; Williams, S.; Valle, X.; Diaz-Cueli, D.; Malliaras, P.; Morrissey, D. Patellar Tendinopathy and Potential Risk Factors: An International Database of Cases and Controls. Clin. J. Sport Med. 2017, 27, 468–474. [Google Scholar] [CrossRef]
- Abate, M.; Schiavone, C.; Salini, V.; Andia, I. Occurrence of Tendon Pathologies in Metabolic Disorders. Rheumatology 2013, 52, 599–608. [Google Scholar] [CrossRef]
- Li, Y.; Fessel, G.; Georgiadis, M.; Snedeker, J.G. Advanced Glycation End-Products Diminish Tendon Collagen Fiber Sliding. Matrix Biol. 2013, 32, 169–177. [Google Scholar] [CrossRef]
- Couppé, C.; Svensson, R.B.; Kongsgaard, M.; Kovanen, V.; Grosset, J.-F.; Snorgaard, O.; Bencke, J.; Larsen, J.O.; Bandholm, T.; Christensen, T.M.; et al. Human Achilles Tendon Glycation and Function in Diabetes. J. Appl. Physiol. 2016, 120, 130–137. [Google Scholar] [CrossRef]
- Abate, M.; Gravare Silbernagel, K.; Siljeholm, C.; Di Iorio, A.; De Amicis, D.; Salini, V.; Werner, S.; Paganelli, R. Pathogenesis of Tendinopathies: Inflammation or Degeneration? Arthritis Res. Ther. 2009, 11, 235. [Google Scholar] [CrossRef]
- Thorpe, C.T.; Screen, H.R.C. Tendon Structure and Composition. In Metabolic Influences on Risk for Tendon Disorders; Advances in Experimental Medicine and, Biology; Ackermann, P.W., Hart, D.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 920, pp. 3–10. ISBN 978-3-319-33941-2. [Google Scholar]
- Buckley, M.R.; Evans, E.; Satchel, L.N.; Matuszewski, P.E.; Chen, Y.-L.; Elliott, D.M.; Soslowsky, L.J.; Dodge, G.R. Distributions of Types I, II and III Collagen by Region in the Human Supraspinatus Tendon. Connect. Tissue Res. 2013, 54, 374–379. [Google Scholar] [CrossRef]
- Sharma, P.; Maffulli, N. Tendon Injury and Tendinopathy: Healing and Repair. J. Bone Jt. Surg. 2005, 87, 187–202. [Google Scholar] [CrossRef]
- Screen, H.R.C.; Berk, D.E.; Kadler, K.E.; Ramirez, F.; Young, M.F. Tendon Functional Extracellular Matrix. J. Orthop. Res. 2015, 33, 793–799. [Google Scholar] [CrossRef]
- Bosman, F.T.; Stamenkovic, I. Functional Structure and Composition of the Extracellular Matrix. J. Pathol. 2003, 200, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the Extracellular Matrix in Development and Disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Schilling, T.F. Tendon Development and Musculoskeletal Assembly: Emerging Roles for the Extracellular Matrix. Development 2015, 142, 4191–4204. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, P.W. Neuronal Regulation of Tendon Homoeostasis. Int. J. Exp. Pathol. 2013, 94, 271–286. [Google Scholar] [CrossRef]
- Killian, M.L.; Cavinatto, L.; Galatz, L.M.; Thomopoulos, S. The Role of Mechanobiology in Tendon Healing. J. Shoulder Elb. Surg. 2012, 21, 228–237. [Google Scholar] [CrossRef]
- Guzzoni, V.; Selistre-de-Araújo, H.; de Cássia Marqueti, R. Tendon Remodeling in Response to Resistance Training, Anabolic Androgenic Steroids and Aging. Cells 2018, 7, 251. [Google Scholar] [CrossRef]
- Magnusson, S.P.; Kjaer, M. The Impact of Loading, Unloading, Ageing and Injury on the Human Tendon. J. Physiol. 2019, 597, 1283–1298. [Google Scholar] [CrossRef]
