

IZUMO1 Receptor Localization during Hyaluronic Acid Selection in Human Spermatozoa
 , , , and
, , , and 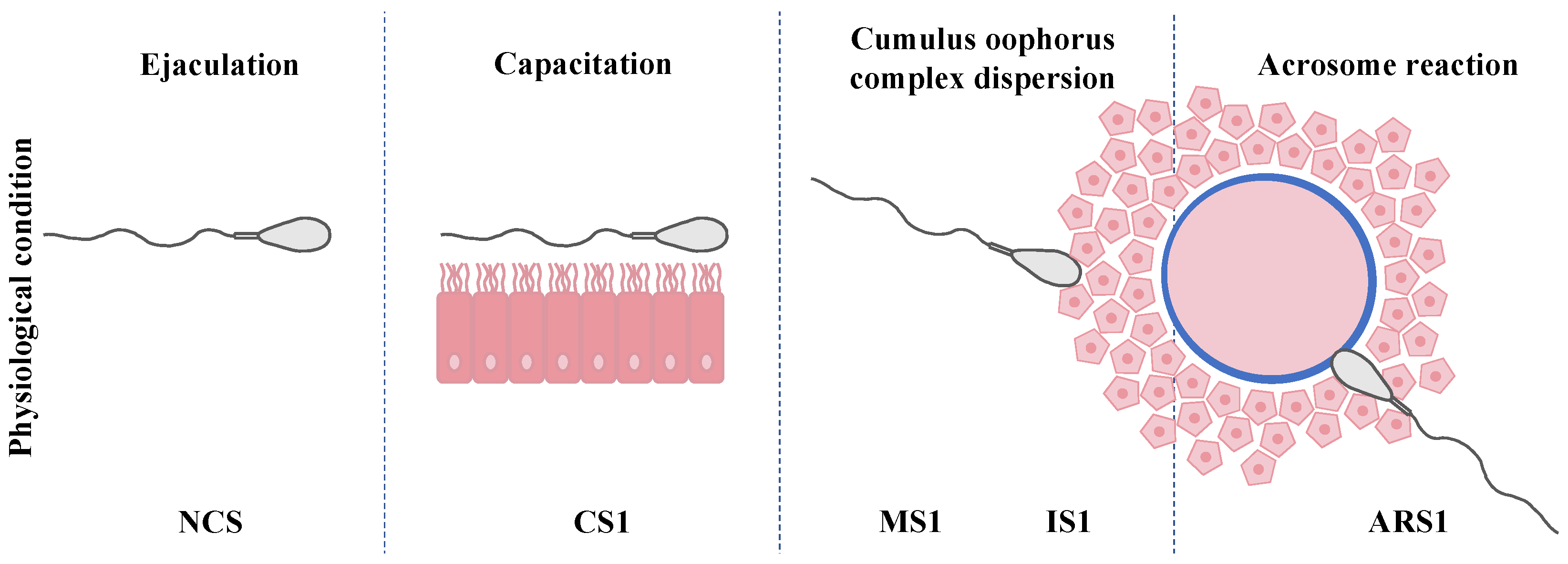{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Semen Sample Analysis
2.3. Sperm Capacitation by Swim-Up
2.4. Hyaluronic Acid Sperm Selection
2.5. Induction and Evaluation of Acrosomal Reaction
2.6. Fixation
2.7. Immunolocation of IZUMO1
2.8. Immunofluorescent Costaining of IZUMO1 and Acrosome Status
2.9. Statistical Analysis
3. Results
3.1. Seminal Sample Analysis
3.2. Acrosome Reaction Assesment
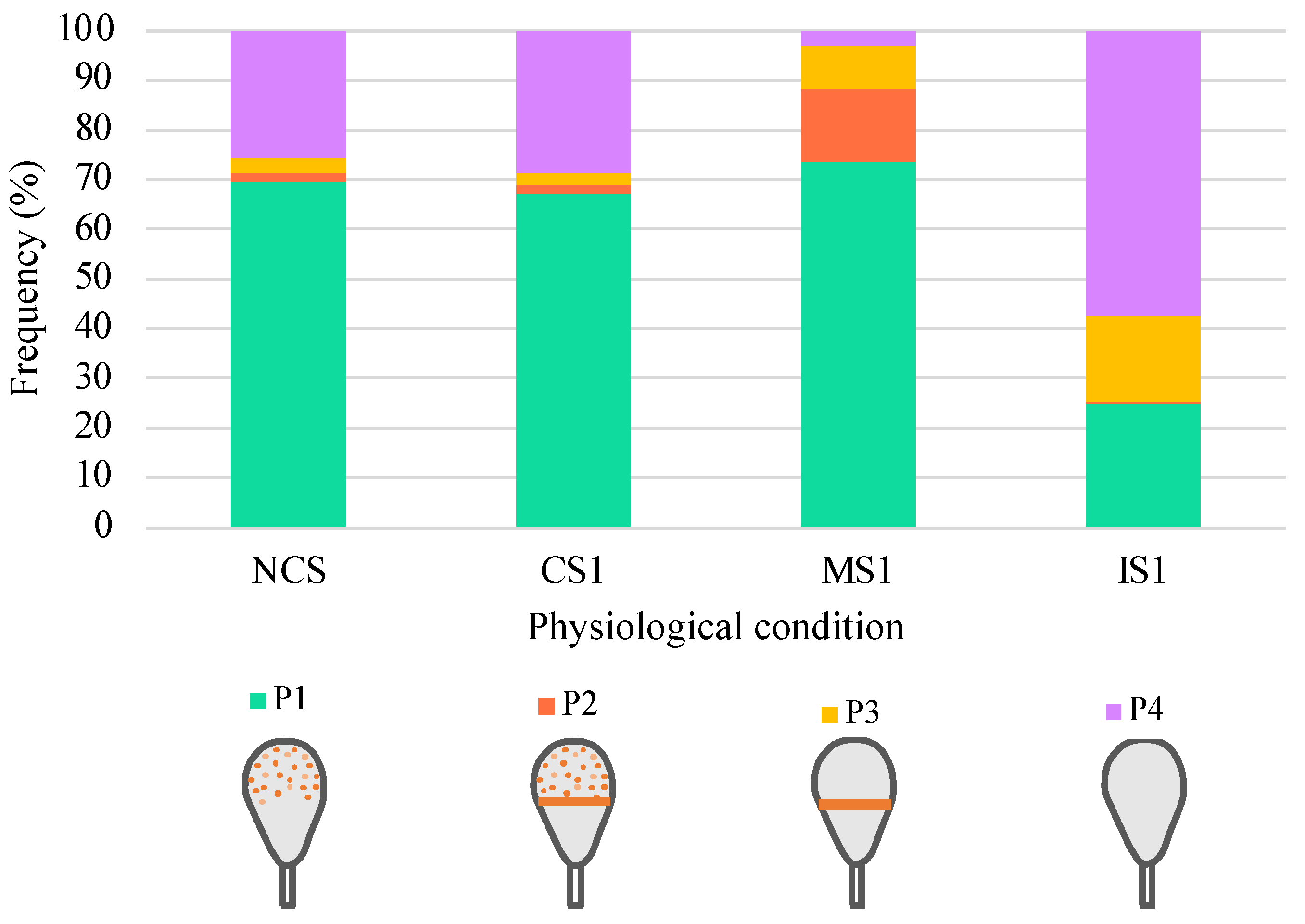
3.3. Distribution of IZUMO1 in Noncapacitated Sperm, One-Hour-Capacitated Sperm, and Selected Human Sperm by Hyaluronic Acid Test
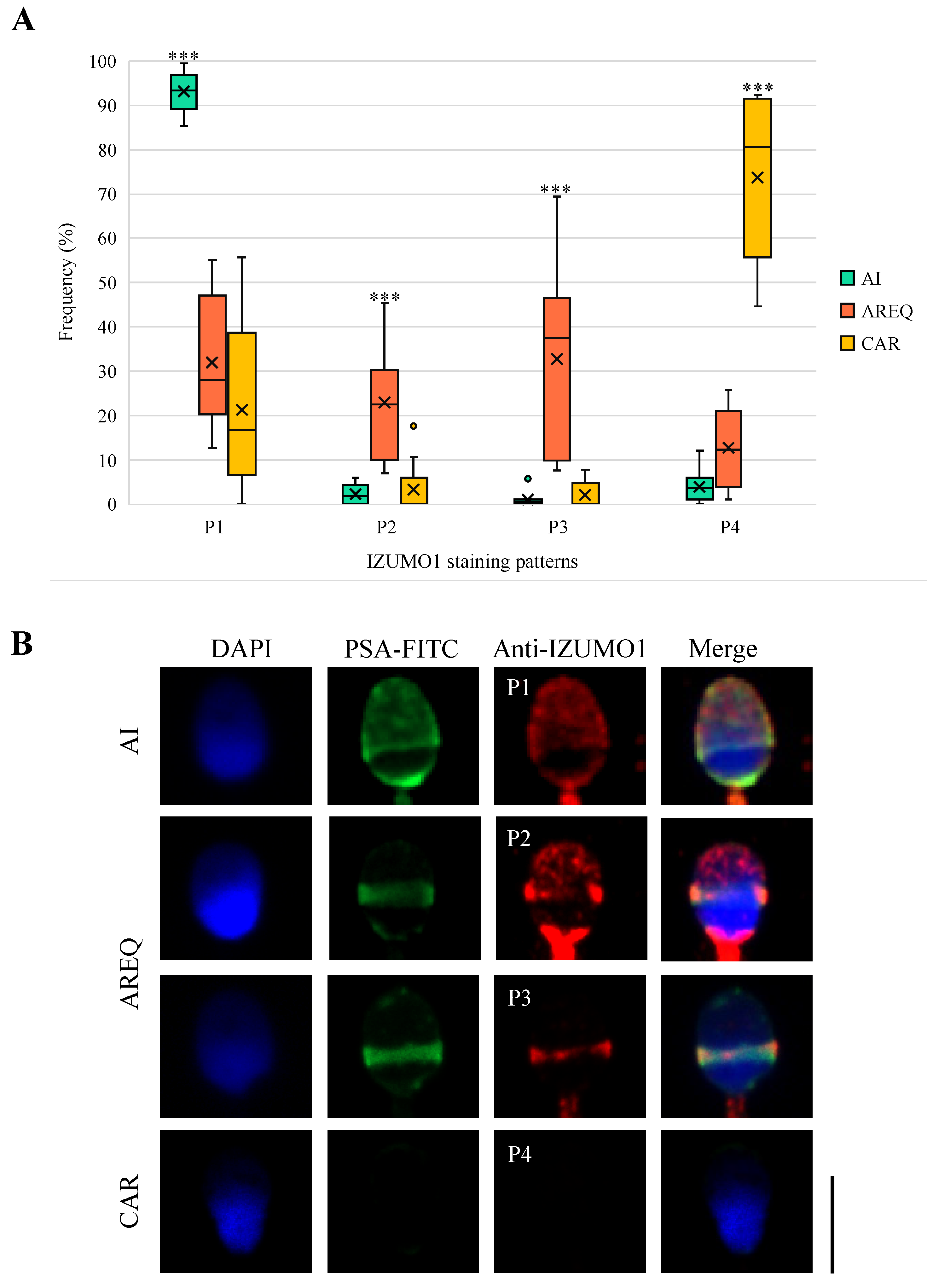
3.4. Distribution of IZUMO1 in Induced Acrosome Reaction from One-Hour-Capacitated Sperm According to Acrosomal Status
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Jonge, C. Biological basis for human capacitation-revisited. Hum. Reprod. Update 2017, 23, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Espinosa, P.; Torrijo-Boix, S.; Huerta-Retamal, N.; Avilés, M.; Aizpurua, J.; Romero, A.; Gómez-Torres, M.J. Capacitation and acrosome reaction are associated with changes in sialic acid location and head morphometry in human sperm. Rev. Int. Androl. 2018, 16, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Matamoros-Volante, A.; Moreno-Irusta, A.; Torres-Rodriguez, P.; Giojalas, L.; Gervasi, M.G.; Visconti, P.E.; Treviño, C.L. Semi-automatized segmentation method using image-based flow cytometry to study sperm physiology: The case of capacitation-induced tyrosine phosphorylation. Mol. Hum. Reprod. 2018, 24, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Redgrove, K.A.; Nixon, B.; Baker, M.A.; Hetherington, L.; Baker, G.; Liu, D.Y.; Aitken, R.J. The molecular chaperone HSPA2 plays a key role in regulating the expression of sperm surface receptors that mediate sperm-egg recognition. PLoS ONE 2012, 7, e50851. [Google Scholar] [CrossRef] [PubMed]
