
Vitamin D Ameliorates the Hepatic Oxidative Damage and Fibrotic Effect Caused by Thioacetamide in Rats

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals and Experimental Design
2.3. Handling of Biological Samples
2.4. Assessment of Serum Biochemistry Parameters
2.5. Evaluation of Antioxidant and Hepatic Lipid Peroxidation Markers
2.6. Hepatic Gene Expression Analysis
2.7. Liver Histopathology Assessment
2.8. Immunohistochemistry of α-SMA and TGF-β Expression
2.9. Statistical Analysis
3. Results
3.1. Influence of TAA and VD3 on Serum Biochemical Parameters
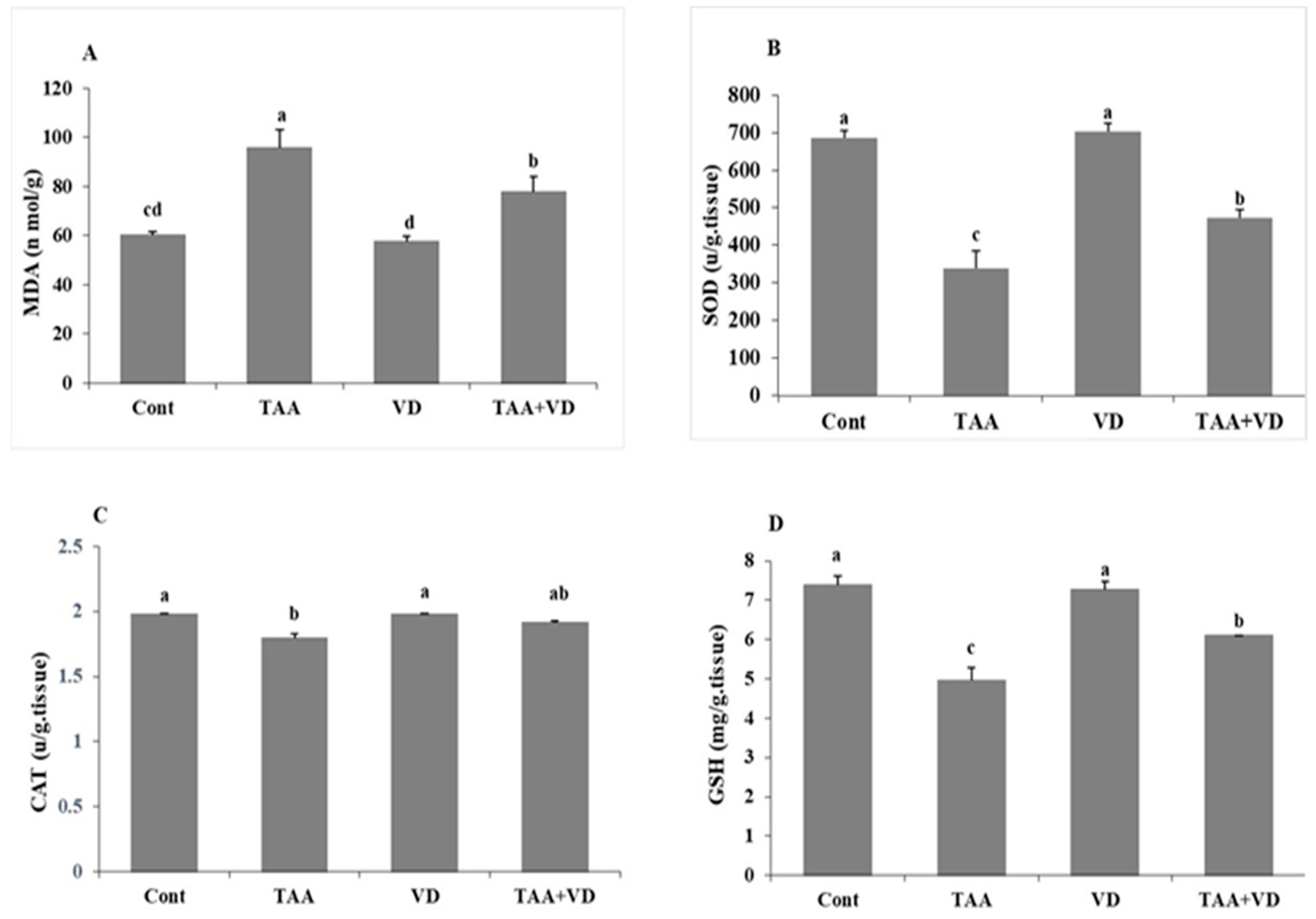
3.2. Influence of TAA and VD3 on Hepatic Lipid Peroxidation/Antioxidant Markers
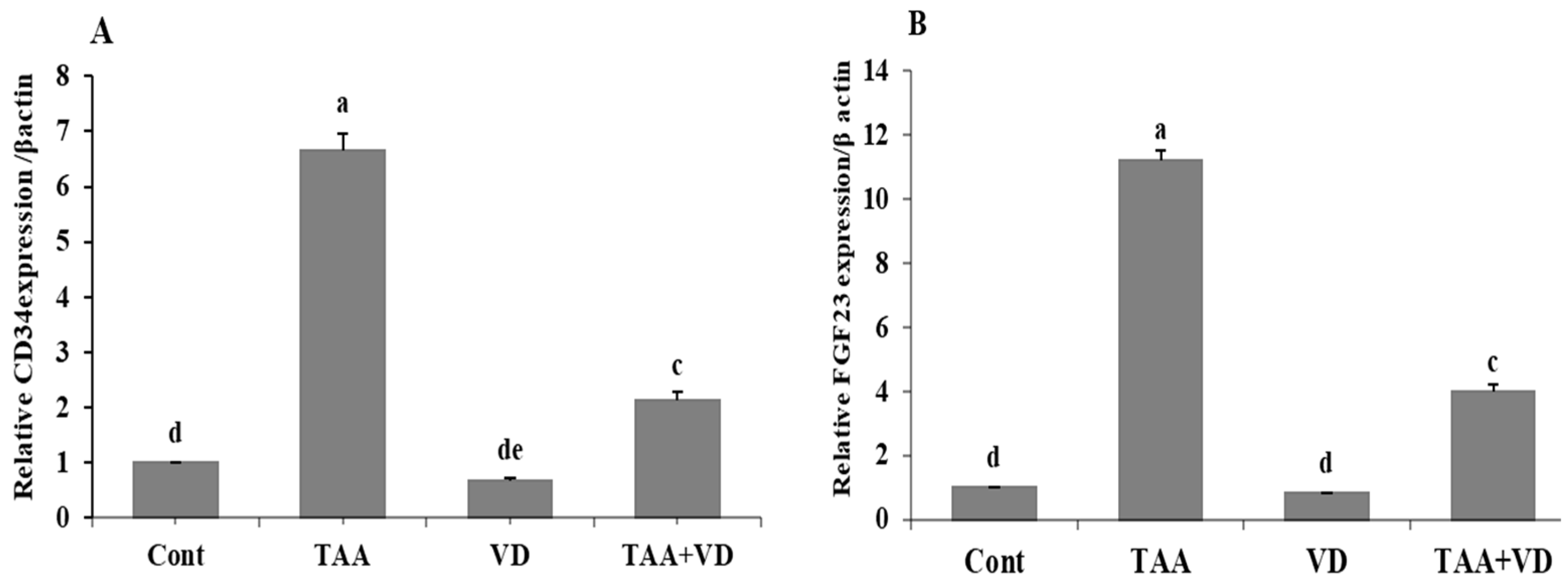
3.3. Influence of TAA and VD3 on Hepatic CD34 and FGF23 Gene Expression
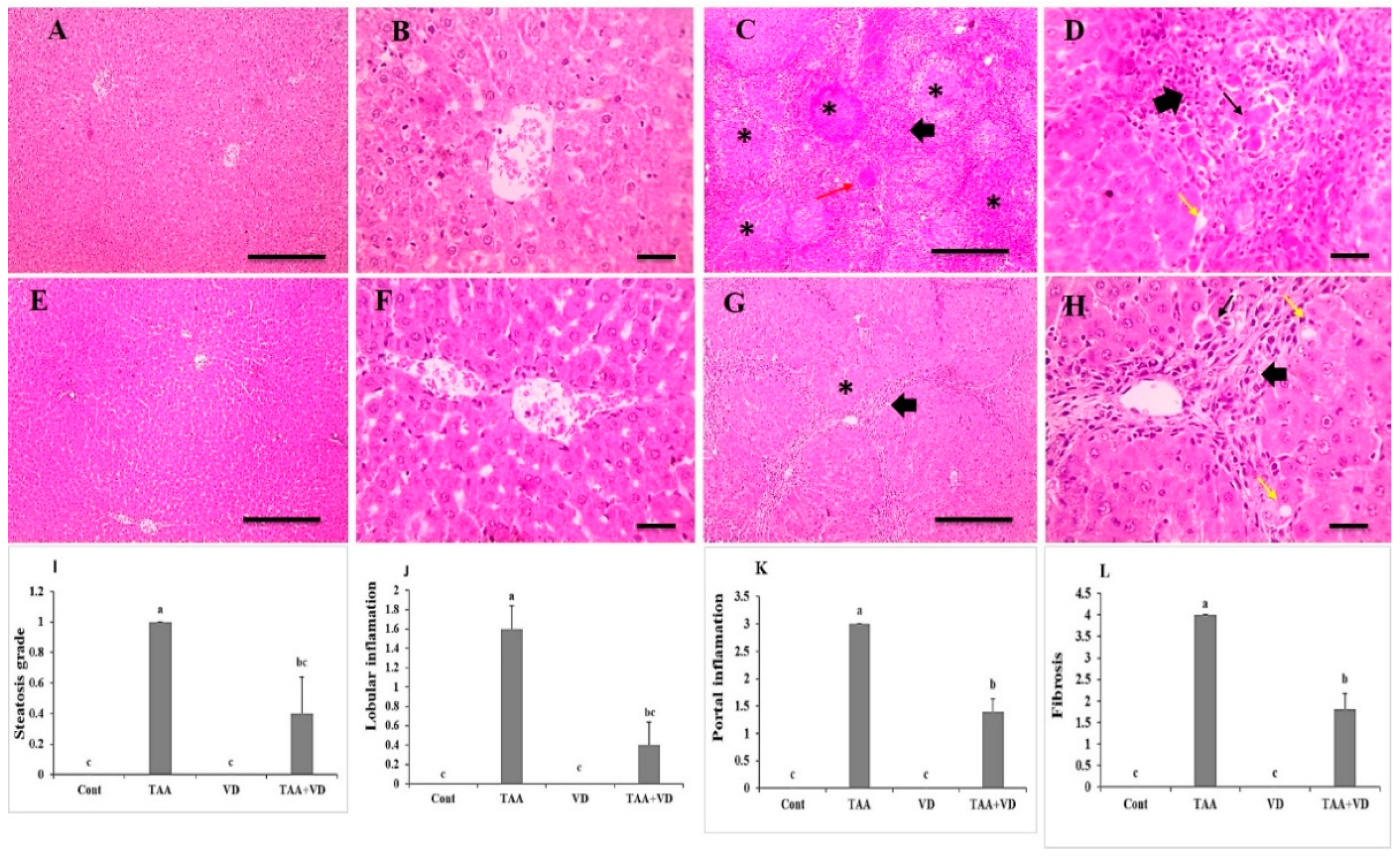
3.4. Histopathological Examination of Liver Sections Using H&E Staining
3.5. Histopathological Examination of Liver Sections Using Masson’s Trichrome
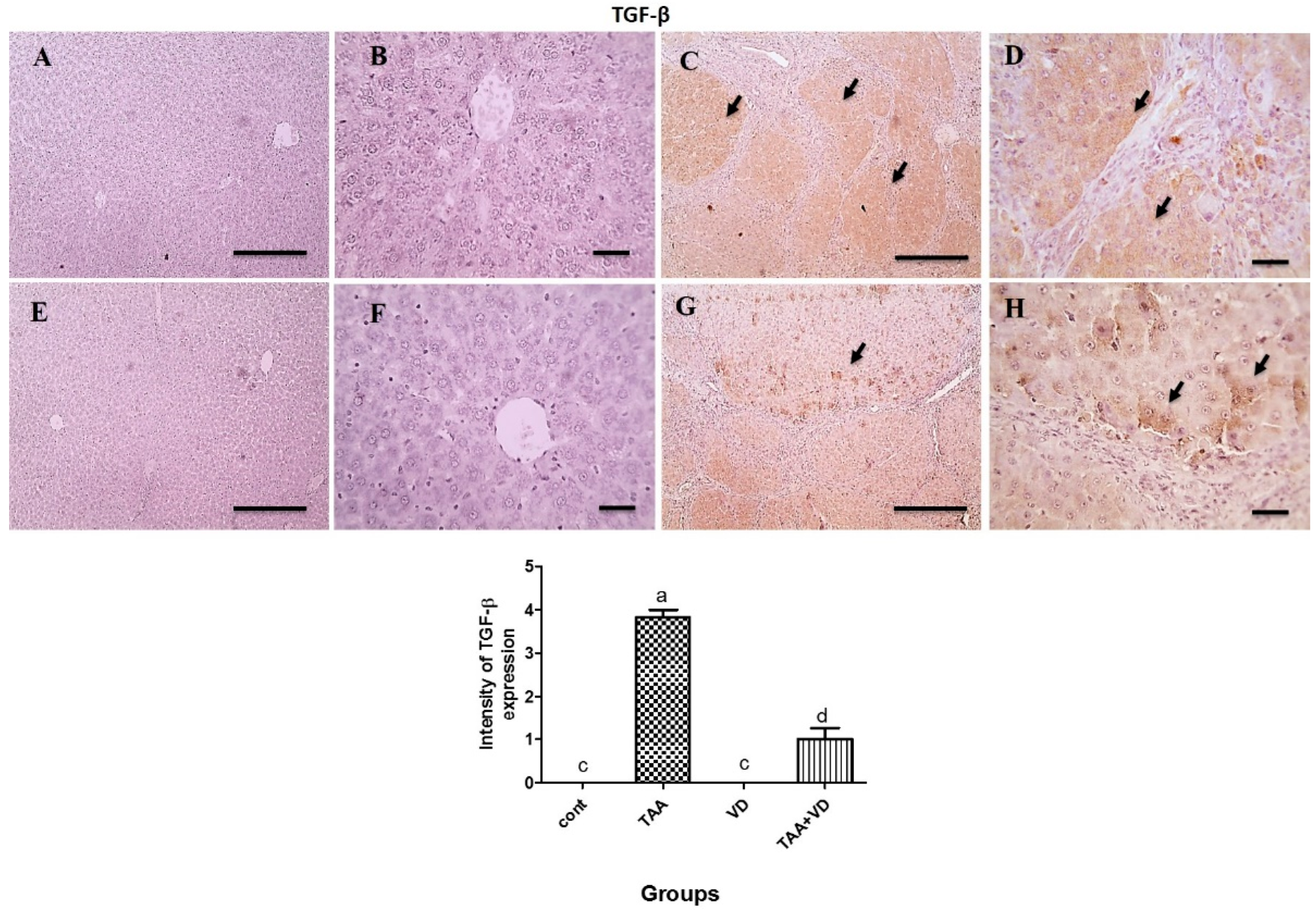
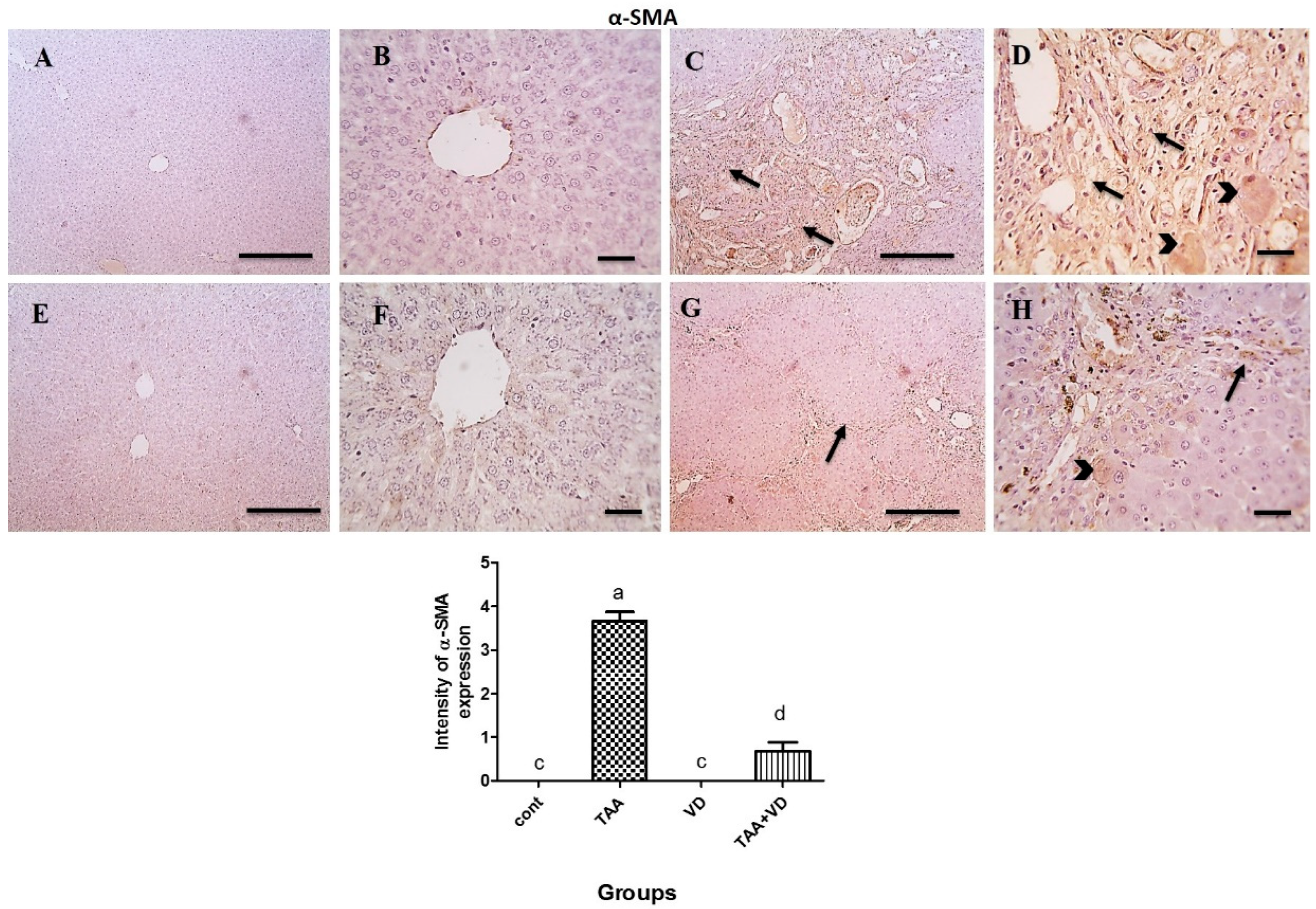
3.6. Immunohistochemical Expression of TGF-β and α-SMA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahsan, R.; Islam, K.M.; Musaddik, A.; Haque, E. Hepatoprotective activity of methanol extract of some medicinal plants against carbon tetrachloride induced hepatotoxicity in albino rats. Glob. J. Pharmacol. 2009, 3, 116–122. [Google Scholar]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Alegre, F.; Pelegrin, P.; Feldstein, A.E. Inflammasomes in liver fibrosis. Semin. Liver Dis. 2017, 37, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Insel, P.A.; Murray, F.; Yokoyama, U.; Romano, S.; Yun, H.; Brown, L.; Snead, A.; Lu, D.; Aroonsakool, N. cAMP and Epac in the regulation of tissue fibrosis. Br. J. Pharmacol. 2012, 166, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Wahlang, B.; McClain, C.; Barve, S.; Gobejishvili, L. Role of cAMP and phosphodiesterase signaling in liver health and disease. Cell. Signal. 2018, 49, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Stumm, C.L.; Wettlaufer, S.H.; Jancar, S. Peters-Golden M. Airway remodeling in murine asthma correlates with a defect in PGE2 synthesis by lung fibroblasts. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2011, 301, L636–L644. [Google Scholar] [CrossRef]
