
A Normalization Protocol Reduces Edge Effect in High-Throughput Analyses of Hydroxyurea Hypersensitivity in Fission Yeast



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drug
2.2. Fission Yeast Techniques
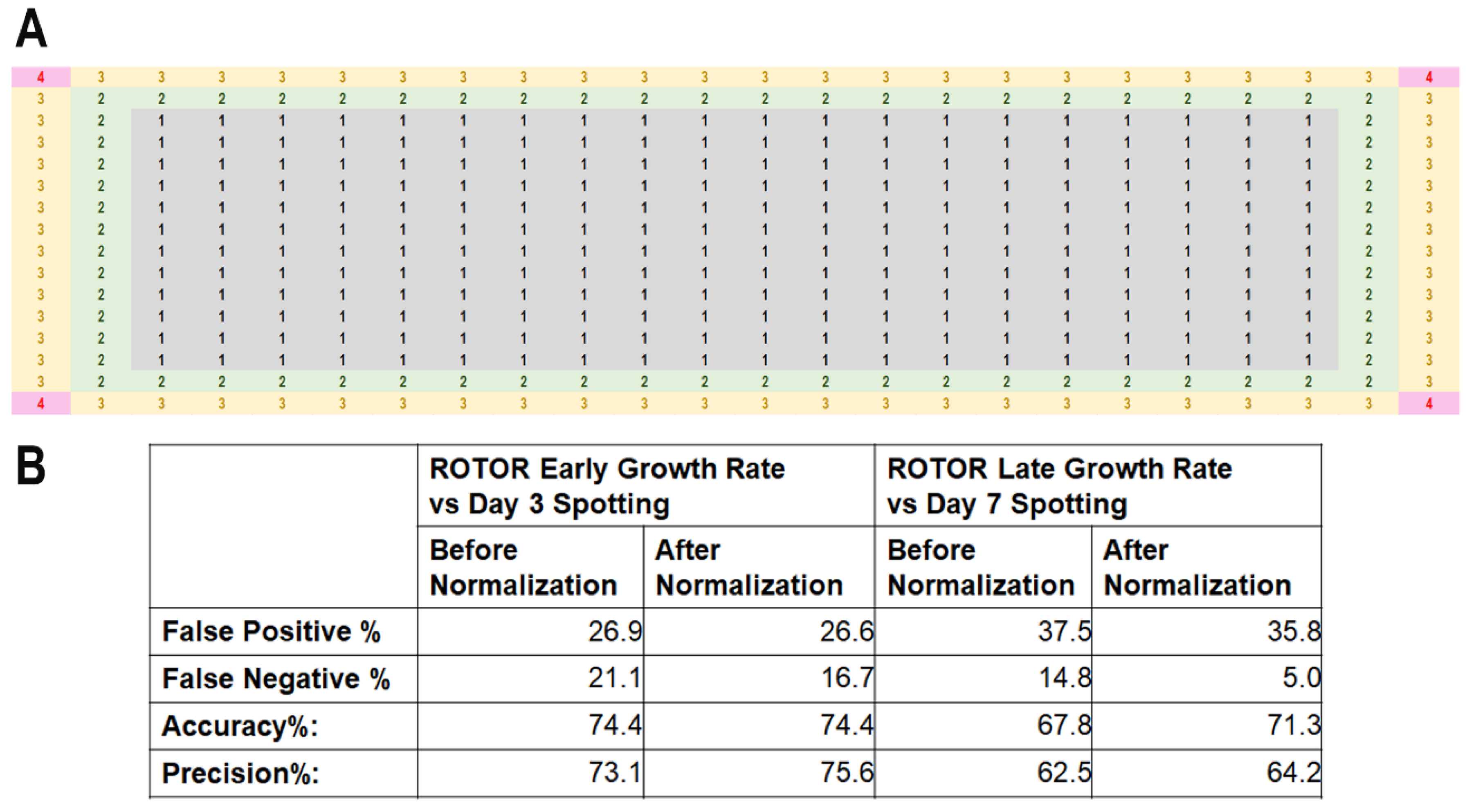
2.3. ROTOR Manipulation and Establishment of Normalization Table
2.4. Microbial Array Assay
2.5. Mathematical Analyses
3. Results
3.1. Subsection
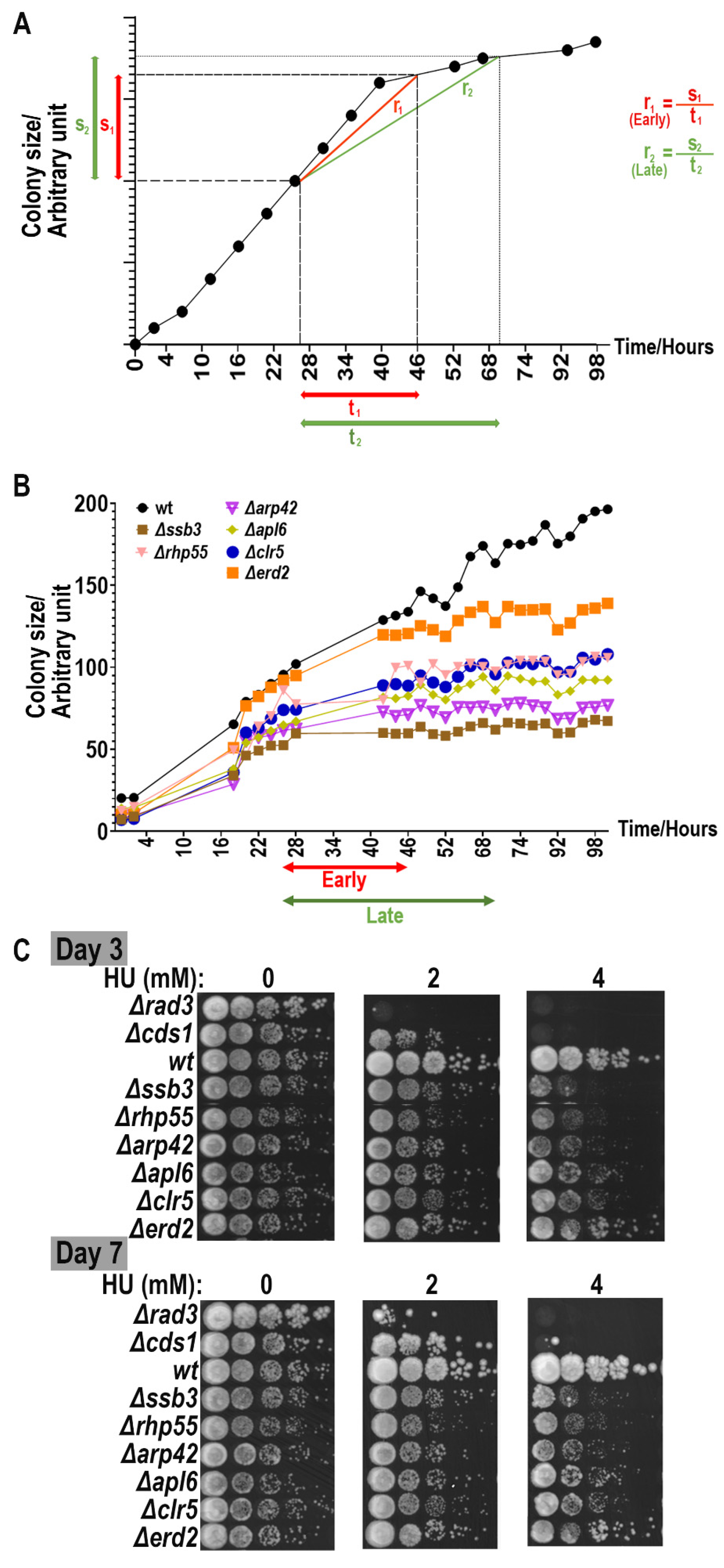
3.1.1. Time of Documenting Improves Description of Growth Outcome
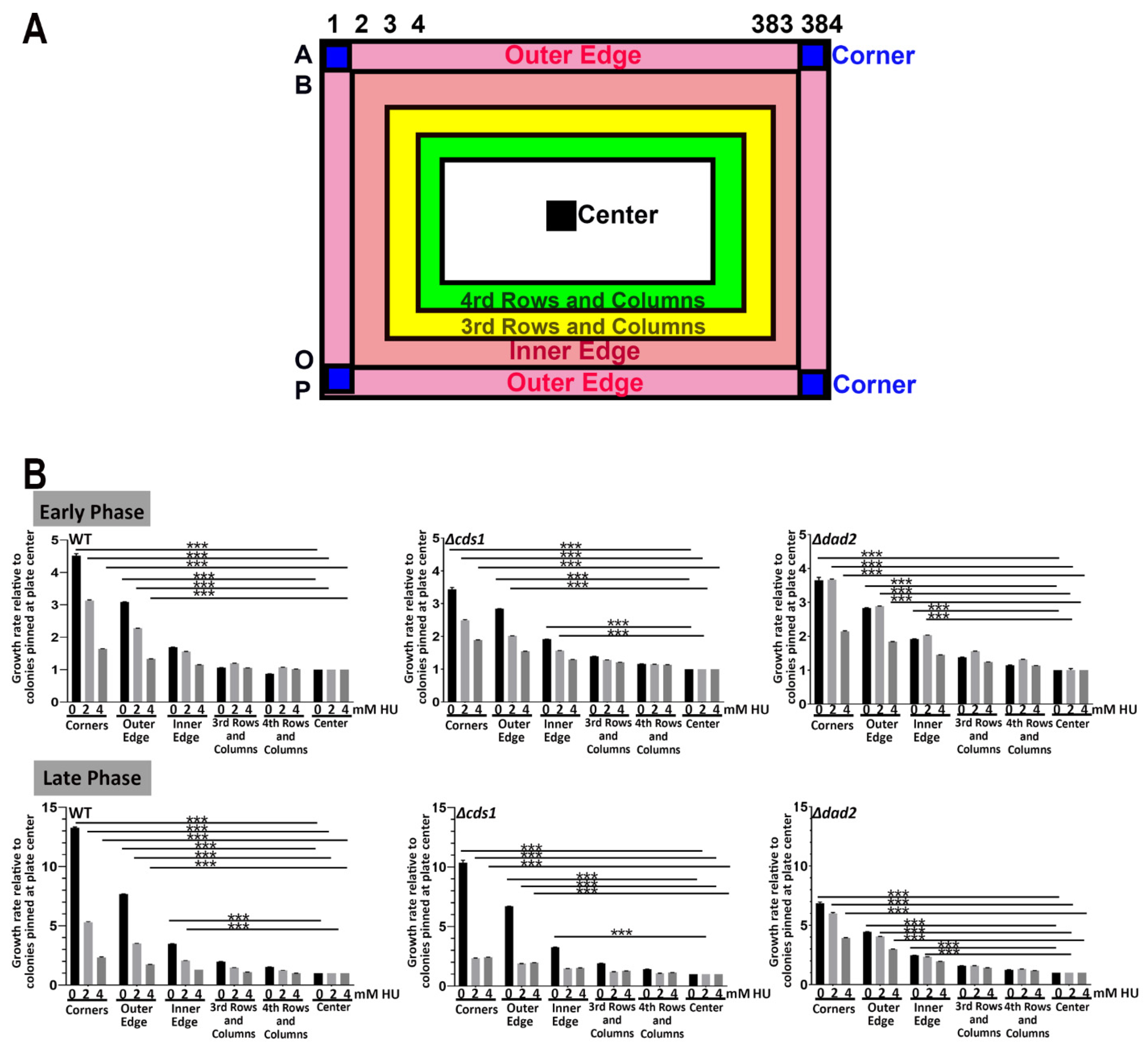
3.1.2. Growth Discrepancy of Colonies at Designated Positions on Agar Plates
3.1.3. Normalization for HU Screening with MER Strains
4. Discussion
4.1. Challenges of Colony Size Normalization via Image Processing Procedures
4.2. Imaging Quantification Versus Serial Dilution Assay
4.3. High-Density Array Analysis versus Plate Reader-Based Continuous Growth Measurement
4.4. Advantages of Colony Size Correction with a Normalization Table
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roguev, A.; Bandyopadhyay, S.; Zofall, M.; Zhang, K.; Fischer, T.; Collins, S.R.; Qu, H.; Shales, M.; Park, H.-O.; Hayles, J.; et al. Conservation and rewiring of functional modules revealed by an epistasis map in fission yeast. Science 2008, 322, 405–410. [Google Scholar] [CrossRef]
- Malecki, M.; Bähler, J. Identifying genes required for respiratory growth of fission yeast. Wellcome Open Res. 2016, 1, 12. [Google Scholar] [CrossRef] [PubMed]
- Rallis, C.; Bähler, J. Cell-based screens and phenomics with fission yeast. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Ojima, T.; Sugimoto, K.; Nakano, H.; Kawarasaki, Y. Quantitative Y2H screening: Cloning and signal peptide engineering of a fungal secretory LacA gene and its application to yeast two-hybrid system as a quantitative reporter. J. Biotechnol. 2010, 146, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Becker, F.; Murthi, K.; Smith, C.; Come, J.; Costa-Roldán, N.; Kaufmann, C.; Hanke, U.; Degenhart, C.; Baumann, S.; Wallner, W.; et al. A three-hybrid approach to scanning the proteome for targets of small molecule kinase inhibitors. Chem. Biol. 2004, 11, 211–223. [Google Scholar] [CrossRef]
- Parihar, A.; Pandita, V.; Khan, R. 3D printed human organoids: High throughput system for drug screening and testing in currrect COVID-19 pandemic. Biotechnol. Bioeng. 2022, 119, 2669–2688. [Google Scholar] [CrossRef]
