
Involvement of Gut Microbiota in SLE and Lupus Nephritis


Abstract
1. Introduction
2. SLE and LN Pathogenesis
3. Gut Microbiota Dysbiosis of SLE
3.1. Dietary Habits
3.2. Gender
3.3. Geographical Factor
3.4. Concomitant Medication
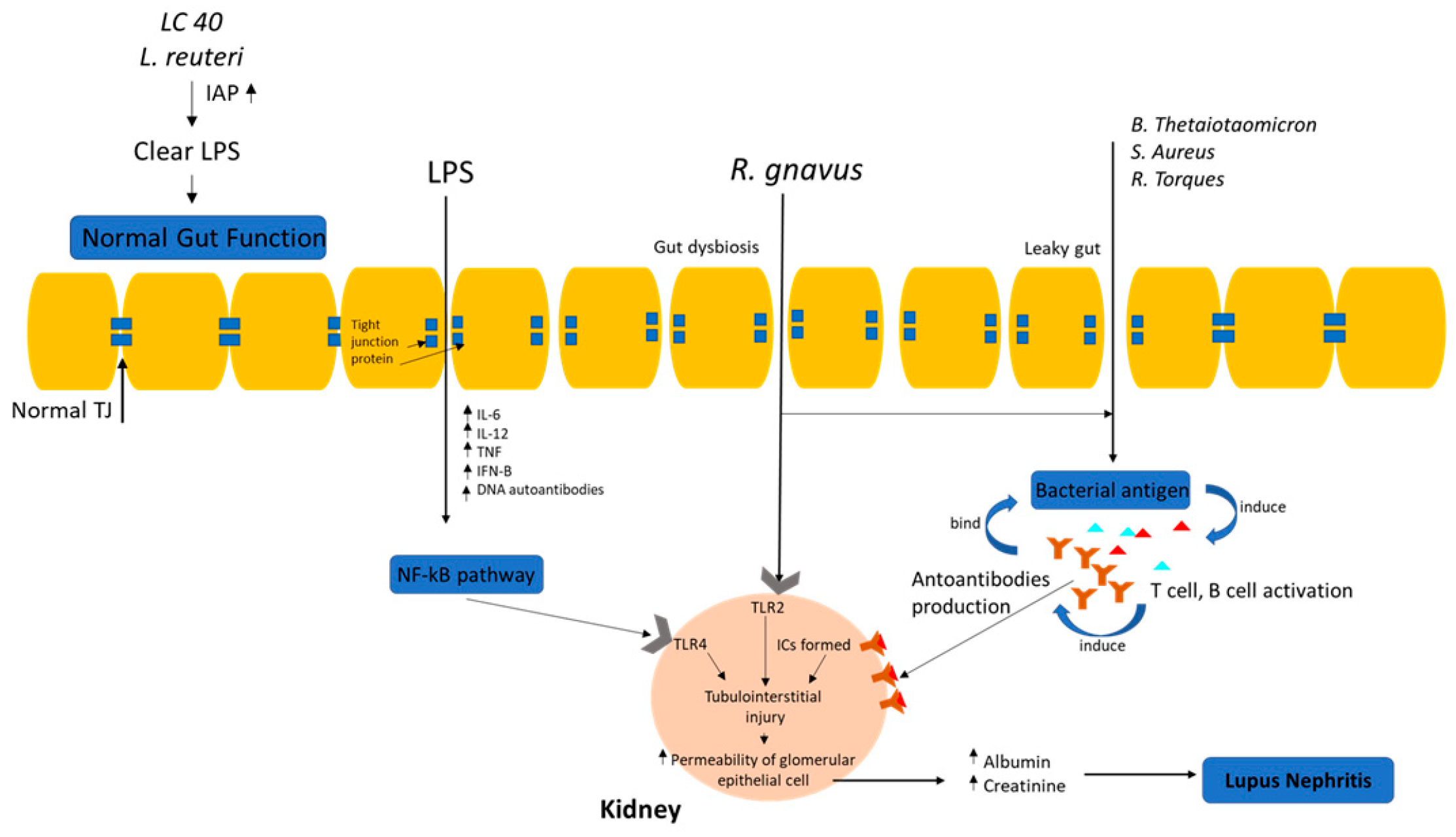
4. Potential Initiation of LN by Gut Microbiota
4.1. Microbes Metabolite Host Axis
4.2. Sex Microbes Axis
5. Clinical Significance of Gut Microbiota in Lupus Nephritis
5.1. Leaky Gut in Lupus Nephritis
5.2. Molecular Mimicry
5.3. SFB Colonization
5.4. Th17/Treg Balance
5.5. Role of IgA
6. Potential Treatment for Lupus Nephritis
6.1. Probiotic Treatment
6.2. Tuftsin-Phosphorylcholine (TPC) Treatment
6.3. Butyrate Supplementary Treatment
6.4. Treatment for Leaky Gut Symptom for IBD

{kind=link}
| Study Types | Method | Study Group | Results | Reference |
|---|---|---|---|---|
| Animal Experimental Study | Chronic oral administration of Probiotic | SLE Mice | Decrease in renal oxidative stress and inflammation Reduced plasma levels of autoantibodies and LPS. | [69] |
| Animal Experimental Study | Tuftsin-phosphorylcholine treatment | SLE Mice | Increases CD1d expression level and reduces CD86 expression level Reduce Akkermansia muciniphila, genera Clostridium, Anaerostipes, and Anaerotruncus Increase Bifidobacterium and Adlercreutzia | [77] |
| Animal Experimental Study | Short-term FMT treatment | SLE Mice | Suppress lupus progression (reduce anti-double-stranded DNA, increase interferon-alpha) | [74] |
| Human Study | Dietary Butyrate treatment | RA Patient | Reduce Urea enzyme, liver enzyme | [84] |
| Animal Study | Probiotic treatment | IBD Mice | Increase colonic interleukin-10 expression and mucin protein-2+ cell and claudin-1+ cell numbers expression Reduce tumor necrosis factor alpha and interleukin-1 βeta expression, NF-κB+Iba1+ cell and LPS+Iba1+ cell numbers in the hippocampus | [87] |
7. Challenges and Future Outlook
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rekvig, O.P. Systemic lupus erythematosus: Definitions, contexts, conflicts, enigmas. Front. Immunol. 2018, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.M.P.; Parks, C.G.; Jacobsen, S.; Costenbader, K.H.; Bernatsky, S. The role of environmental exposures and gene–environment interactions in the etiology of systemic lupus erythematous. J. Intern. Med. 2022, 291, 755–778. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.; Aranow, C.; Mackay, M. Lupus nephritis: Challenges and progress. Curr. Opin. Rheumatol. 2019, 31, 682–688. [Google Scholar] [CrossRef]
- Mu, Q.; Zhang, H.; Liao, X.; Lin, K.; Liu, H.; Edwards, M.R.; Ahmed, S.A.; Yuan, R.; Li, L.; Cecere, T.E.; et al. Control of lupus nephritis by changes of gut microbiota. Microbiome 2017, 5, 73. [Google Scholar] [CrossRef]
- Shaharir, S.S.; Heng Ting, W.M.; Wahab, A.A.; Rajalingham, S.; Abdul Gafor, A.H.; Mohamed Said, M.S.; Mustafar, R. AB0560 A 10-year review of damage accrual and progression in a multi-ethnic cohort of systemic lupus erythematosus (sle) in malaysia. Ann. Rheum. Dis. 2019, 78, 1742. [Google Scholar]