- Magnusson, S.P.; Langberg, H.; Kjaer, M. The Pathogenesis of Tendinopathy: Balancing the Response to Loading. Nat. Rev. Rheumatol. 2010, 6, 262–268. [Google Scholar] [CrossRef]
- Snedeker, J.G.; Foolen, J. Tendon Injury and Repair—A Perspective on the Basic Mechanisms of Tendon Disease and Future Clinical Therapy. Acta Biomater. 2017, 63, 18–36. [Google Scholar] [CrossRef]
- Yang, G.; Im, H.J.; Wang, J.H.C. Repetitive Mechanical Stretching Modulates IL-1β Induced COX-2, MMP-1 Expression, and PGE2 Production in Human Patellar Tendon Fibroblasts. Gene 2005, 363, 166–172. [Google Scholar] [CrossRef]
- Khan, M.H.; Li, Z.; Wang, J.H.C. Repeated Exposure of Tendon to Prostaglandin-E2 Leads to Localized Tendon Degeneration. Clin. J. Sport Med. 2005, 15, 27–33. [Google Scholar] [CrossRef]
- Sullo, A.; Maffulli, N.; Capasso, G.; Testa, V. The Effects of Prolonged Peritendinous Administration of PGE1 to the Rat Achilles Tendon: A Possible Animal Model of Chronic Achilles Tendinopathy. J. Orthop. Sci. 2001, 6, 349–357. [Google Scholar] [CrossRef]
- Morita, W.; Dakin, S.G.; Snelling, S.J.B.; Carr, A.J. Cytokines in Tendon Disease: A Systematic Review. Bone Jt. Res. 2017, 6, 656–664. [Google Scholar] [CrossRef]
- Tang, C.; Chen, Y.; Huang, J.; Zhao, K.; Chen, X.; Yin, Z.; Heng, B.C.; Chen, W.; Shen, W. The Roles of Inflammatory Mediators and Immunocytes in Tendinopathy. J. Orthop. Transl. 2018, 14, 23–33. [Google Scholar] [CrossRef]
- Abraham, A.C.; Shah, S.A.; Golman, M.; Song, L.; Li, X.; Kurtaliaj, I.; Akbar, M.; Millar, N.L.; Abu-Amer, Y.; Galatz, L.M.; et al. Targeting the NF-KB Signaling Pathway in Chronic Tendon Disease. Sci. Transl. Med. 2019, 11, eaav4319. [Google Scholar] [CrossRef]
- Schubert, T.E.O.; Weidler, C.; Lerch, K.; Hofstädter, F.; Straub, R.H. Achilles Tendinosis Is Associated with Sprouting of Substance P Positive Nerve Fibres. Ann. Rheum. Dis. 2005, 64, 1083–1086. [Google Scholar] [CrossRef]
- Lana, J.F.J.F.; Huber, S.C.S.C.; Purita, J.; Tambeli, C.H.C.H.; Santos, G.S.G.S.; Paulus, C.; Annichino-Bizzacchi, J.M.J.M. Leukocyte-Rich PRP versus Leukocyte-Poor PRP—The Role of Monocyte/Macrophage Function in the Healing Cascade. J. Clin. Orthop. Trauma 2019, 10, S7–S12. [Google Scholar] [CrossRef]
- Orekhov, A.N.; Orekhova, V.A.; Nikiforov, N.G.; Myasoedova, V.A.; Grechko, A.V.; Romanenko, E.B.; Zhang, D.; Chistiakov, D.A. Monocyte Differentiation and Macrophage Polarization. Vessel Plus 2019, 2019, 10. [Google Scholar] [CrossRef]
- Arvind, V.; Huang, A.H. Reparative and Maladaptive Inflammation in Tendon Healing. Front. Bioeng. Biotechnol. 2021, 9, 719047. [Google Scholar] [CrossRef]
- Kaji, D.A.; Howell, K.L.; Balic, Z.; Hubmacher, D.; Huang, A.H. Tgfβ Signaling Is Required for Tenocyte Recruitment and Functional Neonatal Tendon Regeneration. eLife 2020, 9, e51779. [Google Scholar] [CrossRef]