- Cafe, S.L.; Anderson, A.L.; Nixon, B. In vitro Induction and Detection of Acrosomal Exocytosis in Human Spermatozoa. Bio-protocol 2020, 10, e3689. [Google Scholar] [CrossRef] [PubMed]
- Ito, C.; Toshimori, K. Acrosome markers of human sperm. Anat. Sci. Int. 2016, 91, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, L.; Kimata, K. Cumulus oophorus extracellular matrix: Its construction and regulation. Cell Struct. Funct. 2001, 26, 189–196. [Google Scholar] [CrossRef]
- Prinosilova, P.; Kruger, T.; Sati, L.; Ozkavukcu, S.; Vigue, L.; Kovanci, E.; Huszar, G. Selectivity of hyaluronic acid binding for spermatozoa with normal Tygerberg strict morphology. Reprod. Biomed. Online 2009, 18, 177–183. [Google Scholar] [CrossRef]
- Myles, D.G.; Primakoff, P. Why did the sperm cross the cumulus? To get to the oocyte. Functions of the sperm surface proteins PH-20 and fertilin in arriving at, and fusing with, the egg. Biol. Reprod. 1997, 56, 320–327. [Google Scholar] [CrossRef][Green Version]
- Rashki Ghaleno, L.; Rezazadeh Valojerdi, M.; Chehrazi, M.; Sahraneshin Samani, F.; Salman Yazdi, R. Hyaluronic Acid Binding Assay Is Highly Sensitive to Select Human Spermatozoa with Good Progressive Motility, Morphology, and Nuclear Maturity. Gynecol. Obstet. Investig. 2016, 81, 244–250. [Google Scholar] [CrossRef]
- Huszar, G.; Ozenci, C.C.; Cayli, S.; Zavaczki, Z.; Hansch, E.; Vigue, L. Hyaluronic acid binding by human sperm indicates cellular maturity, viability, and unreacted acrosomal status. Fertil. Steril. 2003, 79, 1616–1624. [Google Scholar] [CrossRef] [PubMed]
- Huszar, G.; Jakab, A.; Sakkas, D.; Ozenci, C.C.; Cayli, S.; Delpiano, E.; Ozkavukcu, S. Fertility testing and ICSI sperm selection by hyaluronic acid binding: Clinical and genetic aspects. Reprod. Biomed. Online 2007, 14, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Cayli, S.; Jakab, A.; Ovari, L.; Delpiano, E.; Celik-Ozenci, C.; Sakkas, D.; Ward, D.; Huszar, G. Biochemical markers of sperm function: Male fertility and sperm