- Shaharir, S.S.; Hussein, H.; Rajalingham, S.; Said, M.S.M.; Gafor, A.H.A.; Mohd, R.; Mustafar, R. Damage in the multiethnic Malaysian systemic lupus erythematosus (SLE) cohort: Comparison with other cohorts worldwide. PLoS ONE 2016, 11, e0166270. [Google Scholar] [CrossRef]
- Jasmin, R.; Sockalingam, S.; Cheah, T.E.; Goh, K.J. Systemic lupus erythematosus in the multiethnic Malaysian population: Disease expression and ethnic differences revisited. Lupus 2013, 22, 967–971. [Google Scholar] [CrossRef]
- Chen, B.; Jia, X.; Xu, J.; Zhao, L.; Ji, J.; Wu, B.; Ma, Y.; Li, H.; Zuo, X.; Pan, W.; et al. An Autoimmunogenic and Proinflammatory Profile Defined by the Gut Microbiota of Patients with Untreated Systemic Lupus Erythematosus. Arthritis Rheumatol. 2021, 73, 232–243. [Google Scholar] [CrossRef]
- Wang, H.; Wang, G.; Banerjee, N.; Liang, Y.; Du, X.; Boor, P.J.; Hoffman, K.L.; Khan, M.F. Aberrant Gut Microbiome Contributes to Intestinal Oxidative Stress, Barrier Dysfunction, Inflammation and Systemic Autoimmune Responses in MRL/lpr Mice. Front. Immunol. 2021, 12, 651191. [Google Scholar] [CrossRef]
- Bagavant, H.; Araszkiewicz, A.M.; Ingram, J.K.; Cizio, K.; Merrill, J.T.; Arriens, C.; Guthridge, J.M.; James, J.A.; Deshmukh, U.S. Immune Response to Enterococcus gallinarum in Lupus Patients Is Associated with a Subset of Lupus-Associated Autoantibodies. Front. Immunol. 2021, 12, 635072. [Google Scholar] [CrossRef]
- Battaglia, M.; Garrett-Sinha, L.A. Bacterial infections in lupus: Roles in promoting immune activation and in pathogenesis of the disease. J. Transl. Autoimmun. 2021, 4, 100078. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Abele, G.; Miggiano, D.; Gasbarrini, A.; Mele, M.C. microorganisms What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Rahbar Saadat, Y.; Hejazian, M.; Bastami, M.; Hosseinian Khatibi, S.M.; Ardalan, M.; Zununi Vahed, S. The role of microbiota in the pathogenesis of lupus: Dose it impact lupus nephritis? Pharmacol. Res. 2019, 139, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Valiente, G.R.; Munir, A.; Hart, M.L.; Blough, P.; Wada, T.T.; Dalan, E.E.; Willis, W.L.; Wu, L.C.; Freud, A.G.; Jarjour, W.N. Gut dysbiosis is associated with acceleration of lupus nephritis. Sci. Rep. 2022, 12, 152. [Google Scholar] [CrossRef] [PubMed]
- Chi, M.; Ma, K.; Wang, J.; Ding, Z.; Li, Y.; Zhu, S.; Liang, X.; Zhang, Q.; Song, L.; Liu, C. The Immunomodulatory Effect of the Gut Microbiota in Kidney Disease. J. Immunol. Res. 2021, 2021, 5516035. [Google Scholar] [CrossRef]
- López, P.; Sánchez, B.; Margolles, A.; Suárez, A. Intestinal dysbiosis in systemic lupus erythematosus: Cause or consequence? Curr. Opin. Rheumatol. 2016, 28, 515–522. [Google Scholar] [CrossRef]
- Gupta, S.; Kaplan, M.J. Bite of the wolf: Innate immune responses propagate autoimmunity in lupus. J. Clin. Investig. 2021, 131, e144918. [Google Scholar] [CrossRef]
