
Photodynamic Activation of Cholecystokinin 1 Receptor Is Conserved in Mammalian and Avian Pancreatic Acini

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of Rat, Mouse, and Peking Duck Pancreatic Acini
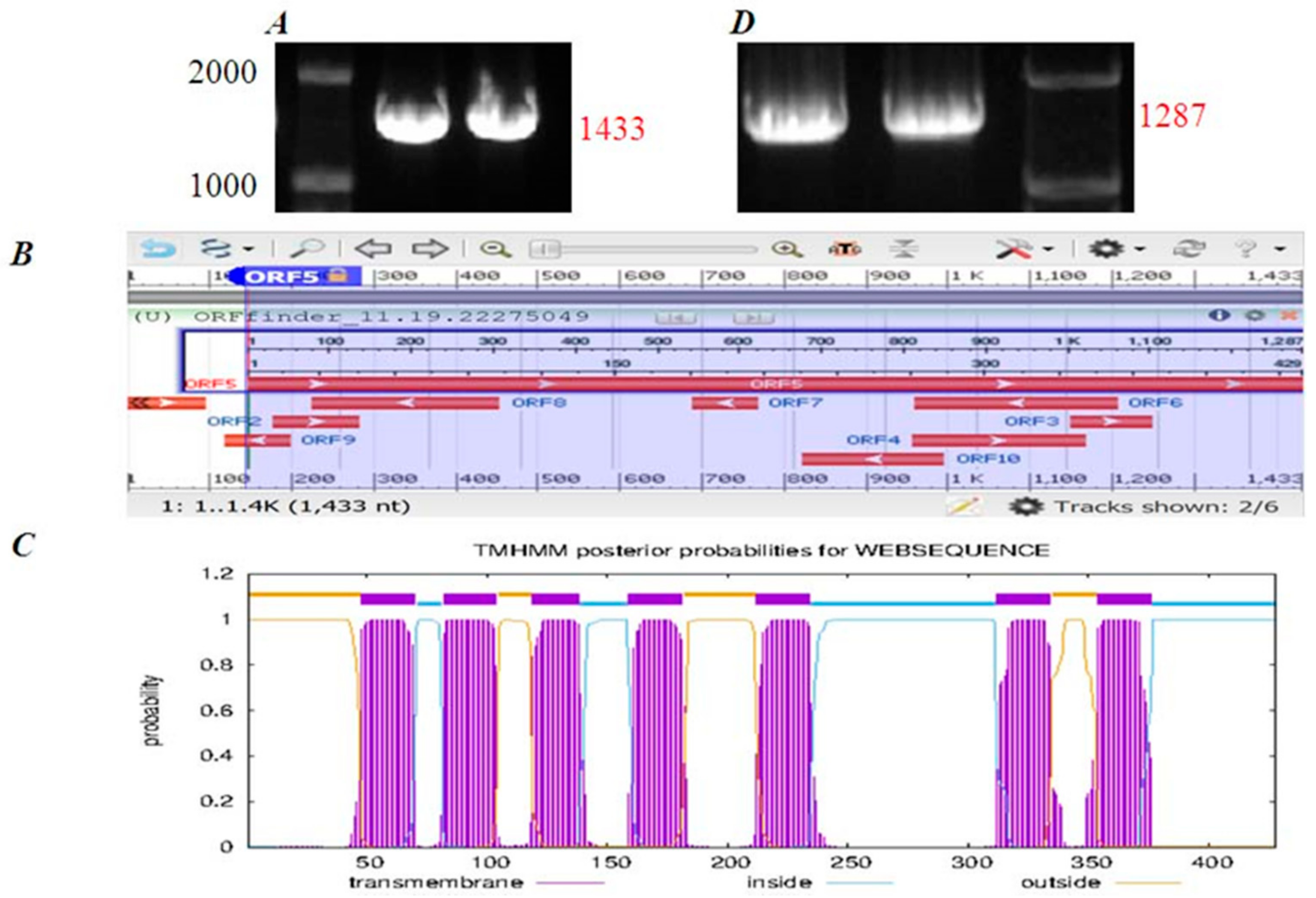
2.3. The Cloning of CCK1R Gene from Peking Duck Pancreatic Acini
2.4. Immunocytochemistry
2.5. Photodynamic Treatment
2.6. Calcium Imaging
2.7. Statistical Analysis
3. Results
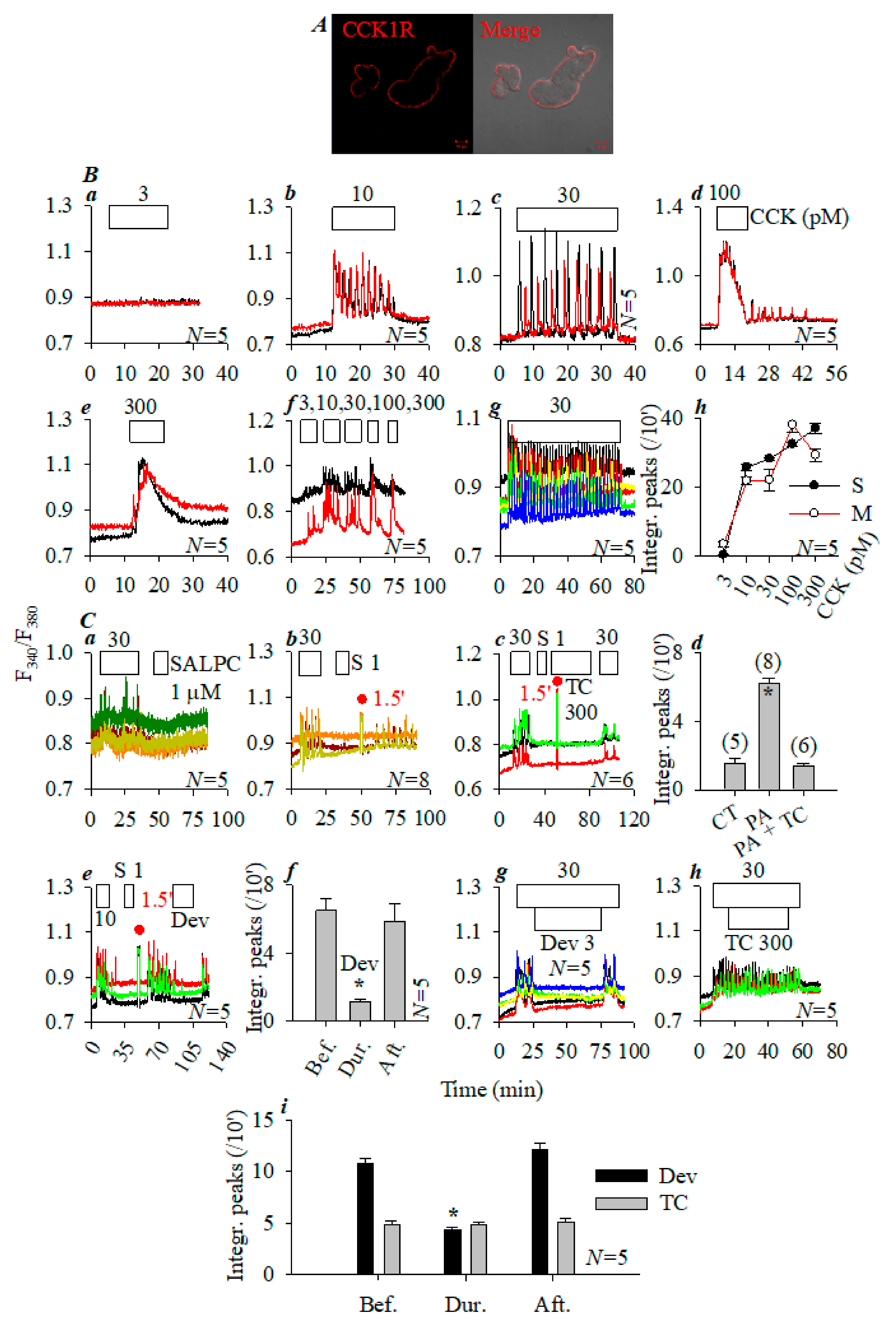
3.1. Photodynamic CCK1R Activation in Rat Pancreatic Acini
3.2. Photodynamic CCK1R Activation in Mouse Pancreatic Acini
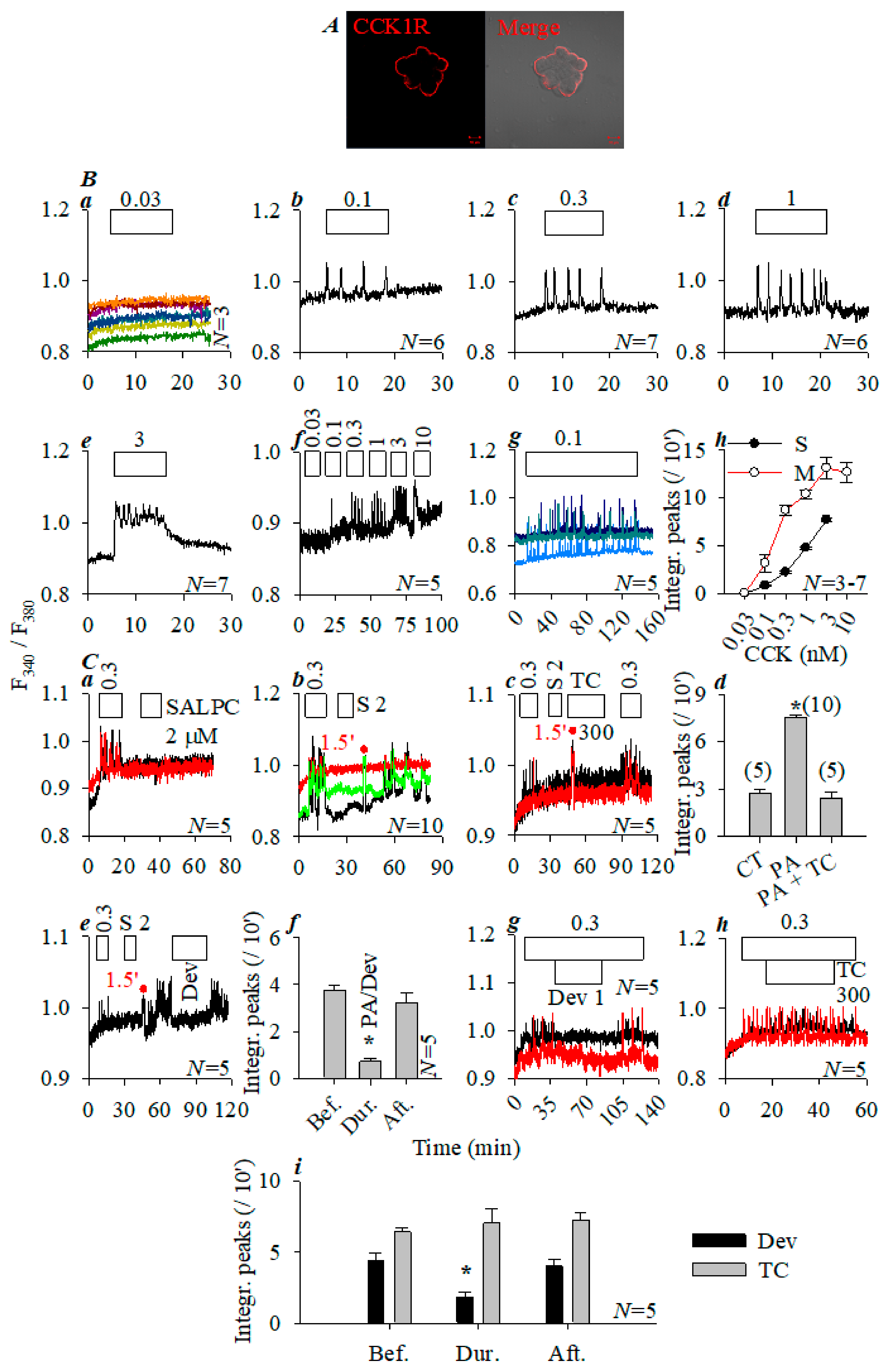
3.3. Photodynamic CCK1R Activation in Peking Duck Pancreatic Acini
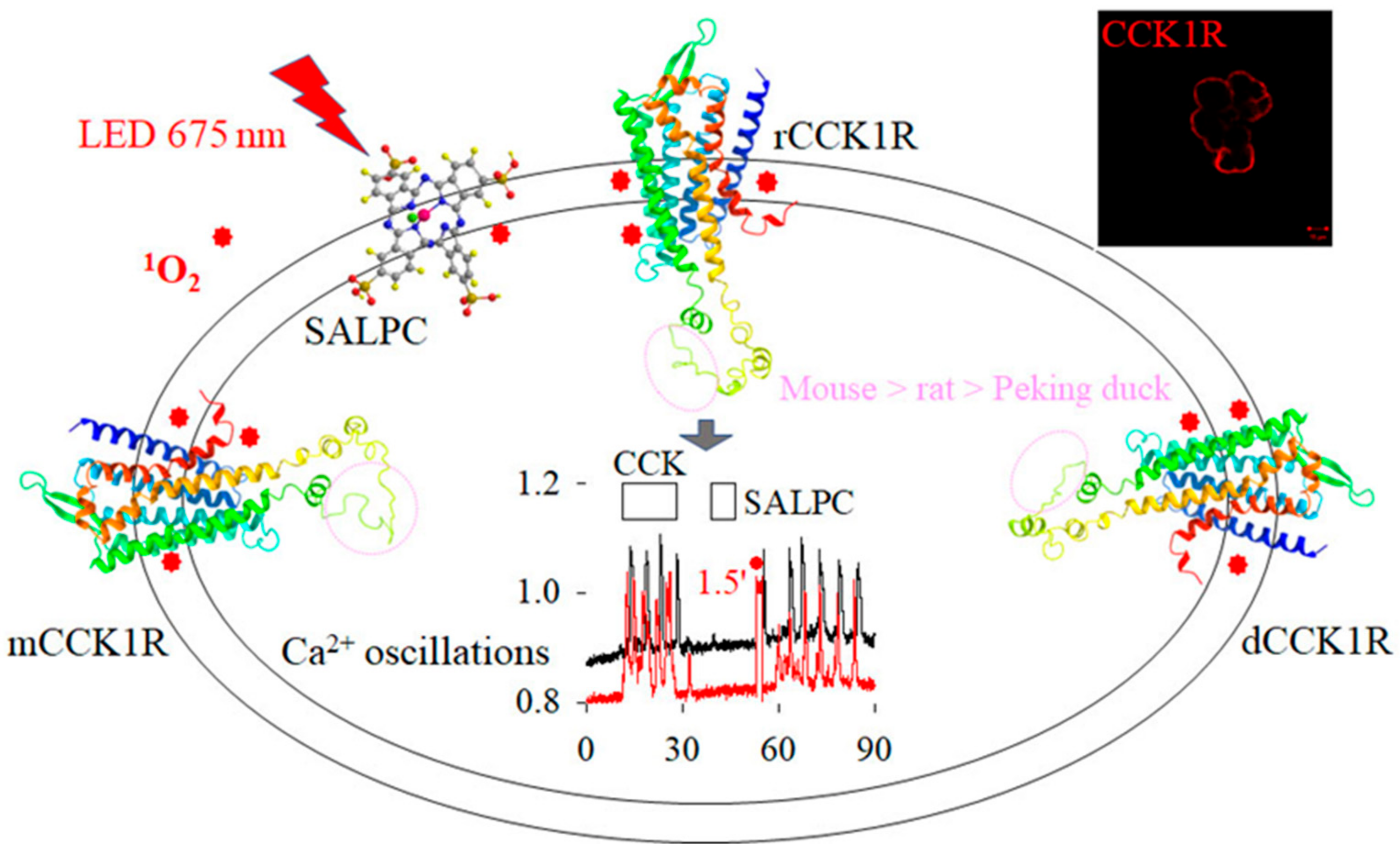
3.4. Comparison of Photodynamic Activation of Rat, Mouse, and Peking Duck Pancreatic Acinar Cell CCK1R
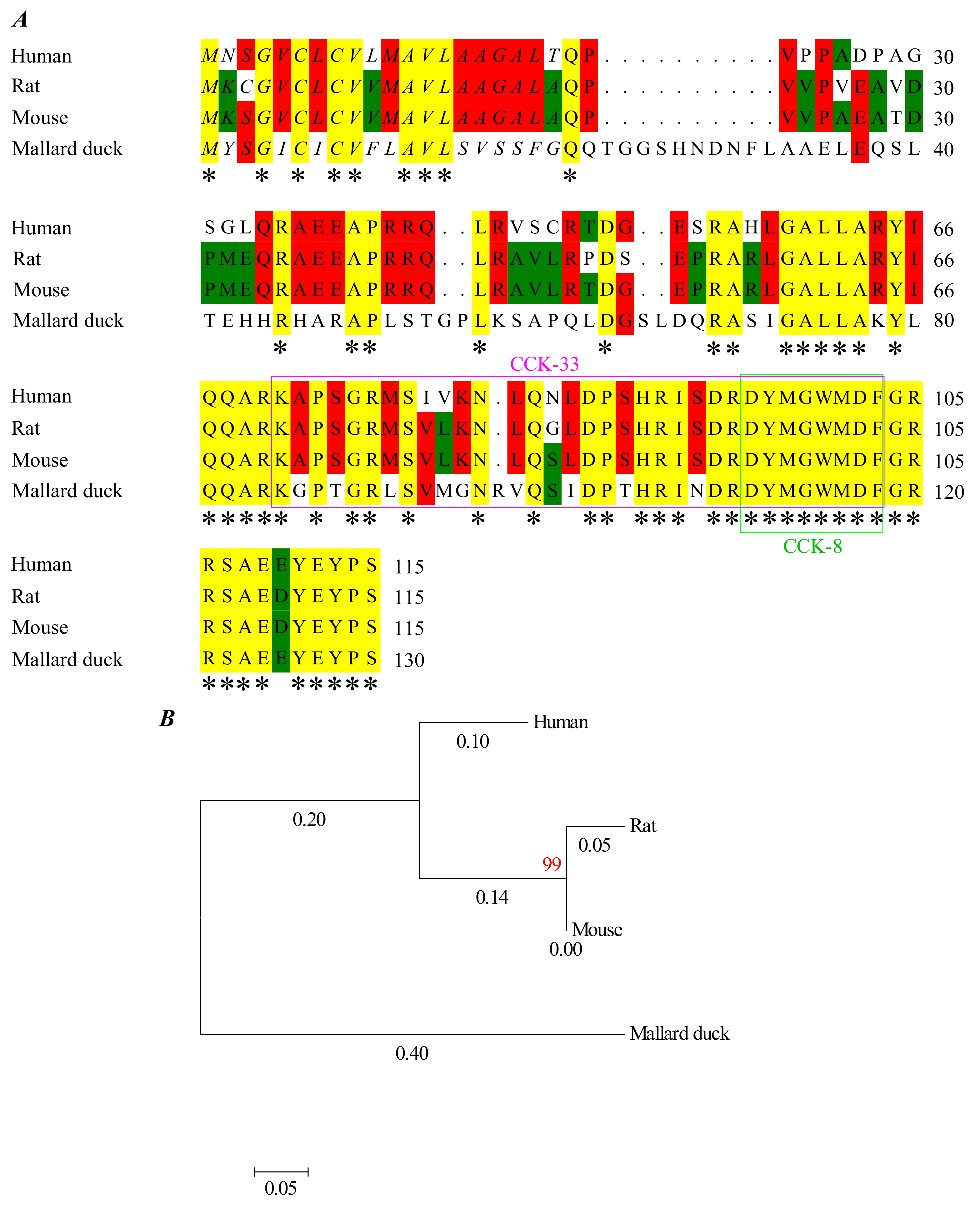
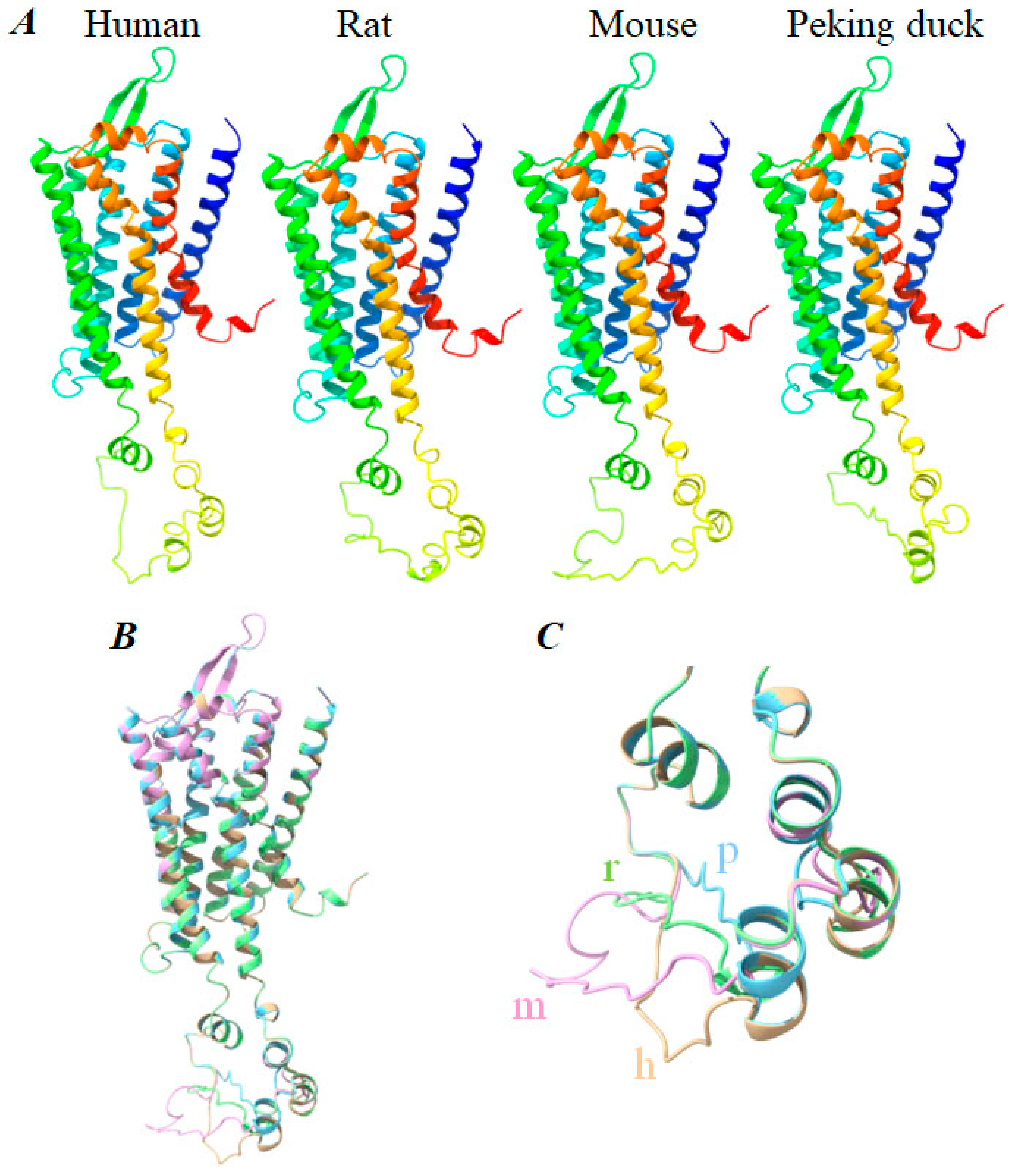
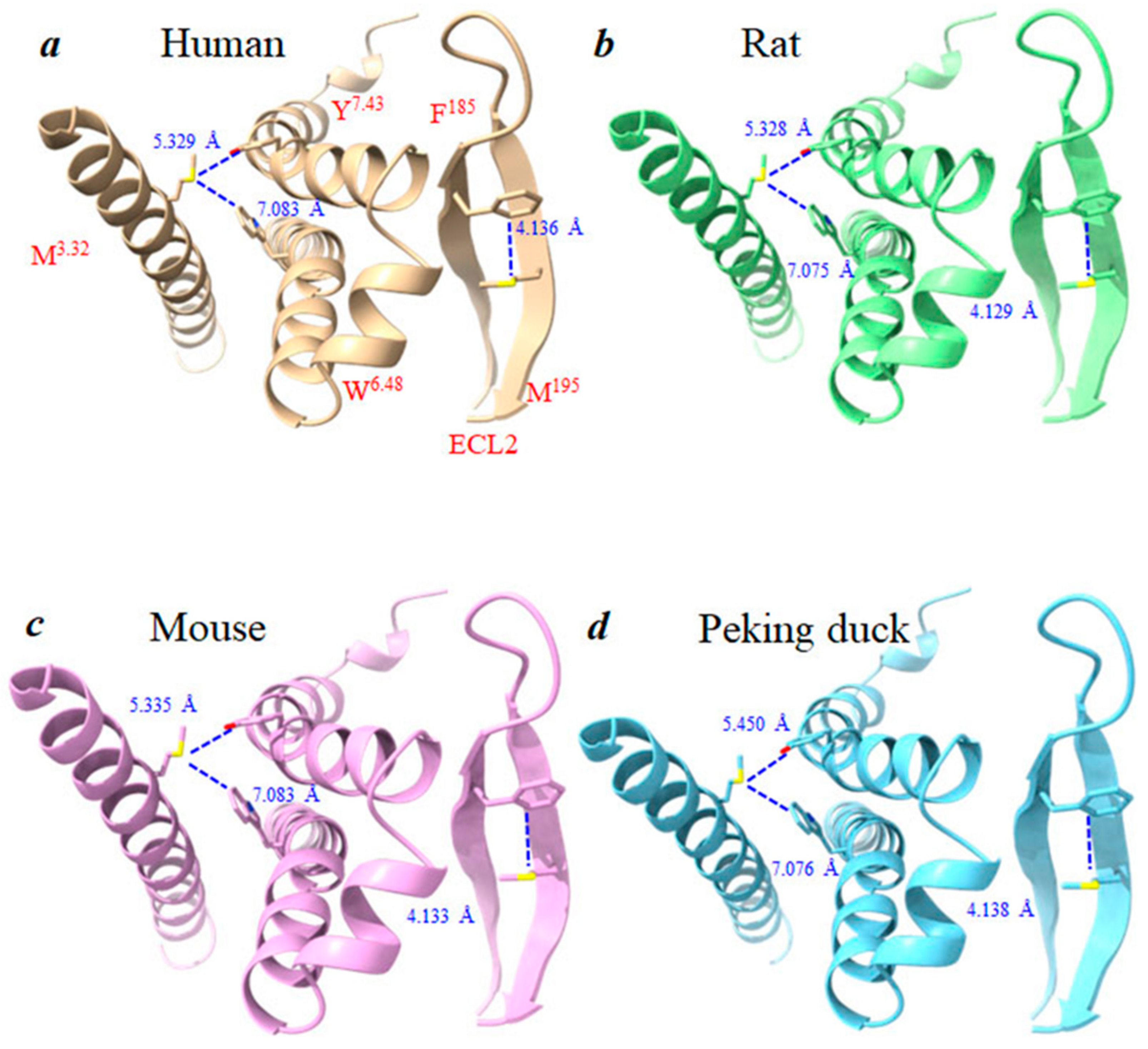
3.5. Structural Basis of Photodynamic Activation of Rat, Mouse, and Peking Duck CCK1R

4. Discussion
4.1. Evolution of CCK-Stimulated Activation of CCK1R
4.2. Conserved Photodynamic Activation of CCK1R
4.3. Better Conservation of Photodynamic Activation of CCK1R
4.4. Conserved Photodynamic Activation Motif “YFM” and Met-Aromatic Interactions
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alexander, S.P.; Kelly, E.; Mathie, A.; Peters, J.A.; Veale, E.L.; Armstrong, J.F.; Faccenda, E.; Harding, S.D.; Pawson, A.J.; Southan, C.; et al. The concise guide to pharmacology 2021/22: Introduction and other protein targets. Br. J. Pharmacol. 2021, 178 (Suppl. S1), S1–S26. [Google Scholar] [CrossRef] [PubMed]
- Bradley, S.J.; Tobin, A.B. Design of next-generation G protein-coupled receptor drugs: Linking novel pharmacology and in vivo animal models. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 535–559. [Google Scholar] [CrossRef] [PubMed]