- Ackermann, P.W.; Renström, P. Tendinopathy in Sport. Sports Health 2012, 4, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Setti, T.; Arab, M.G.L.; Santos, G.S.; Alkass, N.; Andrade, M.A.P.; Lana, J.F.S.D. The Protective Role of Glutathione in Osteoarthritis. J. Clin. Orthop. Trauma 2020, 15, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Hafsi, K.; McKay, J.; Li, J.; Lana, J.F.; Macedo, A.; Santos, G.S.; Murrell, W.D. Nutritional, Metabolic and Genetic Considerations to Optimise Regenerative Medicine Outcome for Knee Osteoarthritis. J. Clin. Orthop. Trauma 2019, 10, 2–8. [Google Scholar] [CrossRef]
- Marcum, Z.A.; Hanlon, J.T. Recognizing the Risks of Chronic Nonsteroidal Anti-Inflammatory Drug Use in Older Adults. Ann. Long-Term Care 2010, 18, 24–27. [Google Scholar]
- Kruel, A.V.S.; Ribeiro, L.L.; Gusmão, P.D.; Huber, S.C.; Lana, J.F.S.D. Orthobiologics in the Treatment of Hip Disorders. World J. Stem. Cells 2021, 13, 304–316. [Google Scholar] [CrossRef]
- Maheu, E.; Rannou, F.; Reginster, J.Y. Efficacy and Safety of Hyaluronic Acid in the Management of Osteoarthritis: Evidence from Real-Life Setting Trials and Surveys. Semin. Arthritis Rheum. 2016, 45, S28–S33. [Google Scholar] [CrossRef]
- Prieto, J.G.; Pulido, M.M.; Zapico, J.; Molina, A.J.; Gimeno, M.; Coronel, P.; Alvarez, A.I. Comparative Study of Hyaluronic Derivatives: Rheological Behaviour, Mechanical and Chemical Degradation. Int. J. Biol. Macromol. 2005, 35, 63–69. [Google Scholar] [CrossRef]
- Ghosh, P.; Guidolin, D. Potential Mechanism of Action of Intra-Articular Hyaluronan Therapy in Osteoarthritis: Are the Effects Molecular Weight Dependent? Semin. Arthritis Rheum. 2002, 32, 10–37. [Google Scholar] [CrossRef] [PubMed]
- Guidolin, D.D.; Ronchetti, I.P.; Lini, E.; Guerra, D.; Frizziero, L. Morphological Analysis of Articular Cartilage Biopsies from a Randomized, Clinical Study Comparing the Effects of 500–730 KDa Sodium Hyaluronate (Hyalgan®) and Methylprednisolone Acetate on Primary Osteoarthritis of the Knee. Osteoarthr. Cartil. 2001, 9, 371–381. [Google Scholar] [CrossRef]
- Bruyère, O.; Cooper, C.; Pelletier, J.P.; Branco, J.; Luisa Brandi, M.; Guillemin, F.; Hochberg, M.C.; Kanis, J.A.; Kvien, T.K.; Martel-Pelletier, J.; et al. An Algorithm Recommendation for the Management of Knee Osteoarthritis in Europe and Internationally: A Report from a Task Force of the European Society for Clinical and Economic Aspects of Osteoporosis and Osteoarthritis (ESCEO). Semin. Arthritis Rheum. 2014, 44, 253–263. [Google Scholar] [CrossRef]
- Lajeunesse, D.; Delalandre, A.; Martel-Pelletier, J.; Pelletier, J.P. Hyaluronic Acid Reverses the Abnormal Synthetic Activity of Human Osteoarthritic Subchondral Bone Osteoblasts. Bone 2003, 33, 703–710. [Google Scholar] [CrossRef]
- Brun, P.; Zavan, B.; Vindigni, V.; Schiavinato, A.; Pozzuoli, A.; Iacobellis, C.; Abatangelo, G. In Vitro Response of Osteoarthritic Chondrocytes and Fibroblast-like Synoviocytes to a 500-730 KDa Hyaluronan Amide Derivative. J. Biomed. Mater. Res. Part B Appl. Biomater. 2012, 100, 2073–2081. [Google Scholar] [CrossRef]
- Campo, G.M.; Avenoso, A.; Campo, S.; D’Ascola, A.; Traina, P.; Rugolo, C.A.; Calatroni, A. Differential Effect of Molecular Mass Hyaluronan on Lipopolysaccharide-Induced Damage in Chondrocytes. Innate Immun 2010, 16, 48–63. [Google Scholar] [CrossRef]
- Day, A.J.; de la Motte, C.A. Hyaluronan Cross-Linking: A Protective Mechanism in Inflammation? Trends Immunol. 2005, 26, 637–643. [Google Scholar] [CrossRef]