- Mahajan, A.; Herrmann, M.; Muñoz, L.E. Clearance deficiency and cell death pathways: A model for the pathogenesis of SLE. Front. Immunol. 2016, 7, 35. [Google Scholar] [CrossRef]
- Kaewraemruaen, C.; Ritprajak, P.; Hirankarn, N. Dendritic cells as key players in systemic lupus erythematosus. Asian Pacific J. Allergy Immunol. 2020, 38, 225–232. [Google Scholar] [CrossRef]
- Jacob, N.; Stohl, W. Cytokine disturbances in systemic lupus erythematosus. Arthritis Res. Ther. 2011, 13, 228. [Google Scholar] [CrossRef]
- Chen, B.; Sun, L.; Zhang, X. Integration of microbiome and epigenome to decipher the pathogenesis of autoimmune diseases. J. Autoimmun. 2017, 83, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Rosser, E.C.; Mauri, C. A clinical update on the significance of the gut microbiota in systemic autoimmunity. J. Autoimmun. 2016, 74, 85–93. [Google Scholar] [CrossRef]
- He, Z.; Shao, T.; Li, H.; Xie, Z.; Wen, C. Alterations of the gut microbiome in Chinese patients with systemic lupus erythematosus. Gut Pathog. 2016, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Silverman, G.J.; Azzouz, D.F.; Alekseyenko, A.V. Systemic Lupus Erythematosus and dysbiosis in the microbiome: Cause or effect or both? Curr. Opin. Immunol. 2019, 61, 80–85. [Google Scholar] [CrossRef]
- Sprouse, M.L.; Bates, N.A.; Felix, K.M.; Wu, H.J.J. Impact of gut microbiota on gut-distal autoimmunity: A focus on T cells. Immunology 2019, 156, 305–318. [Google Scholar] [CrossRef]
- Vieira, J.R.P.; Rezende, A.T.d.O.; Fernandes, M.R.; da Silva, N.A. Intestinal microbiota and active systemic lupus erythematosus: A systematic review. Adv. Rheumatol. 2021, 61, 42. [Google Scholar] [CrossRef] [PubMed]
- Hevia, A.; Milani, C.; López, P.; Cuervo, A.; Arboleya, S.; Duranti, S.; Turroni, F.; González, S.; Suárez, A.; Gueimonde, M.; et al. Intestinal dysbiosis associated with systemic lupus erythematosus. MBio 2014, 5, e01548-14. [Google Scholar] [CrossRef]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut Dysbiosis is Linked to Hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Xu, H.; Xu, J.; Zhao, H.; Lin, Q.; Zhou, Y.; Nie, Y. Sodium Butyrate Ameliorates Gut Microbiota Dysbiosis in Lupus-Like Mice. Front. Nutr. 2020, 7, 604283. [Google Scholar] [CrossRef]
- Choi, S.C.; Brown, J.; Gong, M.; Ge, Y.; Zadeh, M.; Li, W.; Croker, B.P.; Michailidis, G.; Garrett, T.J.; Mohamadzadeh, M.; et al. Gut microbiota dysbiosis and altered tryptophan catabolism contribute to autoimmunity in lupus-susceptible mice. Sci. Transl. Med. 2020, 12, eaax2220. [Google Scholar] [CrossRef]
- Rodríguez-Carrio, J.; López, P.; Sánchez, B.; González, S.; Gueimonde, M.; Margolles, A.; de los Reyes-Gavilán, C.G.; Suárez, A. Intestinal dysbiosis is associated with altered short-chain fatty acids and serum-free fatty acids in systemic lupus erythematosus. Front. Immunol. 2017, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- De Luca, F.; Shoenfeld, Y. The microbiome in autoimmune diseases. Clin. Exp. Immunol. 2019, 195, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Zegarra-Ruiz, D.F.; El Beidaq, A.; Iñiguez, A.J.; Lubrano Di Ricco, M.; Manfredo Vieira, S.; Ruff, W.E.; Mubiru, D.; Fine, R.L.; Sterpka, J.; Greiling, T.M.; et al. A Diet-Sensitive Commensal Lactobacillus Strain Mediates TLR7-Dependent Systemic Autoimmunity. Cell Host Microbe 2019, 25, 113–127.e6. [Google Scholar] [CrossRef]