- El-Mihi, K.A.; Kenawy, H.I.; El-Karef, A.; Elsherbiny, N.M.; Eissa, L.A. Naringin attenuates thioacetamide-induced liver fibrosis in rats through modulation of the PI3K/Akt pathway. Life Sci. 2017, 187, 50–57. [Google Scholar] [CrossRef]
- Gad, S.C. Thioacetamide. In Encyclopedia of Toxicology; Elsevier Science & Technology: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Abbasi, M.H.; Akhtar, T.; Malik, I.A.; Fatima, S.; Khawar, B.; Mujeeb, K.A.; Mustafa, G.; Hussain, S.; Iqbal, J.; Sheikh, N. Acute and chronic toxicity of thioacteamide and alterations in blood cell indices in rats. J. Cancer Ther. 2013, 04, 251–259. [Google Scholar] [CrossRef]
- Yogalakshmi, B.; Viswanathan, P.; Anuradha, C.V. Investigation of antioxidant, anti-inflammatory and DNA-protective properties of eugenol in thioacetamide-induced liver injury in rats. Toxicology 2010, 268, 204–212. [Google Scholar] [CrossRef]
- Eissa, L.A.; Kenawy, H.I.; El-Karef, A.; Elsherbiny, N.M.; El-Mihi, K.A. Antioxidant and anti-inflammatory activities of berberine attenuate hepatic fibrosis induced by thioacetamide injection in rats. Chem. Biol. Interact. 2018, 294, 91–100. [Google Scholar] [CrossRef]
- .Holick, M.F. Vitamin D: Evolutionary, physiological and health perspectives. Curr. Drug Targets 2011, 12, 4–18. [Google Scholar] [CrossRef]
- Nagpal, S.; Na, S.; Rathnachalam, R. Noncalcemic actions of vitamin D receptor ligands. Endocr. Rev. 2005, 26, 662–687. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D: Important for prevention of osteoporosis, cardiovascular heart disease, type 1 diabetes, autoimmune diseases, and some cancers. South. Med. J. 2005, 98, 1024–1027. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.; Maseeh, A. Vitamin D: The “sunshine” vitamin. J. Pharmacol. Pharmacother. 2012, 3, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Labudzynskyi, D.O.; Zaitseva, O.V.; Latyshko, N.V.; Gudkova, O.O.; Veliky, M.M. Vitamin d3 contribution to the regulation of oxidative metabolism in the liver of diabetic mice. Ukr. Biochem. J. 2015, 87, 75–90. [Google Scholar] [CrossRef]