selection for ICSI. Reprod. Biomed. Online 2003, 7, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Scaruffi, P.; Bovis, F.; Casciano, I.; Maccarini, E.; De Leo, C.; Gazzo, I.; Massarotti, C.; Sozzi, F.; Stigliani, S.; Anserini, P. Hyaluronic acid-sperm selection significantly improves the clinical outcome of couples with previous ICSI cycles failure. Andrology 2022, 10, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Pavitt, S.; Sharma, V.; Forbes, G.; Hooper, R.; Bhattacharya, S.; Kirkman-Brown, J.; Coomarasamy, A.; Lewis, S.; Cutting, R.; et al. Physiological, hyaluronan-selected intracytoplasmic sperm injection for infertility treatment (HABSelect): A parallel, two-group, randomised trial. Lancet 2019, 393, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Novoselsky Persky, M.; Hershko-Klement, A.; Solnica, A.; Bdolah, Y.; Hurwitz, A.; El Gilad, M.K.; Nefesh, I.; Esh-Broder, E. Conventional ICSI vs. physiological selection of spermatozoa for ICSI (picsi) in sibling oocytes. Andrology 2021, 9, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Mokánszki, A.; Tóthné, E.V.; Bodnár, B.; Tándor, Z.; Molnár, Z.; Jakab, A.; Ujfalusi, A.; Oláh, É. Is sperm hyaluronic acid binding ability predictive for clinical success of intracytoplasmic sperm injection: PICSI vs. ICSI? Syst. Biol. Reprod. Med. 2014, 60, 348–354. [Google Scholar] [CrossRef]
- Parmegiani, L.; Cognigni, G.E.; Bernardi, S.; Troilo, E.; Ciampaglia, W.; Filicori, M. “Physiologic ICSI”: Hyaluronic acid (HA) favors selection of spermatozoa without DNA fragmentation and with normal nucleus, resulting in improvement of embryo quality. Fertil. Steril. 2010, 93, 598–604. [Google Scholar] [CrossRef]
- Brukman, N.G.; Nakajima, K.P.; Valansi, C.; Flyak, K.; Li, X.; Higashiyama, T.; Podbilewicz, B. A novel function for the sperm adhesion protein IZUMO1 in cell-cell fusion. J. Cell Biol. 2023, 222, e202207147. [Google Scholar] [CrossRef]
- Yanagimachi, R. Mysteries and unsolved problems of mammalian fertilization and related topics. Biol. Reprod. 2022, 106, 644–675. [Google Scholar] [CrossRef]