- Butland, G.; Babu, M.; Díaz-Mejía, J.J.; Bohdana, F.; Phanse, S.; Gold, B.; Yang, W.; Li, J.; Gagarinova, A.G.; Pogoutse, O.; et al. eSGA: E. coli synthetic genetic array analysis. Nat. Methods 2008, 5, 789–795. [Google Scholar] [CrossRef]
- Louis, F.; Sowa, Y.; Kitano, S.; Matsusaki, M. High-throughput drug screening models of mature adipose tissues with replicable physiology of patients’ Body Mass Index (BMI). Bioact. Mater. 2022, 7, 227–241. [Google Scholar] [CrossRef]
- Giri, A.K.; Ianevski, A. High-throughput screening for drug discovery targeting the cancer cell-microenvironment interactions in haematological cancers. Expert Opin. Drug Discov. 2022, 17, 181–190. [Google Scholar] [CrossRef]
- Kaminska, J.; Soczewka, P.; Rzepnikowska, W.; Zoladek, T. Yeast as a model to find new drugs and drug targets for VPS13-dependent neurodegenerative diseases. Int. J. Mol. Sci. 2022, 23, 5106. [Google Scholar] [CrossRef]
- Yeh, C.-S.; Wang, Z.; Miao, F.; Ma, H.; Kao, C.-T.; Hsu, T.-S.; Yu, J.-H.; Hung, E.-T.; Lin, C.-C.; Kuan, C.-Y.; et al. A novel synthetic-genetic-array-based yeast one-hybrid system for high discovery rate and short processing time. Genome Res. 2019, 29, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Bohets, H.; Annaert, P.; Mannens, G.; Van Beijsterveldt, L.; Anciaux, K.; Verboven, P.; Meuldermans, W.; Lavrijsen, K. Strategies for adsorption screening in drug discovery and development. Curr. Top. Med. Chem. 2001, 1, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Niles, A.L.; Moravec, R.A.; Riss, T.L. In vitro viability and cytotoxicity testing and same-well multi-parametric combinations for high throughput screening. Curr. Chem. Genom. 2009, 3, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Niles, A.L.; Moravec, R.A.; Riss, T.L. Update on in vitro cytotoxicity assays for drug development. Exp. Opin. Drug Discov. 2008, 3, 655–669. [Google Scholar] [CrossRef]
- Schirle, M.; Jenkins, J.L. Identifying compound efficacy targets in phenotypic drug discovery. Drug Discov. Today 2016, 21, 82–89. [Google Scholar] [CrossRef]
- Van Vleet, T.R.; Liguori, M.J.; Lynch, J.J., III; Rao, M.; Warder, S. Screening strategies and methods for better off-target liability prediction and identification of small-molecule pharmaceuticals. SLAS Discov. 2019, 24, 1–24. [Google Scholar] [CrossRef]