- Julovi, S.M.; Yasuda, T.; Shimizu, M.; Hiramitsu, T.; Nakamura, T. Inhibition of Interleukin-1β-Stimulated Production of Matrix Metalloproteinases by Hyaluronan via CD44 in Human Articular Cartilage. Arthritis Rheum. 2004, 50, 516–525. [Google Scholar] [CrossRef]
- Kalaci, A.; Yilmaz, R.H.; Aslan, B.; Söğüt, S.; Yanat, A.N.; Uz, E. Effects of Hyaluronan on Nitric Oxide Levels and Superoxide Dismutase Activities in Synovial Fluid in Knee Osteoarthritis. Clin. Rheumatol. 2007, 26, 1306–1311. [Google Scholar] [CrossRef]
- Karna, E.; Miltyk, W.; Surażyński, A.; Pałka, J.A. Protective Effect of Hyaluronic Acid on Interleukin-1-Induced Deregulation of Βeta 1 -Integrin and Insulin-like Growth Factor-I Receptor Signaling and Collagen Biosynthesis in Cultured Human Chondrocytes. Mol. Cell. Biochem. 2008, 308, 57–64. [Google Scholar] [CrossRef]
- Abate, M.; Pelotti, P.; De Amicis, D.; Di Iorio, A.; Galletti, S.; Salini, V. Viscosupplementation with Hyaluronic Acid in Hip Osteoarthritis (a Review). Upsala J. Med. Sci. 2008, 113, 261–278. [Google Scholar] [CrossRef]
- Dicker, K.T.; Gurski, L.A.; Pradhan-Bhatt, S.; Witt, R.L.; Farach-Carson, M.C.; Jia, X. Hyaluronan: A Simple Polysaccharide with Diverse Biological Functions. Acta Biomater. 2014, 10, 1558–1570. [Google Scholar] [CrossRef]
- Abatangelo, G.; Vindigni, V.; Avruscio, G.; Pandis, L.; Brun, P. Hyaluronic Acid: Redefining Its Role. Cells 2020, 9, 1743. [Google Scholar] [CrossRef]
- Vigetti, D.; Karousou, E.; Viola, M.; Deleonibus, S.; De Luca, G.; Passi, A. Hyaluronan: Biosynthesis and Signaling. Biochim. Biophys. Acta 2014, 1840, 2452–2459. [Google Scholar] [CrossRef] [PubMed]
- Kouvidi, K.; Berdiaki, A.; Nikitovic, D.; Katonis, P.; Afratis, N.; Hascall, V.C.; Karamanos, N.K.; Tzanakakis, G.N. Role of Receptor for Hyaluronic Acid-Mediated Motility (RHAMM) in Low Molecular Weight Hyaluronan (LMWHA)-Mediated Fibrosarcoma Cell Adhesion. J. Biol. Chem. 2011, 286, 38509–38520. [Google Scholar] [CrossRef]
- McCain, J.P.; Balazs, E.A.; de la Rua, H. Preliminary Studies on the Use of a Viscoelastic Solution in Arthroscopic Surgery of the Temporomandibular Joint. J. Oral Maxillofac. Surg. 1989, 47, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Altman, R.D.; Moskowitz, R. Intraarticular Sodium Hyaluronate (Hyalgan) in the Treatment of Patients with Osteoarthritis of the Knee: A Randomized Clinical Trial. Hyalgan Study Group. J. Rheumatol. 1998, 25, 2203–2212. [Google Scholar] [PubMed]
- Kolarz, G.; Kotz, R.; Hochmayer, I. Long-Term Benefits and Repeated Treatment Cycles of Intra-Articular Sodium Hyaluronate (Hyalgan) in Patients with Osteoarthritis of the Knee. Semin. Arthritis Rheum. 2003, 32, 310–319. [Google Scholar] [CrossRef]
- Migliore, A.; Bizzi, E.; De Lucia, O.; Delle Sedie, A.; Bentivegna, M.; Mahmoud, A.; Foti, C. Differences among Branded Hyaluronic Acids in Italy, Part 1: Data from In Vitro and Animal Studies and Instructions for Use. Clin. Med. Insights Arthritis Musculoskelet. Disord. 2016, 9, 89–101. [Google Scholar] [CrossRef]