- Zhang, Q.; Yin, X.; Wang, H.; Wu, X.; Li, X.; Li, Y.; Zhang, X.; Fu, C.; Li, H.; Qiu, Y. Fecal metabolomics and potential biomarkers for systemic lupus erythematosus. Front. Immunol. 2019, 10, 976. [Google Scholar] [CrossRef] [PubMed]
- Shaharir, S.S.; Kadir, W.D.A.; Nordin, F.; Bakar, F.A.; Ting, M.W.H.; Jamil, A.; Mohd, R.; Wahab, A.A. Systemic lupus erythematosus among male patients in Malaysia: How are we different from other geographical regions? Lupus 2019, 28, 137–144. [Google Scholar] [CrossRef]
- Khan, D.; Ansar Ahmed, S. The immune system is a natural target for estrogen action: Opposing effects of estrogen in two prototypical autoimmune diseases. Front. Immunol. 2016, 6, 635. [Google Scholar] [CrossRef]
- Singh, R.P.; Hahn, B.H.; Bischoff, D.S. Interferon Genes Are Influenced by 17β-Estradiol in SLE. Front. Immunol. 2021, 12, 725325. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.R.; Dai, R.; Heid, B.; Cowan, C.; Werre, S.R.; Cecere, T.; Ahmed, S.A. Low-dose 17α-ethinyl estradiol (EE) exposure exacerbates lupus renal disease and modulates immune responses to TLR7/9 agonists in genetically autoimmune-prone mice. Sci. Rep. 2020, 10, 5210. [Google Scholar] [CrossRef]
- Singh, R.P.; Bischoff, D.S. Sex Hormones and Gender Influence the Expression of Markers of Regulatory T Cells in SLE Patients. Front. Immunol. 2021, 12, 619268. [Google Scholar] [CrossRef] [PubMed]
- Neuman, H.; Koren, O. The gut microbiota: A possible factor influencing systemic lupus erythematosus. Curr. Opin. Rheumatol. 2017, 29, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Schwartzman-Morris, J.; Putterman, C. Gender differences in the pathogenesis and outcome of lupus and of lupus nephritis. Clin. Dev. Immunol. 2012, 2012, 604892. [Google Scholar] [CrossRef]
- Wei, F.; Xu, H.; Yan, C.; Rong, C.; Liu, B.; Zhou, H. Changes of intestinal flora in patients with systemic lupus erythematosus in northeast China. PLoS ONE 2019, 14, e0213063. [Google Scholar] [CrossRef]
- Pan, Q.; Li, Y.; Ye, L.; Deng, Z.; Li, L.; Feng, Y.; Liu, W.; Liu, H. Geographical distribution, a risk factor for the incidence of lupus nephritis in China. BMC Nephrol. 2014, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wang, H.; Xu, S.; Zhuang, Y.; An, J.; Su, C.; Xia, Y.; Chen, J.; Xu, Z.Z.; Liu, Q.; et al. Alteration in gut microbiota is associated with dysregulation of cytokines and glucocorticoid therapy in systemic lupus erythematosus. Gut Microbes 2020, 11, 1758–1773. [Google Scholar] [CrossRef] [PubMed]