- Bühler, M.; Spies, N.; Bartel, D.P.; Moazed, D. TRAMP-mediated RNA surveillance prevents spurious entry of RNAs into the Schizosaccharomyces pombe siRNA pathway. Nat. Struct. Mol. Biol. 2008, 15, 1015–1023. [Google Scholar] [CrossRef]
- Chen, E.S.; Zhang, K.; Nicolas, E.; Cam, H.P.; Zofall, M.; Grewal, S.I.S. Cell cycle control of centromeric repeat transcription and heterochromatin assembly. Nature 2008, 451, 734–737. [Google Scholar] [CrossRef]
- Chiron, S.; Gaisne, M.; Guillou, E.; Belenguer, P.; Clark-Walker, G.D.; Bonnefoy, N. Studying mitochondria in an attractive model: Schizosaccharomyces pombe. Methods Mol. Biol. 2007, 372, 91–105. [Google Scholar] [CrossRef]
- Lin, S.-J.; Austriaco, N. Aging and cell death in the other yeasts, Schizosaccharomyces pombe and Candida albicans. FEMS Yeast Res. 2014, 14, 119–135. [Google Scholar] [CrossRef]
- Hoffman, C.S.; Wood, V.; Fantes, P.A. An ancient yeast for young geneticists: A primer on the Schizosaccharomyces pombe model system. Genetics 2015, 201, 403–423. [Google Scholar] [CrossRef]
- Takahashi, K.; Chen, E.S.; Yanagida, M. Requirement of Mis6 centromere connector for localizing a CENP-A-like protein in fission yeast. Science 2000, 288, 2215–2219. [Google Scholar] [CrossRef] [PubMed]
- Stefanini, I.; De Filippo, C.; Cavalieri, D. Yeast as a model in high-throughput screening of small-molecule libraries. Drivers Synth. 2013, 455–482. [Google Scholar] [CrossRef]
- Kachroo, A.H.; Laurent, J.M.; Yellman, C.M.; Meyer, A.G.; Wilke, C.O.; Marcotte, E.M. Evolution. Systematic humanization of yeast genes reveals conserved functions and genetic modularity. Science 2015, 348, 921–925. [Google Scholar] [CrossRef]
- Sinclair, D.A. Studying the replicative life span of yeast cells. Methods Mol. Biol. 2013, 1048, 49–63. [Google Scholar] [CrossRef]
- Winzeler, E.A.; Shoemaker, D.D.; Astromoff, A.; Liang, H.; Anderson, K.; Andre, B.; Bangham, R.; Benito, R.; Boeke, J.D.; Bussey, H.; et al. Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science 1999, 285, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Roguev, A.; Wiren, M.; Weissman, J.S.; Krogan, N.J. High throughput genetic interaction mapping in the fission yeast Schizosaccharomyces pombe. Nat. Methods 2007, 4, 861–866. [Google Scholar] [CrossRef]