- Wacker, M.; Holick, M.F. Sunlight and vitamin D: A global perspective for health. Dermatoendocrinol 2013, 5, 51–108. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.M.Y.; Chen, K.B.; Chao, P.L. Antioxidative effect of vitamin D3 on zinc-induced oxidative stress in CNS. Ann. New York Acad. Sci. 2005, 1053, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.; Ismail, A. Vitamin D treatment protects against and reverses oxidative stress induced muscle proteolysis. J. Steroid Biochem. Mol. Biol. 2015, 152, 171–179. [Google Scholar] [CrossRef]
- Alatawi, F.S.; Faridi, U.A.; Alatawi, M.S. Effect of treatment with vitamin D plus calcium on oxidative stress in streptozotocin-induced diabetic rats. Saudi Pharm. J. 2018, 26, 1208–1213. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Salama, A.; Salama, R.A.A. Therapeutic Effect of Dunaliella salina Microalgae on Thioacetamide- (TAA-) Induced Hepatic Liver Fibrosis in Rats: Role of TGF- β and MMP9. BioMed Res. Int. 2019, 2019, 7028314. [Google Scholar] [CrossRef]
- Abo El-Magd, N.F.; El-Karef, A.; El-Shishtawy, M.M.; Eissa, L.A. Hepatoprotective effects of glycyrrhizin and omega-3 fatty acids on Nuclear Factor-kappa B pathway in thioacetamide-induced fibrosis in rats. Egypt. J. Basic Appl. Sci. 2015, 2, 65–74. [Google Scholar] [CrossRef]
- Wahsh, E.; Abu-Elsaad, N.; El-Karef, A.; Ibrahim, T. The vitamin D receptor agonist, calcipotriol, modulates fibrogenic pathways mitigating liver fibrosis in-vivo: An experimental study. Eur. J. Pharmacol. 2016, 789, 362–369. [Google Scholar] [CrossRef]
- BaSalamah, M.A.; Abdelghany, A.H.; El-Boshy, M.; Ahmad, J.; Idris, S.; Refaat, B. Vitamin D alleviates lead induced renal and testicular injuries by immunomodulatory and antioxidant mechanisms in rats. Sci. Rep. 2018, 8, 4853. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D status: Measurement, interpretation, and clinical application. Ann. Epidemiol. 2009, 19, 73–78. [Google Scholar] [CrossRef]
- Banni, M.; Messaoudi, I.; Said, L.; El Heni, J.; Kerkeni, A.; Said, K. Metallothionein gene expression in liver of rats exposed to cadmium and supplemented with zinc and selenium. Arch. Environ. Contam. Toxicol. 2010, 59, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Matsuda, A.; An, J.; Koshiba, R.; Nishio, J.; Negishi, H.; Ikushima, H.; Onoe, T.; Ohdan, H.; Yoshida, N.; et al. Conditional ablation of HMGB1 in mice reveals its protective function against endotoxemia and bacterial infection. Proc. Natl. Acad. Sci. USA 2013, 110, 20699–20704. [Google Scholar] [CrossRef]