- Van der Westhuizen, E.T.; Choy, K.H.C.; Valant, C.; McKenzie-Nickson, S.; Bradley, S.J.; Tobin, A.B.; Sexton, P.M.; Christopoulos, A. Fine tuning muscarinic acetylcholine receptor signaling through allostery and bias. Front. Pharmacol. 2021, 11, 606656. [Google Scholar] [CrossRef]
- Dengler, D.G.; Sun, Q.; Harikumar, K.G.; Miller, L.J.; Sergienko, E.A. Screening for positive allosteric modulators of cholecystokinin type 1 receptor potentially useful for management of obesity. SLAS Discov. 2022, 27, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.Y.; Song, Z.M.; Cui, Z.J. Lasting inhibition of receptor-mediated calcium oscillations in pancreatic acini by neutrophil respiratory burst—A novel mechanism for secretory blockade in acute pancreatitis? Biochem. Biophys. Res. Commun. 2013, 437, 361–367. [Google Scholar] [CrossRef]
- Guo, H.Y.; Cui, Z.J. Extracellular histones activate plasma membrane Toll-like receptor 9 to trigger calcium oscillations in rat pancreatic acinar tumor cell AR4-2J. Cells 2018, 8, 3. [Google Scholar] [CrossRef]
- Tran, Q.T.; Sendler, M.; Wiese, M.L.; Doller, J.; Zierke, L.; Gischke, M.; Glaubitz, J.; Tran, V.H.; Lalk, M.; Bornscheuer, U.T.; et al. Systemic bile acids affect the severity of acute pancreatitis in mice depending on their hydrophobicity and the disease pathogenesis. Int. J. Mol. Sci. 2022, 23, 13592. [Google Scholar] [CrossRef]
- Desai, A.J.; Miller, L.J. Sensitivity of cholecystokinin receptors to membrane cholesterol content. Front. Endocrinol. 2012, 3, 123. [Google Scholar] [CrossRef]
- Geiger, J.; Sexton, R.; Al-Sahouri, Z.; Lee, M.Y.; Chun, E.; Harikumar, K.G.; Miller, L.J.; Beckstein, O.; Liu, W. Evidence that specific interactions play a role in the cholesterol sensitivity of G protein-coupled receptors. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183557. [Google Scholar] [CrossRef]