- Salamanna, F.; Giavaresi, G.; Parrilli, A.; Martini, L.; Nicoli Aldini, N.; Abatangelo, G.; Frizziero, A.; Fini, M. Effects of Intra-Articular Hyaluronic Acid Associated to Chitlac (Arty-Duo®) in a Rat Knee Osteoarthritis Model. J. Orthop. Res. 2019, 37, 867–876. [Google Scholar] [CrossRef]
- Tarricone, E.; Mattiuzzo, E.; Belluzzi, E.; Elia, R.; Benetti, A.; Venerando, R.; Vindigni, V.; Ruggieri, P.; Brun, P. Anti-Inflammatory Performance of Lactose-Modified Chitosan and Hyaluronic Acid Mixtures in an In Vitro Macrophage-Mediated Inflammation Osteoarthritis Model. Cells 2020, 9, E1328. [Google Scholar] [CrossRef]
- Tarricone, E.; Elia, R.; Mattiuzzo, E.; Faggian, A.; Pozzuoli, A.; Ruggieri, P.; Brun, P. The Viability and Anti-Inflammatory Effects of Hyaluronic Acid-Chitlac-Tracimolone Acetonide- β-Cyclodextrin Complex on Human Chondrocytes. Cartilage 2021, 13, 920S–924S. [Google Scholar] [CrossRef]
- Hollander, A.P.; Dickinson, S.C.; Sims, T.J.; Brun, P.; Cortivo, R.; Kon, E.; Marcacci, M.; Zanasi, S.; Borrione, A.; De Luca, C.; et al. Maturation of Tissue Engineered Cartilage Implanted in Injured and Osteoarthritic Human Knees. Tissue Eng. 2006, 12, 1787–1798. [Google Scholar] [CrossRef]
- Lana, J.F.S.D.; Weglein, A.; Sampson, S.E.; Vicente, E.F.; Huber, S.C.; Souza, C.V.; Ambach, M.A.; Vincent, H.; Urban-Paffaro, A.; Onodera, C.M.K.; et al. Randomized Controlled Trial Comparing Hyaluronic Acid, Platelet-Rich Plasma and the Combination of Both in the Treatment of Mild and Moderate Osteoarthritis of the Knee. J. Stem. Cells Regen Med. 2016, 12, 69–78. [Google Scholar]
- Levett, P.A.; Hutmacher, D.W.; Malda, J.; Klein, T.J. Hyaluronic Acid Enhances the Mechanical Properties of Tissue-Engineered Cartilage Constructs. PLoS ONE 2014, 9, e113216. [Google Scholar] [CrossRef]
- Gomis, A.; Miralles, A.; Schmidt, R.F.; Belmonte, C. Intra-Articular Injections of Hyaluronan Solutions of Different Elastoviscosity Reduce Nociceptive Nerve Activity in a Model of Osteoarthritic Knee Joint of the Guinea Pig. Osteoarthr. Cartil. 2009, 17, 798–804. [Google Scholar] [CrossRef]
- Kuroki, K.; Cook, J.L.; Kreeger, J.M. Mechanisms of Action and Potential Uses of Hyaluronan in Dogs with Osteoarthritis. J. Am. Vet. Med. Assoc. 2002, 221, 944–950. [Google Scholar] [CrossRef]
- Díaz-Gallego, L.; Prieto, J.G.; Coronel, P.; Gamazo, L.E.; Gimeno, M.; Alvarez, A.I. Apoptosis and Nitric Oxide in an Experimental Model of Osteoarthritis in Rabbit after Hyaluronic Acid Treatment. J. Orthop. Res. 2005, 23, 1370–1376. [Google Scholar] [CrossRef]
- Sun, Y.-L.; Yang, C.; Amadio, P.C.; Zhao, C.; Zobitz, M.E.; An, K.-N. Reducing Friction by Chemically Modifying the Surface of Extrasynovial Tendon Grafts. J. Orthop. Res. 2004, 22, 984–989. [Google Scholar] [CrossRef]
- Nishida, J.; Araki, S.; Akasaka, T.; Toba, T.; Shimamura, T.; Amadio, P.C.; An, K.N. Effect of Hyaluronic Acid on the Excursion Resistance of Tendon Grafts. A Biomechanical Study in a Canine Model in Vitro. J. Bone Jt. Surg. Br. 2004, 86, 918–924. [Google Scholar] [CrossRef]