- Katsuno, K.; Burrows, J.N.; Duncan, K.; Katsuno, K.; Burrows, J.N.; Duncan, K.; Van Huijsduijnen, R.H.; Kaneko, T.; Kita, K.; Mowbray, C.E.; et al. Hit and lead criteria in drug discovery for infectious diseases of the developing world. Nat. Rev. Drug Discov. 2015, 14, 751–758. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Atorrasagasti, C.; Piccioni, F.; Borowski, S.; Tirado-González, I.; Freitag, N.; Cantero, M.J.; Bayo, J.; Mazzolini, G.; Alaniz, L.D.; Blois, S.M.; et al. Acceleration of TAA-Induced Liver Fibrosis by Stress Exposure Is Associated with Upregulation of Nerve Growth Factor and Glycopattern Deviations. Int. J. Mol. Sci. 2021, 22, 5055. [Google Scholar] [CrossRef]
- Alkreathy, H.M.; Esmat, A. Lycorine Ameliorates Thioacetamide-Induced Hepatic Fibrosis in Rats: Emphasis on Antioxidant, Anti-Inflammatory, and STAT3 Inhibition Effects. Pharmaceuticals 2022, 15, 369. [Google Scholar] [CrossRef]
- Norušis, M.J. SPSS 14.0 Guide to Data Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 2006. [Google Scholar]
- Schuppan, D.; Kim, Y.O. Evolving therapies for liver fibrosis. J. Clin. Investig. 2013, 123, 1887–1901. [Google Scholar] [CrossRef] [PubMed]
- Reiter, F.P.; Hohenester, S.; Nagel, J.M.; Wimmer, R.; Artmann, R.; Wottke, L.; Makeschin, M.C.; Mayr, D.; Rust, C.; Trauner, M.; et al. 1,25-(OH)₂-vitamin D₃ prevents activation of hepatic stellate cells in vitro and ameliorates inflammatory liver damage but not fibrosis in the Abcb4(-/-) model. Biochem. Biophys. Res. Commun. 2015, 459, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Czechowska, G.; Celinski, K.; Korolczuk, A.; Czechowska, G.; Celinski, K.; Korolczuk, A.; Wojcicka, G.; Dudka, J.; Bojarska, A.; Reiter, R.J. Protective effects of melatonin against thioacetamide-induced liver fibrosis in rats. J. Physiol. Pharmacol. 2015, 66, 567–579. [Google Scholar] [PubMed]
- Abd-Elrahman, N.M.; Farid, A.S.; Fararh, K.M. Effect of gentamicin and thioacetamide toxicity on serum proteins. Benha Vet. Med. J. 2014, 27, 456–465. [Google Scholar]
- El-Boshy, M.; BaSalamah, M.A.; Ahmad, J.; Idris, S.; Mahbub, A.; Abdelghany, A.H.; Almaimani, R.A.; Almasmoum, H.; Ghaith, M.M.; Elzubier, M.; et al. Vitamin D protects against oxidative stress, inflammation and hepatorenal damage induced by acute paracetamol toxicity in rat. Free. Radic. Biol. Med. 2019, 141, 310–321. [Google Scholar] [CrossRef]