- Sebkova, N.; Ded, L.; Vesela, K.; Dvorakova-Hortova, K. Progress of sperm IZUMO1 relocation during spontaneous acrosome reaction. Reproduction 2014, 147, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Sosnik, J.; Miranda, P.V.; Spiridonov, N.A.; Yoon, S.Y.; Fissore, R.A.; Johnson, G.R.; Visconti, P.E. Tssk6 is required for Izumo relocalization and gamete fusion in the mouse. J. Cell Sci. 2009, 122, 2741–2749. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, E.; Doe, B.; Goulding, D.; Wright, G.J. Juno is the egg Izumo receptor and is essential for mammalian fertilization. Nature 2014, 508, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Aydin, H.; Sultana, A.; Li, S.; Thavalingam, A.; Lee, J.E. Molecular architecture of the human sperm IZUMO1 and egg JUNO fertilization complex. Nature 2016, 534, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Ohto, U.; Ishida, H.; Krayukhina, E.; Uchiyama, S.; Inoue, N.; Shimizu, T. Structure of IZUMO1–JUNO reveals sperm–oocyte recognition during mammalian fertilization. Nature 2016, 534, 566–569. [Google Scholar] [CrossRef]
- Martinez, G.; Cappetta, D.; Telesca, M.; Urbanek, K.; Castaldo, G.; Dhellemmes, M.; Mele, V.G.; Chioccarelli, T.; Porreca, V.; Barbotin, A.L.; et al. Cytochalasin D restores nuclear size acting on F-actin and IZUMO1 localization in low-quality spermatozoa. Int. J. Biol. Sci. 2023, 19, 2234. [Google Scholar] [CrossRef]
- Inoue, N.; Wada, I. Monitoring dimeric status of IZUMO1 during the acrosome reaction in living spermatozoon. Cell Cycle 2018, 17, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Ikawa, M.; Isotani, A.; Okabe, M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature 2005, 434, 234–238. [Google Scholar] [CrossRef]
- Gómez-Torres, M.J.; Huerta-Retamal, N.; Sáez-Espinosa, P.; Robles-Gómez, L.; Avilés, M.; Aizpurua, J. Molecular Chaperone HSPA2 Distribution During Hyaluronic Acid Selection in Human Sperm. Reprod. Sci. 2022, 30, 1176–1185. [Google Scholar] [CrossRef]