- Taguchi, M.; Zhao, C.; Sun, Y.-L.; Jay, G.D.; An, K.-N.; Amadio, P.C. The Effect of Surface Treatment Using Hyaluronic Acid and Lubricin on the Gliding Resistance of Human Extrasynovial Tendons in Vitro. J. Hand. Surg. Am. 2009, 34, 1276–1281. [Google Scholar] [CrossRef][Green Version]
- Akasaka, T.; Nishida, J.; Imaeda, T.; Shimamura, T.; Amadio, P.C.; An, K.-N. Effect of Hyaluronic Acid on the Excursion Resistance of Tendon Graft: A Biomechanical in Vitro Study in a Modified Human Model. Clin. Biomech. (Bristol Avon) 2006, 21, 810–815. [Google Scholar] [CrossRef]
- Yamada, T.; Gotoh, M.; Nakama, K.; Mitsui, Y.; Higuchi, F.; Nagata, K. Effects of Hyaluronan on Cell Proliferation and MRNA Expression of Procollagens Alpha 1 (I) and Alpha 1 (III) in Tendon-Derived Fibroblasts from Patients with Rotator Cuff Disease: An in Vitro Study. Am. J. Sport. Med. 2007, 35, 1870–1876. [Google Scholar] [CrossRef]
- Shimpuku, E.; Hamada, K.; Handa, A.; Kobayashi, K.; Nakajima, T.; Yamazaki, H.; Nakamura, M.; Fukuda, H.; Mochida, J. Molecular Effects of Sodium Hyaluronate on the Healing of Avian Supracoracoid Tendon Tear: According to in Situ Hybridization and Real-Time Polymerase Chain Reaction. J. Orthop. Res. 2007, 25, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Wiig, M.; Abrahamsson, S.O. Hyaluronic Acid Modulates Cell Proliferation Unequally in Intrasynovial and Extrasynovial Rabbit Tendons in Vitro. J. Hand Surg. Br. 2000, 25, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Gotoh, M.; Kanazawa, T.; Ohta, K.; Nakamura, K.; Honda, H.; Ohzono, H.; Shimokobe, H.; Mitsui, Y.; Shirachi, I.; et al. Effects of Corticosteroids and Hyaluronic Acid on Torn Rotator Cuff Tendons in Vitro and in Rats. J. Orthop. Res. 2015, 33, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, K.; Ohno, S.; Fujimoto, K.; Honda, K.; Ijuin, C.; Tanaka, N.; Doi, T.; Nakahara, M.; Tanne, K. Proinflammatory Cytokines Regulate the Gene Expression of Hyaluronic Acid Synthetase in Cultured Rabbit Synovial Membrane Cells. Connect Tissue Res. 2001, 42, 187–195. [Google Scholar] [CrossRef]
- Smith, M.M.; Ghosh, P. The Synthesis of Hyaluronic Acid by Human Synovial Fibroblasts Is Influenced by the Nature of the Hyaluronate in the Extracellular Environment. Rheumatol. Int. 1987, 7, 113–122. [Google Scholar] [CrossRef]
- Osti, L.; Berardocco, M.; di Giacomo, V.; Di Bernardo, G.; Oliva, F.; Berardi, A.C. Hyaluronic Acid Increases Tendon Derived Cell Viability and Collagen Type I Expression in Vitro: Comparative Study of Four Different Hyaluronic Acid Preparations by Molecular Weight. BMC Musculoskelet. Disord. 2015, 16, 284. [Google Scholar] [CrossRef]
- Neumann, A.; Schinzel, R.; Palm, D.; Riederer, P.; Münch, G. High Molecular Weight Hyaluronic Acid Inhibits Advanced Glycation Endproduct-Induced NF-KappaB Activation and Cytokine Expression. FEBS Lett. 1999, 453, 283–287. [Google Scholar] [CrossRef]
- Samuelson, E.M.; Ebel, J.A.; Reynolds, S.B.; Arnold, R.M.; Brown, D.E. The Cost-Effectiveness of Platelet-Rich Plasma Compared With Hyaluronic Acid Injections for the Treatment of Knee Osteoarthritis. Arthrosc. J. Arthrosc. Relat. Surg. 2020, 36, 3072–3078. [Google Scholar] [CrossRef]