- Feldman, C.H.; Hiraki, L.T.; Winkelmayer, W.C.; Marty, F.M.; Franklin, J.M.; Kim, S.C.; Costenbader, K.H. Serious infections among adult medicaid beneficiaries with systemic lupus erythematosus and lupus nephritis. Arthritis Rheumatol. 2015, 67, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Chang, Y.; Han, N.; Hou, F.; Lee, B.J.Y.; Zhi, F. Short-term high-dose gavage of hydroxychloroquine changes gut microbiota but not the intestinal integrity and immunological responses in mice. Life Sci. 2021, 264, 118450. [Google Scholar] [CrossRef]
- Gottschalk, T.; Tsantikos, E.; Hibbs, M. 35 A diet high in fibre diet can moderate inflammation and kidney pathology in a model of systemic lupus erythematosus. Lupus Sci. Med. 2017, 4, A17. [Google Scholar]
- Barbhaiya, M.; Costenbader, K.H. Environmental exposures and the development of systemic lupus erythematosus. Curr. Opin. Rheumatol. 2016, 28, 497–505. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, C.Z.; Wan, J.Y.; Yao, H.; Yuan, C.S. Dissecting the interplay mechanism between epigenetics and gut microbiota: Health maintenance and disease prevention. Int. J. Mol. Sci. 2021, 22, 6933. [Google Scholar] [CrossRef]
- Felizardo, R.J.F.; de Almeida, D.C.; Pereira, R.L.; Watanabe, I.K.M.; Doimo, N.T.S.; Ribeiro, W.R.; Cenedeze, M.A.; Hiyane, M.I.; Amano, M.T.; Braga, T.T.; et al. Gut microbial metabolite butyrate protects against proteinuric kidney disease through epigenetic- and GPR109a-mediated mechanisms. FASEB J. 2019, 33, 11894–11908. [Google Scholar] [CrossRef]
- Ren, J.; Catalina, M.D.; Eden, K.; Liao, X.; Read, K.A.; Luo, X.; McMillan, R.P.; Hulver, M.W.; Jarpe, M.; Bachali, P.; et al. Selective histone deacetylase 6 inhibition normalizes b cell activation and germinal center formation in a model of systemic lupus erythematosus. Front. Immunol. 2019, 10, 2512. [Google Scholar] [CrossRef]
- Li, M.; Jiang, A. DNA methylation was involved in total glucosides of paeony regulating ERα for the treatment of female systemic lupus erythematosus mice. J. Pharmacol. Sci. 2019, 140, 187–192. [Google Scholar] [CrossRef]
- Syrett, C.M.; Sierra, I.; Beethem, Z.T.; Dubin, A.H.; Anguera, M.C. Loss of epigenetic modifications on the inactive X chromosome and sex-biased gene expression profiles in B cells from NZB/W F1 mice with lupus-like disease. J. Autoimmun. 2020, 107, 102357. [Google Scholar] [CrossRef] [PubMed]
- Christou, E.A.A.; Banos, A.; Kosmara, D.; Bertsias, G.K.; Boumpas, D.T. Sexual dimorphism in SLE: Above and beyond sex hormones. Lupus 2019, 28, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liao, X.; Sparks, J.B.; Luo, X.M. Dynamics of gut microbiota in autoimmune lupus. Appl. Environ. Microbiol. 2014, 80, 7551–7560. [Google Scholar] [CrossRef]
- De Carvalho, J.F.; Do Nascimento, A.P.; Testagrossa, L.A.; Barros, R.T.; Bonfá, E. Male gender results in more severe lupus nephritis. Rheumatol. Int. 2010, 30, 1311–1315. [Google Scholar] [CrossRef]
- Guo, S.; Al-Sadi, R.; Said, H.M.; Ma, T.Y. Lipopolysaccharide causes an increase in intestinal tight junction permeability in vitro and in vivo by inducing enterocyte membrane expression and localization of TLR-4 and CD14. Am. J. Pathol. 2013, 182, 375–387. [Google Scholar] [CrossRef]