- Gómez-Torres, M.J.; Sáez-Espinosa, P.; Manzano-Santiago, P.; Robles-Gómez, L.; Huerta-Retamal, N.; Aizpurua, J. Sperm Adhesion Molecule 1 (SPAM1) Distribution in Selected Human Sperm by Hyaluronic Acid Test. Biomedicines 2022, 10, 2553. [Google Scholar] [CrossRef]
- Cross, N.L.; Morales, P.; Overstreet, J.W.; Hanson, F.W. Two simple methods for detecting acrosome-reacted human sperm. Gamete Res. 1986, 15, 213–226. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2021; ISBN 978 92 4 003078 7. [Google Scholar]
- Calhaz-Jorge, C.; De Geyter, C.; Kupka, M.S.; De Mouzon, J.; Erb, K.; Mocanu, E.; Motrenko, T.; Scaravelli, G.; Wyns, C.; Goossens, V.; et al. Assisted reproductive technology in Europe, 2012: Results generated from European registers by ESHRE. Hum. Reprod. 2016, 31, 1638–1652. [Google Scholar] [CrossRef] [PubMed]
- Belva, F.; Bonduelle, M.; Roelants, M.; Michielsen, D.; Van Steirteghem, A.; Verheyen, G.; Tournaye, H. Semen quality of young adult ICSI offspring: The first results. Hum. Reprod. 2016, 31, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- Kongmanas, K.; Kruevaisayawan, H.; Saewu, A.; Sugeng, C.; Fernandes, J.; Souda, P.; Angel, J.B.; Faull, K.F.; Aitken, R.J.; Whitelegge, J.; et al. Proteomic Characterization of Pig Sperm Anterior Head Plasma Membrane Reveals Roles of Acrosomal Proteins in ZP3 Binding. J. Cell. Physiol. 2015, 230, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Foster, J.A.; Kvasnicka, K.W.; Gerton, G.L. Transitional states of acrosomal exocytosis and proteolytic processing of the acrosomal matrix in guinea pig sperm. Mol. Reprod. Dev. 2011, 78, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Foster, J.A.; Gerton, G.L. Differential Release of Guinea Pig Sperm Acrosomal Components During Exocytosis. Biol. Reprod. 2001, 64, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Kigman, I.; Glassner, M.; Storey, B.T.; Kopf, G.S. Zona pellucida-mediated acrosomal exocytosis in mouse spermatozoa: Characterization of an intermediate stage prior to the completion of the acrosome reaction. Dev. Biol. 1991, 145, 344–355. [Google Scholar] [CrossRef]