- Roguev, A.; Ryan, C.J.; Hartsuiker, E.; Krogan, N.J. High-throughput quantitative genetic interaction mapping in the fission yeast Schizosaccharomyces pombe. Cold Spring Harb. Protoc. 2018, 2018. [Google Scholar] [CrossRef]
- Baek, S.T.; Kim, D.-U.; Han, S.; Woo, I.S.; Nam, M.; Kim, L.; Heo, K.-S.; Lee, H.; Hwang, H.-R.; Choi, S.-J.; et al. Genome-wide drug-induced haploinsufficient screening of fission yeast for identification of hydrazinocurcumin targets. J. Microbiol. Biotechnol. 2008, 18, 263–269. [Google Scholar] [PubMed]
- Hagihara, K.; Kinoshita, K.; Ishida, K.; Hojo, S.; Kameoka, Y.; Satoh, R.; Takasaki, T.; Sugiura, R. A genome-wide screen for FTY720-sensitive mutants reveals genes required for ROS homeostasis. Microb. Cell 2017, 4, 390–401. [Google Scholar] [CrossRef]
- Satoh, R.; Hagihara, K.; Matsuura, K.; Manse, Y.; Kita, A.; Kunoh, T.; Masuko, T.; Moriyama, M.; Moriyama, H.; Tanabe, G.; et al. Identification of ACA-28, a 1′-acetoxychavicol acetate analogue compound, as a novel modulator of ERK MAPK signalling, which preferentially kills human melanoma cells. Genes Cells 2017, 22, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Mori, A.; Yanagida, M. Identification of genes affecting the toxicity of anti-cancer drug bortezomib by genome-wide screening in S. pombe. PLoS ONE 2011, 6, e22021. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.A.; Chen, E.S. Synthetic lethal approaches for assessing combinatorial efficacy of chemotherapeutic drugs. Pharmacol. Ther. 2016, 162, 69–85. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Chua, J.K.K.; Seah, K.S.; Koo, S.H.; Yee, J.Y.; Yang, E.G.; Lim, K.K.; Pang, S.Y.W.; Yuen, A.; Zhang, L.; et al. Predicting chemotherapeutic drug combinations through gene network profiling. Sci. Rep. 2016, 6, 18658. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Lim, Y.J.; Fan, M.H.M.; Jackson, R.A.; Lim, K.K.; Ang, W.H.; Ban, K.H.K.; Chen, E.S. Calcium modulation of doxorubicin cytotoxicity in yeast and human cells. Genes Cells 2016, 21, 226–240. [Google Scholar] [CrossRef]
- Seah, K.S.; Loh, J.Y.; Nguyen, T.T.T.; Tan, H.L.; Hutchinson, P.E.; Lim, K.K.; Dymock, B.W.; Long, Y.C.; Lee, E.J.D.; Shen, H.-M.; et al. SAHA and cisplatin sensitize gastric cancer cells to doxorubicin by induction of DNA damage, apoptosis and perturbation of AMPK-mTOR signalling. Exp. Cell Res. 2018, 370, 283–291. [Google Scholar] [CrossRef]