- Eadon, M.T.; Hack, B.K.; Xu, C.; Ko, B.; Toback, F.G.; Cunningham, P.N. Endotoxemia alters tight junction gene and protein expression in the kidney. Am. J. Physiol. Ren. Physiol. 2012, 303, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, S.C. NF-ΚB in inflammation and renal diseases. Cell Biosci. 2015, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Barnabei, L.; Laplantine, E.; Mbongo, W.; Rieux-Laucat, F.; Weil, R. NF-κB: At the Borders of Autoimmunity and Inflammation. Front. Immunol. 2021, 12, 716469. [Google Scholar] [CrossRef] [PubMed]
- Caster, D.J.; Powell, D.W. Precision Targeting of NF-κB Signaling in Lupus Nephritis. Lupus 2020, 6, 160. [Google Scholar]
- Pawar, R.D.; Castrezana-Lopez, L.; Allam, R.; Kulkarni, O.P.; Segerer, S.; Radomska, E.; Meyer, T.N.; Schwesinger, C.M.; Akis, N.; Gröne, H.J.; et al. Bacterial lipopeptide triggers massive albuminuria in murine lupus nephritis by activating Toll-like receptor 2 at the glomerular filtration barrier. Immunology 2009, 128 (Suppl. 1), e206–e221. [Google Scholar] [CrossRef]
- Fawley, J.; Gourlay, D.M. Intestinal alkaline phosphatase: A summary of its role in clinical disease. J. Surg. Res. 2016, 202, 225–234. [Google Scholar] [CrossRef]
- Azzouz, D.; Omarbekova, A.; Heguy, A.; Schwudke, D.; Gisch, N.; Rovin, B.H.; Caricchio, R.; Buyon, J.P.; Alekseyenko, A.V.; Silverman, G.J. Lupus nephritis is linked to disease-activity associated expansions and immunity to a gut commensal. Ann. Rheum. Dis. 2019, 78, 947–956. [Google Scholar] [CrossRef]
- Azzouz, D.F.; Chen, Z.; Izmirly, P.; Fenyo, D.; Buyon, J.; Alekseyenko, A.V.; Silverman, G.J. 511 Disease flares in lupus are concordant with Ruminococcus Blautia Gnavus blooms arising within unstable gut microbiota communities. Lupus Sci. Med. 2021, 8, A18–A19. [Google Scholar]
- Avni, O.; Koren, O. Molecular (Me)micry? Cell Host Microbe 2018, 23, 576–578. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, M.A.; Gurjar, H.; Sindhu, T.; Haider, A.; Arshad, M. A Curious Case of Acute Glomerulonephritis—Staphylococcus vs. Lupus: Case Report and Literature Review. Cureus 2021, 13, e13138. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Liu, Y.; Liu, F.; Liu, C.; Li, J.; Niu, J.; Han, P.; Xu, D.; Hou, J.; Ma, Y.; et al. A Novel Autoantibody Induced by Bacterial Biofilm Conserved Components Aggravates Lupus Nephritis. Front. Immunol. 2021, 12, 656090. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; Toral, M.; Romero, M.; Sanchez, M.; Jimenez, R.; O’Valle, F.; Rodriguez-Nogales, A.; de la Visitacion, N.; Galvez, J.; Duarte, J. Lactobacillus fermentum CECT5716 consumption protects kidney and improves endothelial dysfunction in murine lupus. Basic Clin. Pharmacol. Toxicol. 2017, 121, 52. [Google Scholar]
- Tezuka, H.; Ohteki, T. Regulation of IgA production by intestinal dendritic cells and related cells. Front. Immunol. 2019, 10, 1891. [Google Scholar] [CrossRef]