- Cui, Z.J.; Kanno, T. Photodynamic triggering of calcium oscillation in the isolated rat pancreatic acini. J. Physiol. 1997, 504, 47–55. [Google Scholar] [CrossRef]
- Cui, Z.J.; Habara, Y.; Wang, D.Y.; Kanno, T. A novel aspect of photodynamic action: Induction of recurrent spikes in cytosolic calcium concentration. Photochem. Photobiol. 1997, 65, 382–386. [Google Scholar] [CrossRef] [PubMed]
- An, Y.P.; Xiao, R.; Cui, H.; Cui, Z.J. Selective activation by photodynamic action of cholecystokinin receptor in the freshly isolated rat pancreatic acini. Br. J. Pharmacol. 2003, 139, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.N.; Li, Y.; Jiang, W.Y.; Cui, Z.J. Cholecystokinin 1 receptor—A unique G protein-coupled receptor activated by singlet oxygen (GPCR-ABSO). Front. Physiol. 2018, 9, 497. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cui, Z.J. Photodynamic activation of the cholecystokinin 1 receptor with tagged genetically encoded protein photosensitizers: Optimizing the tagging patterns. Photochem. Photobiol. 2022, 98, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cui, Z.J. Transmembrane domain 3 is a transplantable pharmacophore in the photodynamic activation of cholecystokinin 1 receptor. ACS Pharmacol. Transl. Sci. 2022, 5, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Galardy, R.E.; Hull, B.E.; Jamieson, J.D. Irreversible photoactivation of a pancreatic secretagogue receptor with cholecystokinin COOH-terminal octapeptides. J. Biol. Chem. 1980, 255, 3148–3155. [Google Scholar] [CrossRef]
- Matthews, E.K.; Cui, Z.J. Photodynamic action of rose bengal on isolated rat pancreatic acini: Stimulattion of amylase release. FEBS Lett. 1989, 256, 29–32. [Google Scholar] [CrossRef]