- Özdemir-Kumral, Z.N.; Erkek, B.E.; Karakuş, B.; Almacı, M.; Fathi, R.; Yüksel, M.; Cumbul, A.; Alican, I. Potential Effect of 1,25 Dihydroxyvitamin D3 on Thioacetamide-Induced Hepatotoxicity in Rats. J. Surg. Res. 2019, 243, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rehim, W.M.; El-Tahan, R.A.; El-Tarawy, M.A.; Shehata, R.R.; Kamel, M.A. The possible antidiabetic effects of vitamin D receptors agonist in rat model of type 2 diabetes. Mol. Cell. Biochem. 2019, 450, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.R.; Islam, M.K.; Haque, Z. Effects of Vitamin-D and sunlight on the hemato-biochemical parameters in mice. Int. J. Curr. Res. Rev. 2016, 8, 23. [Google Scholar]
- Ebrahim, A.T.; El-Gendy, A.M.; El-Zawahry, B. Antihepatotoxic potential of ginseng (Panax ginseng) in thioacetamide-induced acute hepatocellular injury in rats. EJHM 2004, 16, 55–64. [Google Scholar] [CrossRef]
- Abdalla, O.; Elboshy, M.; Risha, E.; Mohammed, F.H.; Abu-Ghanema, W. Hepatoprotective and antioxidant effects of pomegranate against thioacetamide toxicity in rats. Ann Vet. Anim. Sci. 2016, 3, 28–40. [Google Scholar]
- Calle, C.; Maestro, B.; García-Arencibia, M. 1,25-Dihydroxyvitamin D3 effects on the regulation of the insulin receptor gene in the hind limb muscle and heart of streptozotocin-induced diabetic rats. AJMB 2013, 3, 87–97. [Google Scholar] [CrossRef]
- Chagas, C.E.A.; Borges, M.C.; Martini, L.A.; Rogero, M.M. Focus on vitamin D, inflammation and type 2 diabetes. Nutrients 2012, 4, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Shaker, M.E.; Salem, H.A.; Shiha, G.E.; Ibrahim, T.M. Nilotinib counteracts thioacetamide-induced hepatic oxidative stress and attenuates liver fibrosis progression. Fundam. Clin. Pharmacol. 2011, 25, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Abdel Salam, O.M.E.; Mohammed, N.A.; Sleem, A.A.; Farrag, A.R. The effect of antidepressant drugs on thioacetamide-induced oxidative stress. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 735–744. [Google Scholar]