- Lee, M.; Han, S.; Chang, H.; Kwak, Y.-S.; Weller, D.M.; Kim, D. FitSearch: A robust way to interpret a yeast fitness profile in terms of drug’s mode-of-action. BMC Genom. 2013, 14 (Suppl. S1), S6. [Google Scholar] [CrossRef]
- Stirling, D.R.; Swain-Bowden, M.J.; Lucas, A.M.; Carpenter, A.E.; Cimini, B.A.; Goodman, A. CellProfiler 4: Improvements in speed, utility and usability. BMC Bioinform. 2021, 22, 433. [Google Scholar] [CrossRef]
- Collins, S.R.; Schuldiner, M.; Krogan, N.J.; Weissman, J.S. A strategy for extracting and analysing large-scale quantitative interaction data. Genome Biol. 2006, 7, R63. [Google Scholar] [CrossRef]
- Lawless, C.; Wilkinson, D.J.; Young, A.; Addinall, S.G.; Lydall, D.A. Colonyzer: Automated quantification of micro-organism growth characteristics on solid agar. BMC Bioinform. 2010, 11, 287. [Google Scholar] [CrossRef]
- Herricks, T.; Dilworth, D.J.; Mast, F.D.; Li, S.; Smith, J.J.; Ratushny, A.V.; Aitchison, J.D. One-cell doubling evaluation by living arrays of yeast. ODELAY! G3 2017, 7, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Zackrisson, M.; Hallin, J.; Ottosson, L.-G.; Dahl, P.; Fernandez-Parada, E.; Ländström, E.; Fernandez-Ricaud, L.; Kaferle, P.; Skyman, A.; Stenberg, S.; et al. Scan-o-matic: High-Resolution microbial phenomics at a massive scale. G3 2016, 6, 3003–3014. [Google Scholar] [CrossRef] [PubMed]
- Bischof, L.; Převorovský, M.; Rallis, C.; Jeffares, D.C.; Arzhaeva, Y.; Bähler, J. Spotsizer: High-throughput quantitative analysis of microbial growth. Biotechniques 2016, 61, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Parikh, S.B.; Castilho Coelho, N.; Carvunis, A.-R. LI Detector: A framework for sensitive colony-based screens regardless of the distribution of fitness effects. G3 2021, 11, jkaa068. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang, I.H.; Friman, O.; Guertin, D.A.; Chang, J.H.; Lindquist, R.A.; Moffat, J.; et al. CellProfiler: Image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef] [PubMed]
- Barton, D.B.H.; Georghiou, D.; Dave, N.; Alghamdi, M.; Walsh, T.A.; Louis, E.J.; Foster, S.S. PHENOS: A high-throughput and flexible tool for microorganism growth phenotyping on solid media. BMC Microbiol. 2018, 18, 9. [Google Scholar] [CrossRef]