- Matthews, E.K.; Cui, Z.J. Photodynamic action of sulphonated aluminium phthalocyanine (SALPC) on isolated rat pancreatic acini. Biochem. Pharmacol. 1990, 39, 1445–1457. [Google Scholar] [CrossRef]
- Li, Y.; Cui, Z.J. Photogenetical activation of cholecystokinin 1 receptor with different genetically encoded protein photosensitizers and from varied subcellular sites. Biomolecules 2020, 10, 1423. [Google Scholar] [CrossRef]
- Li, Y.; Cui, Z.J. NanoLuc bioluminescence-driven photodynamic activation of cholecystokinin 1 receptor with genetically encoded protein photosensitiser miniSOG. Int. J. Mol. Sci. 2020, 21, 3763. [Google Scholar] [CrossRef]
- Jiang, W.Y.; Li, Y.; Li, Z.Y.; Cui, Z.J. Permanent photodynamic cholecystokinin 1 receptor activation: Dimer-to-monomer conversion. Cell. Mol. Neurobiol. 2018, 38, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.J.; Miller, L.J. Agonist-dependent dissociation of oligomeric complexes G protein-coupled cholecystokinin receptors demonstrated in living cells using bioluminescence resonance energy transfer. J. Biol. Chem. 2001, 276, 48040–48047. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Cui, Z.J. Mutual dependence of VIP/PACAP and CCK receptor signaling for a physiological role in duck exocrine pancreatic secretion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 286, R189–R198. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.Z.; Cui, Z.J. Permanent photodynamic activation of the cholecystokinin 2 receptor. Biomolecules 2020, 10, 236. [Google Scholar] [CrossRef]
- Habara, Y.; Kanno, T. Stimulus-secretion coupling and Ca2+ dynamics in pancreatic acinar cells. Gen. Pharmacol. 1994, 25, 843–850. [Google Scholar] [CrossRef]
- Cui, Z.J.; Guo, L.L. Assessing the physiological concentrations of endogenous substances in situ by induced calcium oscillations in vitro. Case of liver. Acta Pharmacol. Sin. 2002, 23, 27–32. [Google Scholar]