- Almasmoum, H.; Refaat, B.; Ghaith, M.M.; Almaimani, R.A.; Idris, S.; Ahmad, J.; Abdelghany, A.H.; BaSalamah, M.A.; El-Boshy, M. Protective effect of Vitamin D3 against lead induced hepatotoxicity, oxidative stress, immunosuppressive and calcium homeostasis disorders in rat. Environ. Toxicol. Pharmacol. 2019, 72, 103246. [Google Scholar] [CrossRef]
- Wiseman, H. Vitamin D is a membrane antioxidant. Ability to inhibit iron-dependent lipid peroxidation in liposomes compared to cholesterol, ergosterol and tamoxifen and relevance to anticancer action. FEBS Lett. 1993, 326, 285–288. [Google Scholar] [CrossRef]
- Crosas-Molist, E.; Fabregat, I. Role of NADPH oxidases in the redox biology of liver fibrosis. Redox Biol. 2015, 6, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Bucala, R.; Spiegel, L.A.; Chesney, J.; Hogan, M.; Cerami, A. Circulating fibrocytes define a new leukocyte subpopulation that mediates tissue repair. Mol. Med. 1994, 1, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Kisseleva, T.; Uchinami, H.; Feirt, N.; Quintana-Bustamante, O.; Segovia, J.C.; Schwabe, R.F.; Brenner, D.A. Bone marrow-derived fibrocytes participate in pathogenesis of liver fibrosis. J. Hepatol. 2006, 45, 429–438. [Google Scholar] [CrossRef]
- Russo, F.P.; Alison, M.R.; Bigger, B.W.; Amofah, E.; Florou, A.; Amin, F.; Bou–Gharios, G.; Jeffery, R.; Iredale, J.P.; Forbes, S.J. The bone marrow functionally contributes to liver fibrosis. Gastroenterology 2006, 130, 1807–1821. [Google Scholar] [CrossRef]
- Roderfeld, M.; Rath, T.; Voswinckel, R.; Dierkes, C.; Dietrich, H.; Zahner, D.; Graf, J.; Roeb, E. Bone marrow transplantation demonstrates medullar origin of CD34+ fibrocytes and ameliorates hepatic fibrosis in Abcb4-/- mice. Hepatology 2010, 51, 267–276. [Google Scholar] [CrossRef]
- Hao, H.; Li, X.; Li, Q.; Lin, H.; Chen, Z.; Xie, J.; Xuan, W.; Liao, W.; Bin, J.; Huang, X.; et al. FGF23 promotes myocardial fibrosis in mice through activation of β-catenin. Oncotarget 2016, 7, 64649–64664. [Google Scholar] [CrossRef]
- Kuga, K.; Kusakari, Y.; Uesugi, K.; Semba, K.; Urashima, T.; Akaike, T.; Minamisawa, S. Fibrosis growth factor 23 is a promoting factor for cardiac fibrosis in the presence of transforming growth factor-β1. PLoS ONE 2020, 15, e0231905. [Google Scholar] [CrossRef] [PubMed]