- Dittmar, J.C.; Reid, R.J.D.; Rothstein, R. ScreenMill: A freely available software suite for growth measurement, analysis and visualization of high-throughput screen data. BMC Bioinform. 2010, 11, 353. [Google Scholar] [CrossRef]
- Young, B.P.; Loewen, C.J.R. Balony: A software package for analysis of data generated by synthetic genetic array experiments. BMC Bioinform. 2013, 14, 354. [Google Scholar] [CrossRef]
- French, S.; Mangat, C.; Bharat, A.; Côté, J.-P.; Mori, H.; Brown, E.D. A robust platform for chemical genomics in bacterial systems. Mol. Biol. Cell 2016, 27, 1015–1025. [Google Scholar] [CrossRef]
- Lundholt, B.K.; Scudder, K.M.; Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. J. Biomed. Screen. 2003, 8, 566–570. [Google Scholar] [CrossRef]
- Bean, G.J.; Jaeger, P.A.; Bahr, S.; Ideker, T. Development of ultra-high-density screening tools for microbial “omics”. PLoS ONE 2014, 9, e85177. [Google Scholar] [CrossRef] [PubMed]
- Wagih, O.; Usaj, M.; Baryshnikova, A.; VanderSluis, B.; Kuzmin, E.; Costanzo, M.; Myers, C.L.; Andrews, B.J.; Boone, C.M.; Parts, L. SGAtools: One-stop analysis and visualization of array-based genetic interaction screens. Nucleic Acids Res. 2013, 41, W591–W596. [Google Scholar] [CrossRef] [PubMed]
- Auld, D.S.P.; Coassin, P.A.B.S.; Coussens, N.P.P.; Hensley, P.; Klumpp-Thomas, C.; Michael, S.; Sittampalam, G.S.P.; Trask, O.J.B.S.; Wagner, B.K.P.; Weidner, J.R.P.; et al. Microplate selection and recommended practices in high-throughput screening and quantitative biology. In Assay Guidance Manual (Internet); Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]
- Ross, D.; Tonner, P.D.; Vasilyeva, O.B. Method for reproducible automated bacterial cell culture and measurement. Synth. Biol. 2022, 7, ysac013. [Google Scholar] [CrossRef]
- Kevorkov, D.; Makarenkov, V. Statistical analysis of systematic errors in high-throughput screening. J. Biomed. Screen. 2005, 10, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Brideau, C.; Gunter, B.; Pikounis, B.; Liaw, A. Improved statistical methods for hit selection in high-throughput screening. J. Biomed. Screen. 2003, 8, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Forsburg, S.L.; Rhind, N. Basic methods for fission yeast. Yeast 2006, 23, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Tay, Z.; Eng, R.J.; Sajiki, K.; Lim, K.K.; Tang, M.Y.; Yanagida, M.; Chen, E.S. Cellular robustness conferred by genetic crosstalk underlies resistance against chemotherapeutic drug doxorubicin in fission yeast. PLoS ONE 2013, 8, e55041. [Google Scholar] [CrossRef]