- Escrieut, C.; Gigoux, V.; Archer, E.; Verrier, S.; Maigret, B.; Behrendt, R.; Moroder, L.; Bignon, E.; Silvente-Poirot, S.; Pradayrol, L.; et al. The biologically crucial C terminus of cholecystokinin and the non-peptide agonist SR-146,131 share a common binding site in the human CCK1 receptor. Evidence for a crucial role of Met-121 in the activation process. J. Biol. Chem. 2002, 277, 7546–7555. [Google Scholar] [CrossRef]
- Archer-Lahlou, E.; Tikhonova, I.; Escrieut, C.; Dufresne, M.; Seva, C.; Pradayrol, L.; Moroder, L.; Maigret, B.; Fourmy, D. Modeled structure of a G-protein-coupled receptor: The cholecystokinin-1 receptor. J. Med. Chem. 2005, 48, 180–191. [Google Scholar] [CrossRef]
- Archer-Lahlou, E.; Escrieut, C.; Clerc, P.; Martinez, J.; Moroder, L.; Logsdon, C.; Kopin, A.; Seva, C.; Dufresne, M.; Pradayrol, L.; et al. Molecular mechanism underlying partial and full agonism mediated by the human cholecystokinin-1 receptor. J. Biol. Chem. 2005, 280, 10664–10674. [Google Scholar] [CrossRef]
- Ballesteros, J.A.; Weinstein, H. Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Methods Neurosci. 1995, 25, 366–428. [Google Scholar] [CrossRef]
- Liu, Q.F.; Yang, D.H.; Zhuang, Y.W.; Croll, T.I.; Cai, X.Q.; Dai, A.T.; He, X.H.; Duan, J.; Yin, W.C.; Ye, C.Y.; et al. Ligand recognition and G-protein coupling selectivity of cholecystokinin A receptor. Nat. Chem. Biol. 2021, 17, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Yang, D.; Wu, M.; Guo, Y.; Guo, W.; Zhong, L.; Cai, X.; Dai, A.; Jang, W.; Shakhnovich, E.I.; et al. Common activation mechanism of class, A GPCRs. eLife 2019, 8, e50279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; He, C.L.; Wang, M.; Zhou, Q.T.; Yang, D.H.; Zhu, Y.; Feng, W.B.; Zhang, H.; Dai, A.T.; Chu, X.J.; et al. Structures of the human cholecystokinin receptors bound to agonists