- Prié, D.; Forand, A.; Francoz, C.; Elie, C.; Cohen, I.; Courbebaisse, M.; Eladari, D.; Lebrec, D.; Durand, F.; Friedlander, G. Plasma fibroblast growth factor 23 concentration is increased and predicts mortality in patients on the liver-transplant waiting list. PLoS ONE 2013, 8, e66182. [Google Scholar] [CrossRef]
- Chen, I.-S.; Chen, Y.-C.; Chou, C.-H.; Chuang, R.-F.; Sheen, L.-Y.; Chiu, C.-H. Hepatoprotection of silymarin against thioacetamide-induced chronic liver fibrosis. J. Sci. Food Agric. 2012, 92, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Eliades, M.; Spyrou, E.; Agrawal, N.; Lazo, M.; Brancati, F.L.; Potter, J.J.; Koteish, A.A.; Clark, J.M.; Guallar, E.; Hernaez, R. Meta-analysis: Vitamin D and non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2013, 38, 246–254. [Google Scholar] [CrossRef]
- Seki, E.; Schwabe, R.F. Hepatic inflammation and fibrosis: Functional links and key pathways. Hepatology 2015, 61, 1066–1079. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kum, Y.S.; Lee, T.I.; Kim, S.J.; Lee, W.R.; Kim, B.I.; Kim, H.S.; Kim, K.H.; Park, K.K. Melittin attenuates liver injury in thioacetamide-treated mice through modulating inflammation and fibrogenesis. Exp. Biol. Med. 2011, 236, 1306–1313. [Google Scholar] [CrossRef] [PubMed]
- Abramovitch, S.; Dahan-Bachar, L.; Sharvit, E.; Weisman, Y.; Tov, A.B.; Brazowski, E.; Reif, S. Vitamin D inhibits proliferation and profibrotic marker expression in hepatic stellate cells and decreases thioacetamide-induced liver fibrosis in rats. Gut 2011, 60, 1728–1737. [Google Scholar] [CrossRef]
- Chen, T.; Zuo, X.; Wang, S.; Yu, P.; Yuan, J.; Wei, S.; Chen, J.; Sun, Y.; Gao, Y.; Li, X. The effect of vitamin D supplementation on the progression of fibrosis in patients with chronic liver disease: A protocol for a systematic review and meta-analysis. Medicine 2020, 99, e20296. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Reference |
|---|---|---|
| Rat β-actin | F:3′TCCTCCTGAGCGCAAGTACTCT5′ R:5′GCTCAGTAACAGTCCGCCTAGAA3′ | [26] |
| CD34 | F:3′AGCCATGTGCTCACACATCA5′ R:5′CAAACACTCGGGCCTAACCT3′ | [27] |
| FGF23 | F:3′ACGGAACACCCCATCAGACTATC5′ R: 5′TATCACTACGGAGCCAGCATCCTC3′ | [28] |
| Parameter | Cont. | TAA | VD | TAA+VD |
|---|---|---|---|---|
| ALT (U/L) | 43.50 ± 1.17 b | 362.38 ± 47.56 a | 42.94 ± 0.75 b | 103.56 ± 2.78 b |
| AST (U/L) | 136.80 ± 10.32 c | 1178.20 ± 24.58 a | 130.8 ± 12.06 c | 600 ± 154.36 b |