- Powers, D.M.W. Evaluation: From precision, recall and F-measure to ROC, informedness, markedness and correlation. J. Mach. Learn. Technol. 2022, 2, 37–63. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Lim, J.S.L.; Tang, R.M.Y.; Zhang, L.; Chen, E.S. Fitness profiling links topoisomerase II regulation of centromeric integrity to doxorubicin resistance in fission yeast. Sci. Rep. 2015, 5, 8400. [Google Scholar] [CrossRef]
- Lim, K.K.; Koh, N.Z.H.; Zeng, Y.B.; Chuan, J.K.; Raechell, R.; Chen, E.S. Resistance to chemotherapeutic 5-Fluorouracil conferred by modulation of heterochromatic integrity through Ino80 function in fission yeast. Int. J. Mol. Sci. 2023, 24, 10687. [Google Scholar] [CrossRef]
- Zeng, W.; Guo, L.; Xu, S.; Chen, J.; Zhou, J. High-throughput screening technology in industrial biotechnology. Trends Biotechnol. 2020, 38, 888–906. [Google Scholar] [CrossRef] [PubMed]
- Aguiar-Cervera, J.; Severn, O. Development of a new high-throughput method for screening large yeast libraries for use in the beverage industry. Access Microbiol. 2019, 1. [Google Scholar] [CrossRef]
- Mukherjee, V.; Radecka, D.; Aerts, G.; Verstrepen, K.J.; Lievens, B.; Thevelein, J.M. Phenotypic landscape of non-conventional yeast species for different stress tolerance traits desirable in bioethanol fermentation. Biotechnol. Biofuels 2017, 10, 216. [Google Scholar] [CrossRef]
- Bean, G.J.; Ideker, T. Differential analysis of high-throughput quantitative genetic interaction data. Genome Biol. 2012, 13, R123. [Google Scholar] [CrossRef]
- Baryshnikova, A.; Costanzo, M.; Kim, Y.; Ding, H.; Koh, J.; Toufighi, K.; Youn, J.-Y.; Ou, J.; San Luis, B.-J.; Bandyopadhyay, S.; et al. Quantitative analysis of fitness and genetic interactions in yeast on a genomic scale. Nat. Methods 2010, 7, 1017–1024. [Google Scholar] [CrossRef]
- Ard, R.; Tong, P.; Allshire, R.C. Long non-coding RNA-mediated transcriptional interference of a permease gene confers drug tolerance in fission yeast. Nat. Commun. 2014, 5, 5576. [Google Scholar] [CrossRef]
- Burr, R.; Stewart, E.V.; Shao, W.; Zhao, S.; Hannibal-Bach, H.K.; Ejsing, C.S.; Espenshade, P.J. Mga2 transcriptional factor regulates an oxygen-responsive lipid homeostasis pathway in fission yeast. J. Biol. Chem. 2016, 291, 12171–12183. [Google Scholar] [CrossRef]