and antagonists. Nat. Chem. Biol. 2021, 17, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Mobbs, J.I.; Belousoff, M.J.; Harikumar, K.G.; Piper, S.J.; Xu, X.; Furness, S.G.B.; Venugopal, H.; Christopoulos, A.; Danev, R.; Wootten, D.; et al. Structures of the human cholecystokinin 1 (CCK1) receptor bound to Gs and Gq mimetic proteins provide insight into mechanisms of G protein selectivity. PLoS Biol. 2021, 19, e3001295. [Google Scholar] [CrossRef]
- Klotz, L.O.; Kröncke, K.D.; Sies, H. Singlet oxygen-induced signaling effects in mammalian cells. Photochem. Photobiol. Sci. 2003, 2, 88–94. [Google Scholar] [CrossRef]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef]
- Bourassa, J.; Lainé, J.; Kruse, M.L.; Gagnon, M.C.; Calvo, E.; Morisset, J. Ontogeny and species differences in the pancreatic expression and localization of the CCKA receptor. Biochem. Biophys. Res. Commun. 1999, 260, 820–828. [Google Scholar] [CrossRef]
- Konno, K.; Takahashi-Iwanaga, H.; Uchigashima, M.; Miyasaka, K.; Funakoshi, A.; Watanabe, M.; Iwanaga, T. Cellular and subcellular localization of cholecystokinin (CCK)-1 receptors in the pancreas, gallbladder, and stomach of mice. Histochem. Cell Biol. 2015, 143, 301–312. [Google Scholar] [CrossRef]
- Rogers, J.; Hughes, R.G.; Matthews, E.K. Cyclic GMP inhibits protein kinase C-mediated secretion in rat pancreatic acini. J. Biol. Chem. 1988, 263, 3713–3719. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.J.; Han, Z.Q.; Li, Z.Y. Modulating protein activity and cellular function by methionine residue oxidation. Amino Acids 2012, 43, 505–517. [Google Scholar] [CrossRef]
- Liu, J.S.; Cui, Z.J. Pancreatic stellate cells serve as a brake mechanism on pancreatic acinar cell calcium signaling modulated by methionine sulfoxide reductase expression. Cells 2019, 8, 109. [Google Scholar] [CrossRef]