- Gong, J.; Noel, S.; Pluznick, J.L.; Hamad, A.R.A.; Rabb, H. Gut Microbiota-Kidney Cross-Talk in Acute Kidney Injury. Semin. Nephrol. 2019, 39, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Hongyan, L.; Yi, Z.; Bao, D.; Yuewu, L.; Juan, M. A study on clinical and pathologic features in lupus nephritis with mainly IgA deposits and a literature review. Clin. Dev. Immunol. 2013, 2013, 289316. [Google Scholar] [CrossRef]
- Esmaeili, S.A.; Mahmoudi, M.; Momtazi, A.A.; Sahebkar, A.; Doulabi, H.; Rastin, M. Tolerogenic probiotics: Potential immunoregulators in Systemic Lupus Erythematosus. J. Cell. Physiol. 2017, 232, 1994–2007. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Q.; Yu, Y.; Wang, M.; Wen, C.; He, Z. Early and Short-Term Interventions in the Gut Microbiota Affects Lupus Severity, Progression, and Treatment in MRL/lpr Mice. Front. Microbiol. 2020, 11, 628. [Google Scholar] [CrossRef] [PubMed]
- Moeller, A.H.; Sanders, J.G. Roles of the gut microbiota in the adaptive evolution of mammalian species. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190597. [Google Scholar] [CrossRef]
- Yang, Y.; Nguyen, M.; Khetrapal, V.; Sonnert, N.D.; Martin, A.L.; Chen, H.; Kriegel, M.A.; Palm, N.W. Within-host evolution of a gut pathobiont facilitates liver translocation. Nature 2022, 607, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Neuman, H.; Mor, H.; Bashi, T.; Givol, O.; Watad, A.; Shemer, A.; Volkov, A.; Barshack, I.; Fridkin, M.; Blank, M.; et al. Helminth-Based Product and the Microbiome of Mice with Lupus. mSystems 2019, 4, e00160-18. [Google Scholar] [CrossRef]
- Bashi, T.; Blank, M.; Ben-Ami Shor, D.; Fridkin, M.; Versini, M.; Gendelman, O.; Volkov, A.; Barshak, I.; Shoenfeld, Y. Successful modulation of murine lupus nephritis with tuftsin-phosphorylcholine. J. Autoimmun. 2015, 59, 1–7. [Google Scholar] [CrossRef]
- Bungau, S.G.; Behl, T.; Singh, A.; Sehgal, A.; Singh, S.; Chigurupati, S.; Vijayabalan, S.; Das, S.; Palanimuthu, V.R. Targeting probiotics in rheumatoid arthritis. Nutrients 2021, 13, 3376. [Google Scholar] [CrossRef]
- He, J.; Chu, Y.; Li, J.; Meng, Q.; Liu, Y.; Jin, J.; Wang, Y.; Wang, J.; Huang, B.; Shi, L.; et al. Intestinal butyrate-metabolizing species contribute to autoantibody production and bone erosion in rheumatoid arthritis. Sci. Adv. 2022, 8, 730–735. [Google Scholar] [CrossRef]
- Gioia, C.; Lucchino, B.; Tarsitano, M.G.; Iannuccelli, C.; Di Franco, M. Dietary habits and nutrition in rheumatoid arthritis: Can diet influence disease development and clinical manifestations? Nutrients 2020, 12, 1456. [Google Scholar] [CrossRef]
- Häger, J.; Bang, H.; Hagen, M.; Frech, M.; Träger, P.; Sokolova, M.V.; Stéen, U.; Tascilar, K.; Sarter, K.; Schett, G.; et al. The role of dietary fiber in rheumatoid arthritis patients: A feasibility study. Nutrients 2019, 11, 2392. [Google Scholar] [CrossRef]