- Yanagimachi, R. Mammalian sperm acrosome reaction: Where does it begin before fertilization? Biol. Reprod. 2011, 85, 4–5. [Google Scholar] [CrossRef]
- Torabi, F.; Binduraihem, A.; Miller, D. Sedimentation properties in density gradients correspond with levels of sperm DNA fragmentation, chromatin compaction and binding affinity to hyaluronic acid. Reprod. Biomed. Online 2017, 34, 298–311. [Google Scholar] [CrossRef]
- Yagci, A.; Murk, W.; Stronk, J.; Huszar, G. Spermatozoa bound to solid state hyaluronic acid show chromatin structure with high DNA chain integrity: An acridine orange fluorescence study. J. Androl. 2010, 31, 566–572. [Google Scholar] [CrossRef]
- Fukuda, M.; Sakase, M.; Fukushima, M.; Harayama, H. Changes of IZUMO1 in bull spermatozoa during the maturation, acrosome reaction, and cryopreservation. Theriogenology 2016, 86, 2179–2188.e3. [Google Scholar] [CrossRef]
- Santolaria, P.; Vicente-Fiel, S.; Palacín, I.; Fantova, E.; Blasco, M.E.; Silvestre, M.A.; Yániz, J.L. Predictive capacity of sperm quality parameters and sperm subpopulations on field fertility after artificial insemination in sheep. Anim. Reprod. Sci. 2015, 163, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Satouh, Y.; Inoue, N.; Ikawa, M.; Okabe, M. Visualization of the moment of mouse sperm-egg fusion and dynamic localization of IZUMO1. J. Cell Sci. 2012, 125, 4985–4990. [Google Scholar] [CrossRef] [PubMed]
- Llavanera, M.; Delgado-Bermúdez, A.; Fernandez-Fuertes, B.; Recuero, S.; Mateo, Y.; Bonet, S.; Barranco, I.; Yeste, M. GSTM3, but not IZUMO1, is a cryotolerance marker of boar sperm. J. Anim. Sci. Biotechnol. 2019, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Gundogan, G.I.; Aktas, A. Immunolocalization of Fertilin β, IZUMO1, and P34H in Ram Spermatozoa. Biopreserv. Biobank. 2021, 19, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Zhi-da, L.; Wan-jin, X.; Lian-qing, W.; Li-xia, L. Prokaryotic Expression, Ascitic Polyclonal Antibody Preparation and Identification of Cashmere Goat Izumo1. Agric. Sci. China 2010, 9, 605–613. [Google Scholar] [CrossRef]