- Josefsen, L.B.; Boyle, R.W. Unique diagnostic and therapeutic roles of porphyrins and phthalocyanines in photodynamic therapy, imaging and theranostics. Theranostics 2012, 2, 916–966. [Google Scholar] [CrossRef] [PubMed]
- Lan, M.; Zhao, S.; Liu, W.; Lee, C.; Zhang, W.; Wang, P. Photosensitizers for photodynamic therapy. Adv. Healthc. Mater. 2019, 8, e1900132. [Google Scholar] [CrossRef]
- Allison, R.R.; Sibata, C.H. Oncologic photodynamic therapy photosensitizers: A clinical review. Photodiagnosis Photodyn. Ther. 2010, 7, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Kitasaka, S.; Yagi, M.; Kikuchi, A. Suppression of menthyl anthranilate (UV-A sunscreen)-sensitized singlet oxygen generation by Trolox and α-tocopherol. Photochem. Photobiol. Sci. 2020, 19, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Giordano, M.E.; Caricato, R.; Lionetto, M.G. Concentration dependence of the antioxidant and prooxidant activity of Trolox in HeLa cells: Involvement in the induction of apoptotic volume decrease. Antioxidants 2020, 9, 1058. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.S.; Wang, Q.; Han, J.Z.; Goswamee, P.; Gupta, A.; McQuiston, A.R.; Liu, Q.L.; Zhou, L. Photodynamic modification of native HCN channels expressed in thalamocortical neurons. ACS Chem. Neurosci. 2020, 11, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Valley, C.C.; Cembran, A.; Perlmutter, J.D.; Lewis, A.K.; Labello, N.P.; Gao, J.; Sachs, J.N. The methionine-aromatic motif plays a unique role in stabilizing protein structure. J. Biol. Chem. 2012, 287, 34979–34991. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.S.; Warren, J.J. A survey of methionine-aromatic interaction geometries in the oxidoreductase class of enzymes: What could Met-aromatic interactions be doing near metal sites? J. Inorg. Biochem. 