| ALP (U/L) | 148.20 ±8.95 c | 424.80 ± 49.72 ab | 127.70 ± 7.27 c | 330.20 ± 11.52 b |
| Total bilirubin (mg/dL) | 0.31 ± 0.05 c | 1.28 ± 0.18 a | 0.41 ± 0.07 c | 0.89 ± 0.06 b |
| Direct bilirubin (mg/dL) | 0.12 ± 0.03 c | 0.95 ± 0.18 a | 0.19 ± 0.04 c | 0.62 ± 0.08 b |
| Indirect bilirubin (mg/dL) | 0.18 ± 0.04 a | 0.33 ± 0.00 a | 0.21 ± 0.05 a | 0.27 ± 0.06 a |
| Total protein (g/dL) | 8.42 ± 0.20 a | 6.82 ± 0.24 b | 8.13 ± 0.24 a | 6.99 ± 0.25 b |
| Albumin (g/dL) | 4.13 ± 0.14 a | 3.09 ± 0.11 b | 4.06 ± 0.13 a | 3.38 ± 0.15 b |
| Globulin (g/dL) | 4.29 ± 0.21 a | 3.73 ± 0.24 a | 4.07 ± 0.18 a | 3.61 ± 0.19 a |
| A/G ratio | 0.97 ± 0.06 a | 0.84 ± 0.06 a | 1.00 ± 0.05 a | 0.94 ± 0.05 a |
| Parameter | Cont. | TAA | VD | TAA+VD |
|---|---|---|---|---|
| TC (mg/dL) | 50.49 ± 2.29 b | 67.64 ± 1.44 a | 53.57 ± 0.71 b | 58.55 ± 1.19 ab |
| TG (mg/dL) | 100.26 ± 4.06 c | 165.80 ± 10.96 a | 99.36 ± 6.48 c | 133.40 ± 11.66 b |
| Glucose (mg/dL) | 96.26 ± 2.88 a | 68.13 ± 4.60 b | 94.60 ± 3.56 a | 89.25 ± 8.6 ab |
| VD (25[OH] D3) (ng/mL) | 34.35 ± 2.05 c | 2.76 ± 0.14 d | 148.33±4.40 a | 122.33±1.45 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Megahed, A.; Gadalla, H.; Abdelhamid, F.M.; Almehmadi, S.J.; Khan, A.A.; Albukhari, T.A.; Risha, E.F. Vitamin D Ameliorates the Hepatic Oxidative Damage and Fibrotic Effect Caused by Thioacetamide in Rats. Biomedicines 2023, 11, 424. https://doi.org/10.3390/biomedicines11020424
Megahed A, Gadalla H, Abdelhamid FM, Almehmadi SJ, Khan AA, Albukhari TA, Risha EF. Vitamin D Ameliorates the Hepatic Oxidative Damage and Fibrotic Effect Caused by Thioacetamide in Rats. Biomedicines. 2023; 11(2):424. https://doi.org/10.3390/biomedicines11020424
Chicago/Turabian StyleMegahed, Aya, Hossam Gadalla, Fatma M. Abdelhamid, Samah J. Almehmadi, Anmar A. Khan, Talat A. Albukhari, and Engy F. Risha. 2023. "Vitamin D Ameliorates the Hepatic Oxidative Damage and Fibrotic Effect Caused by Thioacetamide in Rats" Biomedicines 11, no. 2: 424. https://doi.org/10.3390/biomedicines11020424
APA StyleMegahed, A., Gadalla, H., Abdelhamid, F. M., Almehmadi, S. J., Khan, A. A., Albukhari, T. A., & Risha, E. F. (2023). Vitamin D Ameliorates the Hepatic Oxidative Damage and Fibrotic Effect Caused by Thioacetamide in Rats. Biomedicines, 11(2), 424. https://doi.org/10.3390/biomedicines11020424