- Dürholz, K.; Hofmann, J.; Iljazovic, A.; Häger, J.; Lucas, S.; Sarter, K.; Strowig, T.; Bang, H.; Rech, J.; Schett, G.; et al. Dietary short-term fiber interventions in arthritis patients increase systemic scfa levels and regulate inflammation. Nutrients 2020, 12, 3207. [Google Scholar] [CrossRef]
- Kamal, E.; Kaddam, L.A.G.; Alagib, A.; Saeed, A. Dietary Fibers (Gum Arabic) Supplementation Modulates Hepatic and Renal Profile Among Rheumatoid Arthritis Patients, Phase II Trial. Front. Nutr. 2021, 8, 552049. [Google Scholar] [CrossRef] [PubMed]
- Pakravan, N.; Kermanian, F.; Mahmoudi, E. Filtered Kombucha tea ameliorates the leaky gut syndrome in young and old mice model of colitis. Iran. J. Basic Med. Sci. 2019, 22, 1158–1165. [Google Scholar] [CrossRef] [PubMed]
- Twardowska, A.; Makaro, A.; Binienda, A.; Fichna, J.; Salaga, M. Preventing Bacterial Translocation in Patients with Leaky Gut Syndrome: Nutrition and Pharmacological Treatment Options. Int. J. Mol. Sci. 2022, 23, 3204. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Joo, M. Tight Junction Protein Expression-Inducing Probiotics Alleviate TNBS-Induced Cognitive Impairment with Colitis in Mice. Nutrients 2022, 14, 2975. [Google Scholar] [CrossRef]
- Li, D.; Pan, Y.; Xia, X.; Liang, J.; Liu, F.; Dou, H.; Hou, Y. Bacteroides fragilis alleviates the symptoms of lupus nephritis via regulating CD1d and CD86 expressions in B cells. Eur. J. Pharmacol. 2020, 884, 173421. [Google Scholar] [CrossRef] [PubMed]
 : Increment.
: Increment.
: Increment.
: Increment.
| Microbial Species | Changes | Study Group | Effects | Reference |
|---|---|---|---|---|
| R. torques spp. | Increase | Mice | Increase of MCP-1, CXCL1 and kidney infiltrating F4/80+CD206+M2-like macrophages | [14] |
| Ruminococcus gnavus | Increase | Human | High anti-RG antibodies, high SLEDAI score, high antinative DNA levels, but low complement 3 and complement 4 level | [64] |
| Staphylococcus aureus | Increase | Human | High anti-HU1 antibodies | [68] |
| Bacteroides thetaiotaomicron | Increase | Mice | High anti-Ro60 antibodies | [66] |
| Lactobacillus fermentum CECT5716 | Increase | Mice | Low anti-double stranded DNA antibodies, T, B, Tregs and T-helper 1 cells | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd, R.; Chin, S.-F.; Shaharir, S.S.; Cham, Q.S. Involvement of Gut Microbiota in SLE and Lupus Nephritis. Biomedicines 2023, 11, 653. https://doi.org/10.3390/biomedicines11030653
Mohd R, Chin S-F, Shaharir SS, Cham QS. Involvement of Gut Microbiota in SLE and Lupus Nephritis. Biomedicines. 2023; 11(3):653. https://doi.org/10.3390/biomedicines11030653
Chicago/Turabian StyleMohd, Rozita, Siok-Fong Chin, Syahrul Sazliyana Shaharir, and Qin Shi Cham. 2023. "Involvement of Gut Microbiota in SLE and Lupus Nephritis" Biomedicines 11, no. 3: 653. https://doi.org/10.3390/biomedicines11030653
APA StyleMohd, R., Chin, S.-F., Shaharir, S. S., & Cham, Q. S. (2023). Involvement of Gut Microbiota in SLE and Lupus Nephritis. Biomedicines, 11(3), 653. https://doi.org/10.3390/biomedicines11030653