- Lorenzetti, D.; Poirier, C.; Zhao, M.; Overbeek, P.A.; Harrison, W.; Bishop, C.E. A transgenic insertion on mouse chromosome 17 inactivates a novel immunoglobulin superfamily gene potentially involved in sperm-egg fusion. Mamm. Genome 2014, 25, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Miyado, K.; Yamada, G.; Yamada, S.; Hasuwa, H.; Nakamura, Y.; Ryu, F.; Suzuki, K.; Kosai, K.; Inoues, K.; Ogura, A.; et al. Requirement of CD9 on the Egg Plasma Membrane for Fertilization. Science 2000, 287, 321–324. [Google Scholar] [CrossRef]
- Okabe, M. The Acrosome Reaction: A Historical Perspective. In Advances in Anatomy, Embryology, and Cell Biology; Springer: Berlin/Heidelberg, Germany, 2016; Volume 220, pp. 1–13. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Torres, M.J.; Hernández-Falcó, M.; López-Botella, A.; Huerta-Retamal, N.; Sáez-Espinosa, P. IZUMO1 Receptor Localization during Hyaluronic Acid Selection in Human Spermatozoa. Biomedicines 2023, 11, 2872. https://doi.org/10.3390/biomedicines11112872
Gómez-Torres MJ, Hernández-Falcó M, López-Botella A, Huerta-Retamal N, Sáez-Espinosa P. IZUMO1 Receptor Localization during Hyaluronic Acid Selection in Human Spermatozoa. Biomedicines. 2023; 11(11):2872. https://doi.org/10.3390/biomedicines11112872
Chicago/Turabian StyleGómez-Torres, María José, Miranda Hernández-Falcó, Andrea López-Botella, Natalia Huerta-Retamal, and Paula Sáez-Espinosa. 2023. "IZUMO1 Receptor Localization during Hyaluronic Acid Selection in Human Spermatozoa" Biomedicines 11, no. 11: 2872. https://doi.org/10.3390/biomedicines11112872
APA StyleGómez-Torres, M. J., Hernández-Falcó, M., López-Botella, A., Huerta-Retamal, N., & Sáez-Espinosa, P. (2023). IZUMO1 Receptor Localization during Hyaluronic Acid Selection in Human Spermatozoa. Biomedicines, 11(11), 2872. https://doi.org/10.3390/biomedicines11112872