2018, 186, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Leemann, S.; Kleinlogel, S. Functional optimization of light-activatable opto-GPCRs: Illiminating the importance of the proximal C-terminus in G-protein specificity. bioRxiv 2023. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | CCK Conc. Ratio | Photodynamic Intensity Ratio | ||
|---|---|---|---|---|
| Rat | 30 pM | 3 | SALPC 1 µM; Light 60 mW.cm−2 | 1.2 |
| Mouse | 10 pM | 1 | SALPC 1 µM; Light 50 mW.cm−2 | 1 |
| Peking duck | 300 pM | 30 | SALPC 2 µM; Light 40 mW.cm−2 | 1.6 |
| Residues for | Human | Rat | Mouse | Peking Duck |
|---|---|---|---|---|
| CCK recognition | E344/ECL3 | E360/ECL3 | E352/ECL3 | D346/ECL3 |
| CCK binding | W39/1.30 Q40/1.31 H207/5.39 L356/7.39 | W39/1.30 Q40/1.31 Q222/5.39 L372/7.39 | W39/1.30 Q40/1.31 Q207/5.39 L364/7.39 | L39/1.30 H40/1.31 Y210/5.39 H358/7.39 |
| Receptor activation | Completely identical | |||
| G protein coupling | I296/ICL3 | I312/ICL3 | I304/ICL3 | A298/ICL3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Cui, Z.J. Photodynamic Activation of Cholecystokinin 1 Receptor Is Conserved in Mammalian and Avian Pancreatic Acini. Biomedicines 2023, 11, 885. https://doi.org/10.3390/biomedicines11030885
Wang J, Cui ZJ. Photodynamic Activation of Cholecystokinin 1 Receptor Is Conserved in Mammalian and Avian Pancreatic Acini. Biomedicines. 2023; 11(3):885. https://doi.org/10.3390/biomedicines11030885
Chicago/Turabian StyleWang, Jie, and Zong Jie Cui. 2023. "Photodynamic Activation of Cholecystokinin 1 Receptor Is Conserved in Mammalian and Avian Pancreatic Acini" Biomedicines 11, no. 3: 885. https://doi.org/10.3390/biomedicines11030885
APA StyleWang, J., & Cui, Z. J. (2023). Photodynamic Activation of Cholecystokinin 1 Receptor Is Conserved in Mammalian and Avian Pancreatic Acini. Biomedicines, 11(3), 885. https://doi.org/10.3390/biomedicines11030885