- Sangkaew, A.; Krungkrai, J.; Yompakdee, C. Development of a high throughput yeast-based screening assay for human carbonic anhydrase isozyme II inhibitors. AMB Express 2018, 8, 124. [Google Scholar] [CrossRef]
- Rodríguez-López, M.; Gonzalez, S.; Hillson, O.; Tunnacliffe, E.; Codlin, S.; Tallada, V.A.; Bähler, J.; Rallis, C. The GATA transcription factor Gaf1 represses tRNAs, inhibits growth, and extends chronological lifespan downstream of fission yeast TORC1. Cell Rep. 2020, 30, 3240–3249. [Google Scholar] [CrossRef]
- Kim, C.; Yang, J.; Jeong, S.-H.; Kim, H.; Park, G.; Shin, H.B.; Ro, M.; Kim, K.-Y.; Park, Y.; Kim, K.P.; et al. Yeast-based assays for characterization of the functional effects of single nucleotide polymorphisms in human DNA repair gene. PLoS ONE 2018, 13, e0193823. [Google Scholar] [CrossRef]
- Kensy, F.; Zang, E.; Faulhammer, C.; Tan, R.; Büchs, J. Validation of a high-throughput fermentation system based on online monitoring of biomass and fluorescence in continuously shaken microtiter plates. Microb. Cell Fact. 2009, 8, 31. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, U.T.-F.; Nguyen, T.T.T.; Raechell, R.; Yang, J.; Singer, H.; Chen, E.S. A Normalization Protocol Reduces Edge Effect in High-Throughput Analyses of Hydroxyurea Hypersensitivity in Fission Yeast. Biomedicines 2023, 11, 2829. https://doi.org/10.3390/biomedicines11102829
Lam UT-F, Nguyen TTT, Raechell R, Yang J, Singer H, Chen ES. A Normalization Protocol Reduces Edge Effect in High-Throughput Analyses of Hydroxyurea Hypersensitivity in Fission Yeast. Biomedicines. 2023; 11(10):2829. https://doi.org/10.3390/biomedicines11102829
Chicago/Turabian StyleLam, Ulysses Tsz-Fung, Thi Thuy Trang Nguyen, Raechell Raechell, Jay Yang, Harry Singer, and Ee Sin Chen. 2023. "A Normalization Protocol Reduces Edge Effect in High-Throughput Analyses of Hydroxyurea Hypersensitivity in Fission Yeast" Biomedicines 11, no. 10: 2829. https://doi.org/10.3390/biomedicines11102829
APA StyleLam, U. T.-F., Nguyen, T. T. T., Raechell, R., Yang, J., Singer, H., & Chen, E. S. (2023). A Normalization Protocol Reduces Edge Effect in High-Throughput Analyses of Hydroxyurea Hypersensitivity in Fission Yeast. Biomedicines, 11(10), 2829. https://doi.org/10.3390/biomedicines11102829