- Rosen, J.; Niazi, F.; Dysart, S. Cost-Effectiveness of Treating Early to Moderate Stage Knee Osteoarthritis with Intra-Articular Hyaluronic Acid Compared to Conservative Interventions. Adv. Ther. 2020, 37, 344–352. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Brand | Source | Concentration | Molecular Weight | Indication |
|---|---|---|---|---|
| Synolis® VA | Biofermentation | HA: 20 mg (2%) Sorbitol: 40 mg (4%) | High molecular weight (2.1 MDa) | Knee or Hip Osteoarthritis |
| RenehaVis® | Biofermentation | Low molecular weight: 15.4 mg (2.2%); High molecular weight 7 mg (1%) | Low molecular weight (<1 MDa) and High molecular weight (2 MDa) | Knee or Hip Osteoarthritis |
| SportVis® | Biofermentation | 12 mg (1%) | Not reported | Soft tissues (tendon injuries) |
| Ostenil® | Biofermentation | 20 mg (1%) | High molecular weight (1–2 MDa) | Osteoarthritis of shoulders, hips, and knees |
| Ostenil® Plus | Biofermentation | HA: 40 mg (2%) Mannitol: 10 (0.5%) | High molecular weight (1–2 MDa) | Knee Osteoarthritis |
| OrthoVisc® | Biofermentation | 30 mg (1.5%) | High molecular weight (1.1–2.9 MDa) | Knee Osteoarthritis |
| OrthoVisc® mini | Biofermentation | 15 mg (1.5%) | High molecular weight (1.4 MDa) | Small joints |
| MonoVisc® | Biofermentation | 80 mg | High molecular weight (1–2.9 MDa) | Knee Osteoarthritis |
| Synvisc® | Rooster Comb | 16 mg (80% HA HMW cross-linked; 20% gel cross-linked) | High molecular weight (6 MDa) | Osteoarthritis of shoulders, hips, and knees |
| Synvisc® One | Rooster Comb | 48 mg (80% HA HMW cross-linked; 20% gel cross-linked) | High molecular weight (6 MDa) | Knee Osteoarthritis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, F.R.; Costa Marques, M.R.; Costa, V.C.; Santos, G.S.; Martins, R.A.; Santos, M.d.S.; Santana, M.H.A.; Nallakumarasamy, A.; Jeyaraman, M.; Lana, J.V.B.; et al. Intra-Articular Hyaluronic Acid in Osteoarthritis and Tendinopathies: Molecular and Clinical Approaches. Biomedicines 2023, 11, 1061. https://doi.org/10.3390/biomedicines11041061
Costa FR, Costa Marques MR, Costa VC, Santos GS, Martins RA, Santos MdS, Santana MHA, Nallakumarasamy A, Jeyaraman M, Lana JVB, et al. Intra-Articular Hyaluronic Acid in Osteoarthritis and Tendinopathies: Molecular and Clinical Approaches. Biomedicines. 2023; 11(4):1061. https://doi.org/10.3390/biomedicines11041061
Chicago/Turabian StyleCosta, Fabio Ramos, Mariana Ramos Costa Marques, Vinicius Calumby Costa, Gabriel Silva Santos, Rubens Andrade Martins, Marcia da Silva Santos, Maria Helena Andrade Santana, Arulkumar Nallakumarasamy, Madhan Jeyaraman, João Vitor Bizinotto Lana, and et al. 2023. "Intra-Articular Hyaluronic Acid in Osteoarthritis and Tendinopathies: Molecular and Clinical Approaches" Biomedicines 11, no. 4: 1061. https://doi.org/10.3390/biomedicines11041061
APA StyleCosta, F. R., Costa Marques, M. R., Costa, V. C., Santos, G. S., Martins, R. A., Santos, M. d. S., Santana, M. H. A., Nallakumarasamy, A., Jeyaraman, M., Lana, J. V. B., & Lana, J. F. S. D. (2023). Intra-Articular Hyaluronic Acid in Osteoarthritis and Tendinopathies: Molecular and Clinical Approaches. Biomedicines, 11(4), 1061. https://doi.org/10.3390/biomedicines11041061